
Impact of IL10, MTP, SOD2, and APOE Gene Polymorphisms on the Severity of Liver Fibrosis Induced by HCV Genotype 4


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Patients
2.3. DNA Extraction and Analysis of IL-10, MTP, SOD2, and ApoE Polymorphism
2.4. Statistical Analysis
3. Results
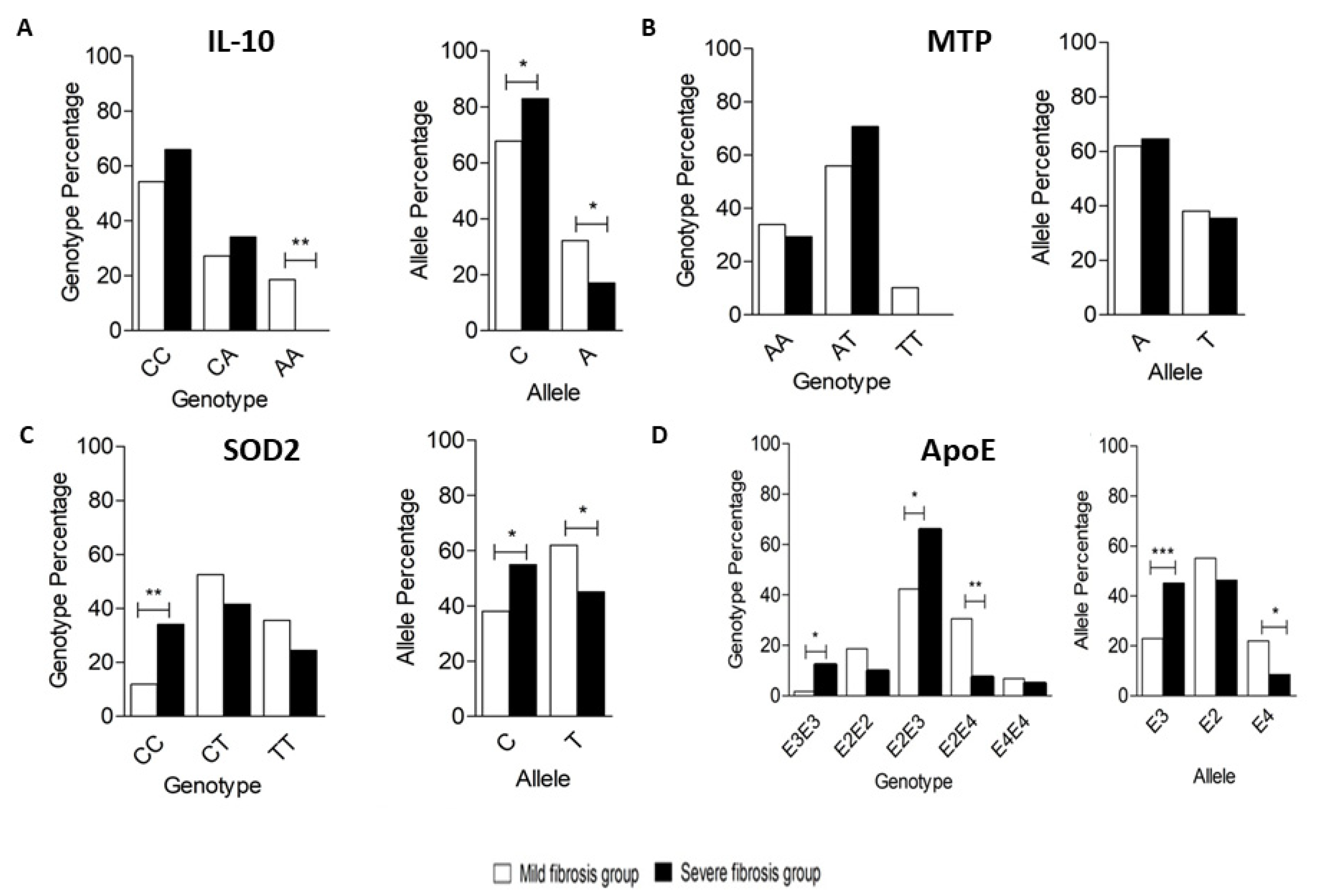
3.1. Genotyping of IL-10 Encoding Gene Polymorphism
3.2. Genotyping of MTP-Encoding Gene Polymorphism
3.3. Genotyping of SOD2-Encoding Gene Polymorphism
3.4. Genotyping of ApoE-Encoding Gene Polymorphism
3.5. Haplotype Analysis
3.5.1. Four Genes Haplotype Analysis
3.5.2. Three and Two Gene Haplotype Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohd Hanafiah, K.; Groeger, J.; Flaxman, A.D.; Wiersma, S.T. Global Epidemiology of Hepatitis C Virus Infection: New Estimates of Age-Specific Antibody to HCV Seroprevalence. Hepatology 2013, 57, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Elghitany, I. Hepatitis C Virus Infection in Egypt: Current Situation and Future Perspective. J. High Inst. Public Health 2019, 49, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Elgharably, A.; Gomaa, A.I.; Crossey, M.M.; Norsworthy, P.J.; Waked, I.; Taylor-Robinson, S.D. Hepatitis C in Egypt—Past, Present, and Future. Int. J. Genet. Med. 2016, 10, 1–6. [Google Scholar] [CrossRef]
- Gomaa, A.; Allam, N.; Elsharkway, A.; El Kassas, M.; Waked, I. Hepatitis C Infection in Egypt: Prevalence, Impact and Management Strategies. Hepatic Med. 2017, 9, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Bataller, R.; North, K.E.; Brenner, D.A. Genetic Polymorphisms and the Progression of Liver Fibrosis: A Critical Appraisal. Hepatology 2003, 37, 493–503. [Google Scholar] [CrossRef]
- Richardson, M.M.; Powell, E.E.; Barrie, H.D.; Clouston, A.D.; Purdie, D.M.; Jonsson, J.R. A Combination of Genetic Polymorphisms Increases the Risk of Progressive Disease in Chronic Hepatitis C. J. Med. Genet. 2005, 42, e45. [Google Scholar] [CrossRef]
- Namikawa, C.; Shu-Ping, Z.; Vyselaar, J.R.; Nozaki, Y.; Nemoto, Y.; Ono, M.; Akisawa, N.; Saibara, T.; Hiroi, M.; Enzan, H.; et al. Polymorphisms of Microsomal Triglyceride Transfer Protein Gene and Manganese Superoxide Dismutase Gene in Non-Alcoholic Steatohepatitis. J. Hepatol. 2004, 40, 781–786. [Google Scholar] [CrossRef]
- Hannivoort, R.A.; Hernandez-Gea, V.; Friedman, S.L. Genomics and Proteomics in Liver Fibrosis and Cirrhosis. Fibrogenesis Tissue Repair 2012, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Boyer, N.; Marcellin, P. Pathogenesis, Diagnosis and Management of Hepatitis C. J. Hepatol. 2000, 32, 98–112. [Google Scholar] [CrossRef]
- Nelson, D.R.; Lauwers, G.Y.; Lau, J.Y.; Davis, G.L. Interleukin 10 Treatment Reduces Fibrosis in Patients with Chronic Hepatitis C: A Pilot Trial of Interferon Nonresponders. Gastroenterology 2000, 118, 655–660. [Google Scholar] [CrossRef]
- Ollier, W.E.R. Cytokine Genes and Disease Susceptibility. Cytokine 2004, 28, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.S.; Tahir, S.; Salman, A.; Baig, T.A.; Shafi, T.; Zaidi, N.U.S.S.; Qadri, I. Analysis of Interleukin-10 Gene Polymorphisms and Hepatitis C Susceptibility in Pakistan. J. Infect. Dev. Ctries. 2011, 5, 473–479. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, L.B.; Véniant, M.; Borén, J.; Raabe, M.; Wong, J.S.; Tam, C.; Flynn, L.; Vanni-Reyes, T.; Gunn, M.D.; Goldberg, I.J.; et al. Genes for Apolipoprotein B and Microsomal Triglyceride Transfer Protein Are Expressed in the Heart: Evidence That the Heart Has the Capacity to Synthesize and Secrete Lipoproteins. Circulation 1998, 98, 13–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wetterau, J.R.; Zilversmit, D.B. Localization of Intracellular Triacylglycerol and Cholesteryl Ester Transfer Activity in Rat Tissues. Biochim. Biophys. Acta 1986, 875, 610–617. [Google Scholar] [CrossRef]
- Borén, J.; Véniant, M.M.; Young, S.G. Apo B100-Containing Lipoproteins Are Secreted by the Heart. J. Clin. Investig. 1998, 101, 1197–1202. [Google Scholar] [CrossRef]
- Mirandola, S.; Bowman, D.; Hussain, M.M.; Alberti, A. Hepatic Steatosis in Hepatitis C Is a Storage Disease Due to HCV Interaction with Microsomal Triglyceride Transfer Protein (MTP). Nutr. Metab. 2010, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide Dismutase Multigene Family: A Comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) Gene Structures, Evolution, and Expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Larrea, E.; Beloqui, O.; Muñoz-Navas, M.A.; Civeira, M.P.; Prieto, J. Superoxide Dismutase in Patients with Chronic Hepatitis C Virus Infection. Free Radic. Biol. Med. 1998, 24, 1235–1241. [Google Scholar] [CrossRef] [Green Version]
- Aizawa, Y.; Seki, N.; Nagano, T.; Abe, H. Chronic Hepatitis C Virus Infection and Lipoprotein Metabolism. World J. Gastroenterol. 2015, 21, 10299–10313. [Google Scholar] [CrossRef]
- Ishak, K.; Baptista, A.; Bianchi, L.; Callea, F.; De Groote, J.; Gudat, F.; Denk, H.; Desmet, V.; Korb, G.; MacSween, R.N. Histological Grading and Staging of Chronic Hepatitis. J. Hepatol. 1995, 22, 696–699. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Hemeda, A.A.; Aziz, R.K.; Abdel-Hakeem, M.S.; Ali-Tammam, M. Body Mass Index (BMI) and Alpha-Fetoprotein (AFP) Level Correlate with the Severity of HCV-Induced Fibrosis in a Cohort of Egyptian Patients with Chronic HCV. Future J. Pharm. Sci. 2020, 6, 71. [Google Scholar] [CrossRef]
- Grove, J.; Daly, A.K.; Bassendine, M.F.; Gilvarry, E.; Day, C.P. Interleukin 10 Promoter Region Polymorphisms and Susceptibility to Advanced Alcoholic Liver Disease. Gut 2000, 46, 540–545. [Google Scholar] [CrossRef] [Green Version]
- Karpe, F.; Lundahl, B.; Ehrenborg, E.; Eriksson, P.; Hamsten, A. A Common Functional Polymorphism in the Promoter Region of the Microsomal Triglyceride Transfer Protein Gene Influences Plasma LDL Levels. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 756–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabris, C.; Vandelli, C.; Toniutto, P.; Minisini, R.; Colletta, C.; Falleti, E.; Smirne, C.; Pirisi, M. Apolipoprotein E Genotypes Modulate Fibrosis Progression in Patients with Chronic Hepatitis C and Persistently Normal Transaminases. J. Gastroenterol. Hepatol. 2011, 26, 328–333. [Google Scholar] [CrossRef]
- Bataller, R.; Brenner, D.A. Hepatic Stellate Cells as a Target for the Treatment of Liver Fibrosis. Semin. Liver Dis. 2001, 21, 437–451. [Google Scholar] [CrossRef]
- Missiha, S.B.; Ostrowski, M.; Heathcote, E.J. Disease Progression in Chronic Hepatitis C: Modifiable and Nonmodifiable Factors. Gastroenterology 2008, 134, 1699–1714. [Google Scholar] [CrossRef] [PubMed]
- Coppola, N.; Pisaturo, M.; Sagnelli, C.; Onorato, L.; Sagnelli, E. Role of Genetic Polymorphisms in Hepatitis C Virus Chronic Infection. World J. Clin. Cases 2015, 3, 807–822. [Google Scholar] [CrossRef]
- Thio, C.L. Host Genetic Factors and Antiviral Immune Responses to Hepatitis C Virus. Clin. Liver Dis. 2008, 12, 713–726. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Joshi-Barve, S.; Barve, S.; McClain, C.J. Advances in Alcoholic Liver Disease. Curr. Gastroenterol. Rep. 2004, 6, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Fickenscher, H.; Hör, S.; Küpers, H.; Knappe, A.; Wittmann, S.; Sticht, H. The Interleukin-10 Family of Cytokines. Trends Immunol. 2002, 23, 89–96. [Google Scholar] [CrossRef]
- Winwood, P.J.; Arthur, M.J. Kupffer Cells: Their Activation and Role in Animal Models of Liver Injury and Human Liver Disease. Semin. Liver Dis. 1993, 13, 50–59. [Google Scholar] [CrossRef]
- Reuss, E.; Fimmers, R.; Kruger, A.; Becker, C.; Rittner, C.; Höhler, T. Differential Regulation of Interleukin-10 Production by Genetic and Environmental Factors—A Twin Study. Genes Immun. 2002, 3, 407–413. [Google Scholar] [CrossRef] [Green Version]
- Khatun, I.; Walsh, M.T.; Hussain, M.M. Loss of Both Phospholipid and Triglyceride Transfer Activities of Microsomal Triglyceride Transfer Protein in Abetalipoproteinemia. J. Lipid Res. 2013, 54, 1541–1549. [Google Scholar] [CrossRef] [Green Version]
- Rehberg, E.F.; Samson-Bouma, M.E.; Kienzle, B.; Blinderman, L.; Jamil, H.; Wetterau, J.R.; Aggerbeck, L.P.; Gordon, D.A. A Novel Abetalipoproteinemia Genotype. Identification of a Missense Mutation in the 97-KDa Subunit of the Microsomal Triglyceride Transfer Protein That Prevents Complex Formation with Protein Disulfide Isomerase. J. Biol. Chem. 1996, 271, 29945–29952. [Google Scholar] [CrossRef] [Green Version]
- Perlemuter, G.; Sabile, A.; Letteron, P.; Vona, G.; Topilco, A.; Chrétien, Y.; Koike, K.; Pessayre, D.; Chapman, J.; Barba, G.; et al. Hepatitis C Virus Core Protein Inhibits Microsomal Triglyceride Transfer Protein Activity and Very Low Density Lipoprotein Secretion: A Model of Viral-Related Steatosis. FASEB J. 2002, 16, 185–194. [Google Scholar] [CrossRef]
- Houldsworth, A.; Metzner, M.; Shaw, S.; Kaminski, E.; Demaine, A.G.; Cramp, M.E. Polymorphic Differences in SOD-2 May Influence HCV Viral Clearance. J. Med. Virol. 2014, 86, 941–947. [Google Scholar] [CrossRef]
- Ezzikouri, S.; Feydi, A.E.E.; Chafik, A.; Afifi, R.; Kihal, L.E.; Benazzouz, M.; Hassar, M.; Pineau, P.; Benjelloun, S. Genetic Polymorphism in the Manganese Superoxide Dismutase Gene Is Associated with an Increased Risk for Hepatocellular Carcinoma in HCV-Infected Moroccan Patients. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2008, 649, 1–6. [Google Scholar] [CrossRef]
- Shimoda-Matsubayashi, S.; Matsumine, H.; Kobayashi, T.; Nakagawa-Hattori, Y.; Shimizu, Y.; Mizuno, Y. Structural Dimorphism in the Mitochondrial Targeting Sequence in the Human Manganese Superoxide Dismutase Gene. A Predictive Evidence for Conformational Change to Influence Mitochondrial Transport and a Study of Allelic Association in Parkinson’s Disease. Biochem. Biophys. Res. Commun. 1996, 226, 561–565. [Google Scholar] [CrossRef]
- Agnello, V.; Abel, G.; Elfahal, M.; Knight, G.B.; Zhang, Q.X. Hepatitis C Virus and Other Flaviviridae Viruses Enter Cells via Low Density Lipoprotein Receptor. Proc. Natl. Acad. Sci. USA 1999, 96, 12766–12771. [Google Scholar] [CrossRef] [Green Version]
- André, P.; Komurian-Pradel, F.; Deforges, S.; Perret, M.; Berland, J.L.; Sodoyer, M.; Pol, S.; Bréchot, C.; Paranhos-Baccalà, G.; Lotteau, V. Characterization of Low- and Very-Low-Density Hepatitis C Virus RNA-Containing Particles. J. Virol. 2002, 76, 6919–6928. [Google Scholar] [CrossRef] [Green Version]
- Mahley, R.W.; Huang, Y. Alzheimer Disease: Multiple Causes, Multiple Effects of Apolipoprotein E4, and Multiple Therapeutic Approaches. Ann. Neurol. 2009, 65, 623–625. [Google Scholar] [CrossRef]
- Villeneuve, S.; Brisson, D.; Marchant, N.L.; Gaudet, D. The Potential Applications of Apolipoprotein E in Personalized Medicine. Front. Aging Neurosci. 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Monazahian, M.; Böhme, I.; Bonk, S.; Koch, A.; Scholz, C.; Grethe, S.; Thomssen, R. Low Density Lipoprotein Receptor as a Candidate Receptor for Hepatitis C Virus. J. Med. Virol. 1999, 57, 223–229. [Google Scholar] [CrossRef]
- Nascimento, J.; Matos, G.; Pereira, L.; Mourão, A.; Sampaio, A.; Oriá, R.; Toniutto, P. Impact of Apolipoprotein E Genetic Polymorphisms on Liver Disease: An Essential Review. Ann. Hepatol. 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, H.E.; Mahmoud, M.; Saad, N.E.; Saad-Hussein, A.; Ismail, S.; Thabet, E.H.; Farouk, H.; Kandil, D.; Heiba, A.; Hafez, W. Impact of Apo E Gene Polymorphism on HCV Therapy Related Outcome in a Cohort of HCV Egyptian Patients. J. Genet. Eng. Biotechnol. 2018, 16, 47–51. [Google Scholar] [CrossRef]
- Mueller, T.; Fischer, J.; Gessner, R.; Rosendahl, J.; Böhm, S.; van Bömmel, F.; Knop, V.; Sarrazin, C.; Witt, H.; Marques, A.M.; et al. Apolipoprotein E Allele Frequencies in Chronic and Self-Limited Hepatitis C Suggest a Protective Effect of APOE4 in the Course of Hepatitis C Virus Infection. Liver Int. 2016, 36, 1267–1274. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.M.; Bhattacharjee, P.S.; Neumann, D.M. Apolipoprotein E Alleles Can Contribute to the Pathogenesis of Numerous Clinical Conditions Including HSV-1 Corneal Disease. Exp. Eye Res. 2007, 84, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Price, D.A.; Bassendine, M.F.; Norris, S.M.; Golding, C.; Toms, G.L.; Schmid, M.L.; Morris, C.M.; Burt, A.D.; Donaldson, P.T. Apolipoprotein Epsilon3 Allele Is Associated with Persistent Hepatitis C Virus Infection. Gut 2006, 55, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, M.A.; Itzhaki, R.F.; Faragher, E.B.; James, M.W.; Ryder, S.D.; Irving, W.L. Apolipoprotein E-Ε4 Protects against Severe Liver Disease Caused by Hepatitis C Virus. Hepatology 2002, 36, 456–463. [Google Scholar] [CrossRef]
- Stachowska, E.; Maciejewska, D.; Ossowski, P.; Drozd, A.; Ryterska, K.; Banaszczak, M.; Milkiewicz, M.; Raszeja-Wyszomirska, J.; Slebioda, M.; Milkiewicz, P.; et al. Apolipoprotein E4 Allele Is Associated with Substantial Changes in the Plasma Lipids and Hyaluronic Acid Content in Patients with Nonalcoholic Fatty Liver Disease. J. Physiol. Pharmacol. 2013, 64, 711–717. [Google Scholar]
- Mueller, T.; Gessner, R.; Sarrazin, C.; Graf, C.; Halangk, J.; Witt, H.; Köttgen, E.; Wiedenmann, B.; Berg, T. Apolipoprotein E4 Allele Is Associated with Poor Treatment Response in Hepatitis C Virus (HCV) Genotype 1. Hepatology 2003, 38, 1592. [Google Scholar] [CrossRef]
- Lee, D.-M.; Lee, S.-O.; Mun, B.-S.; Ahn, H.-S.; Park, H.-Y.; Lee, H.-S.; Kim, D.-G. Relation of apolipoprotein E polymorphism to clinically diagnosed fatty liver disease. Taehan Kan Hakhoe Chi 2002, 8, 355–362. [Google Scholar]
- Zhen, J.; Huang, X.; Van Halm-Lutterodt, N.; Dong, S.; Ma, W.; Xiao, R.; Yuan, L. ApoE Rs429358 and Rs7412 Polymorphism and Gender Differences of Serum Lipid Profile and Cognition in Aging Chinese Population. Front. Aging Neurosci. 2017, 9. [Google Scholar] [CrossRef]
- Kadota, K.; Mori, M.; Yanagimachi, M.; Miyamae, T.; Hara, T.; Kanetaka, T.; Nozawa, T.; Kikuchi, M.; Hara, R.; Imagawa, T.; et al. Analysis of Gender Differences in Genetic Risk: Association of TNFAIP3 Polymorphism with Male Childhood-Onset Systemic Lupus Erythematosus in the Japanese Population. PLoS ONE 2013, 8, e72551. [Google Scholar] [CrossRef] [Green Version]
- Gayam, V.; Mandal, A.K.; Khalid, M.; Mukhtar, O.; Gill, A.; Garlapati, P.; Khalid, M.; Mansour, M. Sofosbuvir Based Regimens in the Treatment of Chronic Hepatitis C with Compensated Liver Cirrhosis in Community Care Setting. Int. J. Hepatol. 2018, 2018, e4136253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaldi, L.; Perrella, A.; Guarino, M.; De Luca, M.; Piai, G.; Coppola, N.; Pafundi, P.C.; Ciardiello, F.; Fasano, M.; Martinelli, E.; et al. Incidence and Risk Factors of Early HCC Occurrence in HCV Patients Treated with Direct Acting Antivirals: A Prospective Multicentre Study. J. Transl. Med. 2019, 17, 292. [Google Scholar] [CrossRef]
- Hsu, S.-J.; Yang, S.-S.; Kao, J.-H. Risk of Hepatocellular Carcinoma Development after Hepatitis C Virus Eradicated by Direct-Acting Antivirals: Fact or Fiction? J. Formos. Med. Assoc. 2020, 119, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Harrington, P.R.; Komatsu, T.E.; Deming, D.J.; Donaldson, E.F.; O’Rear, J.J.; Naeger, L.K. Impact of Hepatitis C Virus Polymorphisms on Direct-Acting Antiviral Treatment Efficacy: Regulatory Analyses and Perspectives. Hepatology 2018, 67, 2430–2448. [Google Scholar] [CrossRef] [Green Version]
- Zidan, H.E.; Talaat, R.M.; Ammar, A.A.A.; Sakr, M.A. Interleukin 28B Polymorphism as a Predictor of Response to Treatment of Egyptian HCV Patients Working in Nuclear Material Authority. Egypt. J. Hosp. Med. 2019, 77, 4742–4747. [Google Scholar] [CrossRef]
- Aly, A.M.; Adel, A.; El-Gendy, A.O.; Essam, T.M.; Aziz, R.K. Gut Microbiome Alterations in Patients with Stage 4 Hepatitis C. Gut Pathog. 2016, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Matute, P.; Íñiguez, M.; Villanueva-Millán, M.J.; Recio-Fernández, E.; Vázquez, A.M.; Sánchez, S.C.; Morano, L.E.; Oteo, J.A. Short-Term Effects of Direct-Acting Antiviral Agents on Inflammation and Gut Microbiota in Hepatitis C-Infected Patients. Eur. J. Intern. Med. 2019, 67, 47–58. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Oligonucleotide Sequence (5’-3’) | Product Size (bp) | Restriction Enzyme | Digested Products | Reference |
|---|---|---|---|---|---|
| IL10 (-627) | Forward: CCTAGGTCACAGTGACGTGG Reverse: GGTGAGCACTACCTGACTAGC | 412 | RsaI | 1 band of 412 bp for CC genotype 2 bands of 212 and 200 bp for AA genotype 3 bands of 412, 212, and 200 bp for AC genotype | [22] |
| MTP (-400) | Forward: GTCCATACAAGAAAAATTAAAATTTGGTTAG Reverse: GTCCATACAAGAAATATTAAAATTTGGTTAG | 838 | SspI | 1 band of 838 bp for AA genotype 2 bands of 494 and 344 bp for TT genotype 3 bands of 838, 494, and 344 bp for AT genotype. | [23] |
| SOD2 (1183) | Forward: CTGACCGGGCTGTGCTTTCTCG Reverse: CTCCCGCCGCTCAGCCTGGACC | 258 | BsaWI | 1 band of 258 bp for CC genotype 2 bands of 210 and 48 bp for AA genotype 3 bands of 258, 210, and 48 bp for CT genotype. | [6] |
| APOE | Outer forward: TGAAGGAGTTGAAGGCCTAC | 367 | HaeII & AflIII | 3 bands of 145, 50, and 23 bp for E3 allele 2 bands of 168 and 50 bp for E2 allele 2 bands of 195 and 23 bp for E4 allele | [24] |
| Outer reverse: CACGCGGCCCTGTTCCACCA | |||||
| Inner forward: | |||||
| TCCAAGGAGCTGCAGGCGGCGCA | 218 | ||||
| Inner reverse: | |||||
| GCCCCGGCCTGGTACACTGCCA |
| Gene Polymorphisms | Mild Group (N = 59) | Severe Group (N = 41) | OR | p-Value |
|---|---|---|---|---|
| % | % | |||
| IL10 Polymorphism | ||||
| Genotype | ||||
| CC | 54.2 | 65.9 | Ref. | |
| CA | 27.1 | 34.1 | 1.037 | 0.936 |
| AA | 18.6 | 0.0 | – | – |
| Allele | ||||
| C | 67.8 | 82.9 | Ref. | – |
| A | 32.2 | 17.1 | 0.433 | 0.016 * |
| MTP Polymorphism | ||||
| Genotype | ||||
| AA | 33.9 | 29.3 | Ref. | |
| AT | 55.9 | 70.7 | 1.465 | 0.390 |
| TT | 10.2 | 0.0 | – | – |
| Allele | ||||
| A | 61.9 | 64.6 | Ref. | – |
| T | 38.1 | 35.4 | 0.888 | 0.690 |
| SOD2 Polymorphism | ||||
| Genotype | ||||
| CC | 11.9 | 34.1 | Ref. | – |
| CT | 52.5 | 41.5 | 0.274 | 0.016 * |
| TT | 35.6 | 24.4 | 0.238 | 0.015 * |
| Allele | ||||
| C | 38.1 | 54.9 | Ref. | – |
| T | 61.9 | 45.1 | 0.507 | 0.019 * |
| APOE Polymorphism | ||||
| Genotype | ||||
| E3/E3 | 1.7 | 12.2 | Ref. | – |
| E2/E2 | 18.6 | 9.8 | 0.072 | 0.046 * |
| E2/E3 | 42.4 | 65.9 | 0.216 | 0.209 |
| E2/E4 | 30.5 | 7.3 | 0.033 | 0.004 ** |
| E4/E4 | 6.8 | 4.9 | 0.100 | 0.242 |
| Allele | ||||
| E3 | 22.9 | 45.1 | Ref. | – |
| E2 | 55.1 | 46.3 | 0.427 | 0.009 ** |
| E4 | 22.0 | 8.5 | 0.197 | 0.001 ** |
| Allele Combinations | Mild Group (N = 59) | Severe Group (N = 41) | p-Value | OR (95%CI) |
|---|---|---|---|---|
| % | % | |||
| IL10 + MTP + SOD2 | ||||
| CAC | 21.2 | 42.7 | Ref. | 1.000 |
| CAT | 22.9 | 17.1 | 0.018 * | 0.370 (0.162–0.845) |
| CTC | 6.8 | 6.1 | 0.199 | 0.446 (0.131–1.527) |
| CTT | 16.9 | 17.1 | 0.112 | 0.500 (0.213–1.175) |
| AAC | 7.6 | 0.0 | –NA | –NA |
| AAT | 10.2 | 4.9 | 0.024 * | 0.238 (0.069–0.825) |
| ATC | 2.5 | 6.1 | 0.822 | 1.191 (0.260–5.447) |
| ATT | 11.9 | 6.1 | 0.019 * | 0.255 (0.081–0.800) |
| IL10+ MTP + APOE | ||||
| CAE3 | 17.8 | 39.0 | Ref. | 1.000 |
| CAE2 | 20.3 | 17.1 | 0.028 * | 0.383 (0.162–0.904) |
| CAE4 | 5.9 | 3.7 | 0.089 | 0.281 (0.065–1.21) |
| CTE3 | 2.5 | 2.4 | 0.387 | 0.438 (0.067–2.844) |
| CTE2 | 15.3 | 18.3 | 0.178 | 0.547 (0.227–1.317) |
| CTE4 | 5.9 | 2.4 | 0.049 * | 0.188 (0.035–0.991) |
| AAE3 | 2.5 | 2.4 | 0.387 | 0.438 (0.067–2.844) |
| AAE2 | 11.9 | 1.2 | 0.004 * | 0.047 (0.006–0.384) |
| AAE4 | 3.4 | 1.2 | 0.117 | 0.164 (0.017–1.571) |
| ATE3 | 0.0 | 1.2 | –NA | –NA |
| ATE2 | 7.6 | 9.8 | 0.337 | 0.164 (0.194–1.753) |
| ATE4 | 6.8 | 1.2 | 0.023 * | 0.082 (0.010–0.705) |
| IL10+ SOD2+ APOE | ||||
| CCE3 | 11.9 | 30.5 | Ref. | 1.000 |
| CCE2 | 11.9 | 15.9 | 0.200 | 0.520 (0.192–1.412) |
| CCE4 | 4.2 | 2.4 | 0.097 | 0.224 (0.038–1.309) |
| CTE3 | 8.5 | 11.0 | 0.228 | 0.504 (0.166–1.534) |
| CTE2 | 23.7 | 19.5 | 0.013 * | 0.320 (0.130–0.785) |
| CTE4 | 7.6 | 3.7 | 0.024 * | 0.187 (0.043–0.805) |
| ACE3 | 2.5 | 1.2 | 0.163 | 0.187 (0.018–1.969) |
| ACE2 | 6.8 | 4.9 | 0.068 | 0.280 (0.071–1.099) |
| ACE4 | 0.8 | 0.0 | –NA | –NA |
| ATE3 | 0.0 | 2.4 | –NA | –NA |
| ATE2 | 12.7 | 6.1 | 0.006 * | 0.187 (0.056–0.623) |
| ATE4 | 9.3 | 2.4 | 0.006 * | 0.102 (0.020–0.526) |
| MTP+ SOD2+ APOE | ||||
| ACE3 | 11.9 | 30.5 | Ref. | 1.000 |
| ACE2 | 13.6 | 9.8 | 0.020 * | 0.280 (0.096–0.818) |
| ACE4 | 3.4 | 2.4 | 0.170 | 0.280 (0.045–1.727) |
| ATE3 | 8.5 | 11.0 | 0.228 | 0.504 (0.166–1.534) |
| ATE2 | 18.6 | 8.5 | 0.002 * | 0.178 (0.061–0.521) |
| ATE4 | 5.9 | 2.4 | 0.035 * | 0.160 (0.029–0.878) |
| TCE3 | 2.5 | 1.2 | 0.163 | 0.187 (0.018–1.969) |
| TCE2 | 5.1 | 11.0 | 0.780 | 0.840 (0.247–2.853) |
| TCE4 | 1.7 | 0.0 | –NA | –NA |
| TTE3 | 0.0 | 2.4 | –NA | –NA |
| TTE2 | 17.8 | 17.1 | 0.040 * | 0.373 (0.146–0.957) |
| TTE4 | 11.0 | 3.7 | 0.005 * | 0.129 (0.031–0.532) |
| Allele Combinations | Mild Group (N = 59) | Severe Group (N = 41) | p-Value | OR (95%CI) |
|---|---|---|---|---|
| % | % | |||
| IL10 + MTP Polymorphism | ||||
| CA | 44.1 | 59.8 | Ref. | 1.000 |
| CT | 23.7 | 23.2 | 0.359 | 0.720(0.357–1.452) |
| AA | 17.8 | 4.9 | 0.006 * | 0.202(0.065–0.631) |
| AT | 14.4 | 12.2 | 0.290 | 0.624(0.261–1.495) |
| IL10 + SOD2 Polymorphism | ||||
| CC | 28.0 | 48.8 | Ref. | 1.000 |
| CT | 39.8 | 34.1 | 0.034 * | 0.491(0.255–0.948) |
| AC | 10.2 | 6.1 | 0.067 | 0.344(0.110–1.075) |
| AT | 22.0 | 11.0 | 0.006 * | 0.286(0.118–0.693) |
| IL10 + APOE Polymorphism | ||||
| CE3 | 20.3 | 41.5 | Ref. | 1.000 |
| CE2 | 35.6 | 35.4 | 0.046 * | 0.487 (0.241–0.986) |
| CE4 | 11.9 | 6.1 | 0.019 * | 0.252 (0.080–0.794) |
| AE3 | 2.5 | 3.7 | 0.685 | 0.706 (0.131–3.801) |
| AE2 | 19.5 | 11.0 | 0.007 * | 0.276 (0.109–0.701) |
| AE4 | 10.2 | 2.4 | 0.008 * | 0.118 (0.024–0.574) |
| MTP + SOD2 Polymorphism | ||||
| AC | 28.8 | 42.7 | Ref. | 1.000 |
| AT | 33.1 | 22.0 | 0.032 * | 0.448(0.216–0.931) |
| TC | 9.3 | 12.2 | 0.803 | 0.883(0.332–2.348) |
| TT | 28.8 | 23.2 | 0.103 | 0.543(0.261–1.130) |
| MTP + APOE Polymorphism | ||||
| AE3 | 20.3 | 41.5 | Ref. | 1.000 |
| AE2 | 32.2 | 18.3 | 0.002 * | 0.279 (0.126–0.616) |
| AE4 | 9.3 | 4.9 | 0.034 * | 0.257 (0.073–0.903) |
| TE3 | 2.5 | 3.7 | 0.685 | 0.706 (0.131–3.801) |
| TE2 | 22.9 | 28.0 | 0.191 | 0.601 (0.280–1.290) |
| TE4 | 12.7 | 3.7 | 0.004 * | 0.141 (0.037–0.542) |
| SOD2 + APOE Polymorphism | ||||
| CE3 | 14.4 | 31.7 | Ref. | 1.000 |
| CE2 | 18.6 | 20.7 | 0.128 | 0.505 (0.210–1.218) |
| CE4 | 5.1 | 2.4 | 0.081 | 0.218 (0.040–1.209) |
| TE3 | 8.5 | 13.4 | 0.539 | 0.719 (0.251–2.060) |
| TE2 | 36.4 | 25.6 | 0.005 * | 0.319 (0.143–0.713) |
| TE4 | 16.9 | 6.1 | 0.002 * | 0.164 (0.052–0.519) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hemeda, A.A.; Ahmad Mohamed, A.; Aziz, R.K.; Abdel-Hakeem, M.S.; Ali-Tammam, M. Impact of IL10, MTP, SOD2, and APOE Gene Polymorphisms on the Severity of Liver Fibrosis Induced by HCV Genotype 4. Viruses 2021, 13, 714. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040714
Hemeda AA, Ahmad Mohamed A, Aziz RK, Abdel-Hakeem MS, Ali-Tammam M. Impact of IL10, MTP, SOD2, and APOE Gene Polymorphisms on the Severity of Liver Fibrosis Induced by HCV Genotype 4. Viruses. 2021; 13(4):714. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040714
Chicago/Turabian StyleHemeda, Amr Ali, Amal Ahmad Mohamed, Ramy Karam Aziz, Mohamed S. Abdel-Hakeem, and Marwa Ali-Tammam. 2021. "Impact of IL10, MTP, SOD2, and APOE Gene Polymorphisms on the Severity of Liver Fibrosis Induced by HCV Genotype 4" Viruses 13, no. 4: 714. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040714