
In Vivo Models to Study the Pathogenesis of Extra-Respiratory Complications of Influenza A Virus Infection


{kind=link}
{kind=link}
Abstract
:1. Introduction
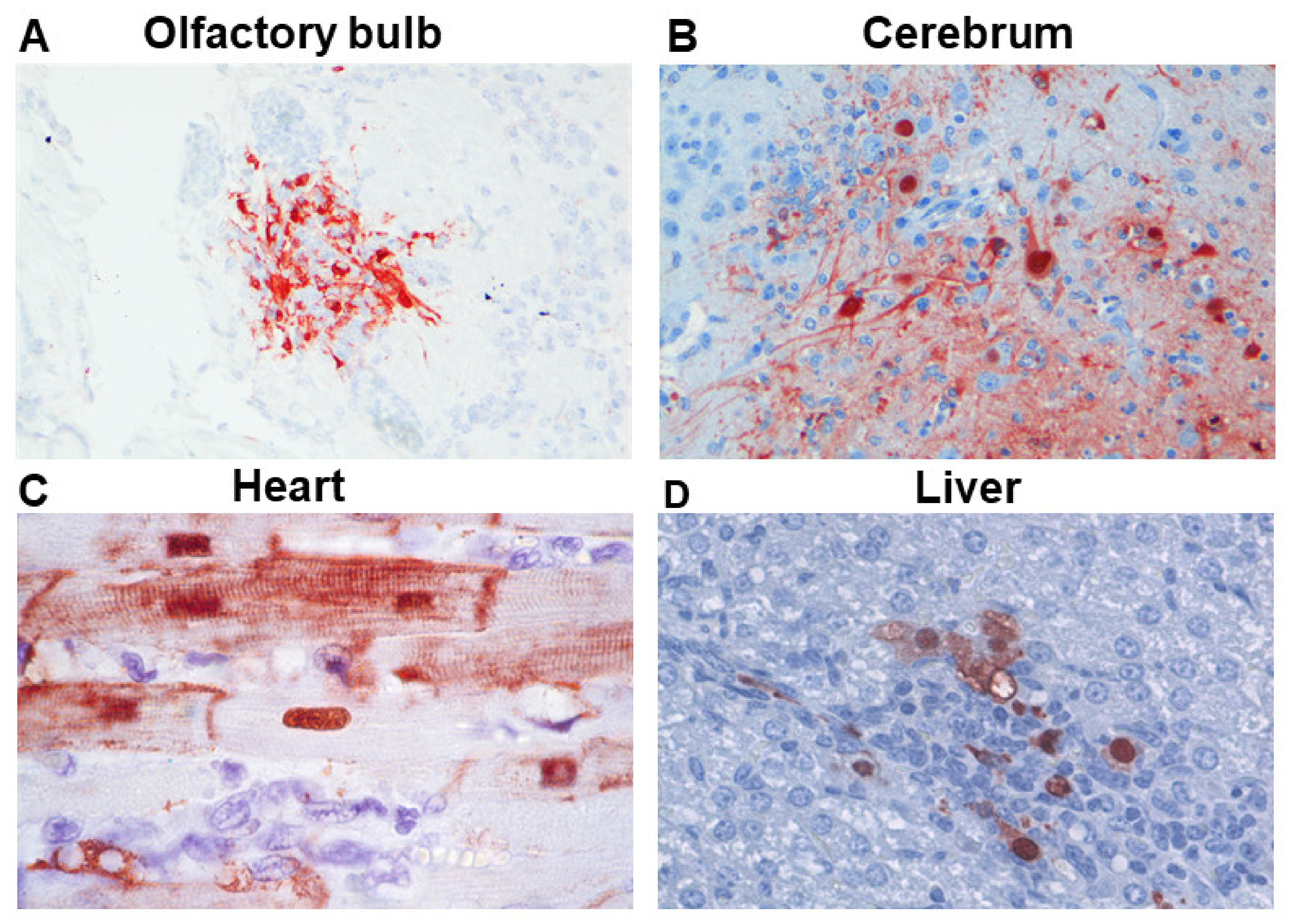
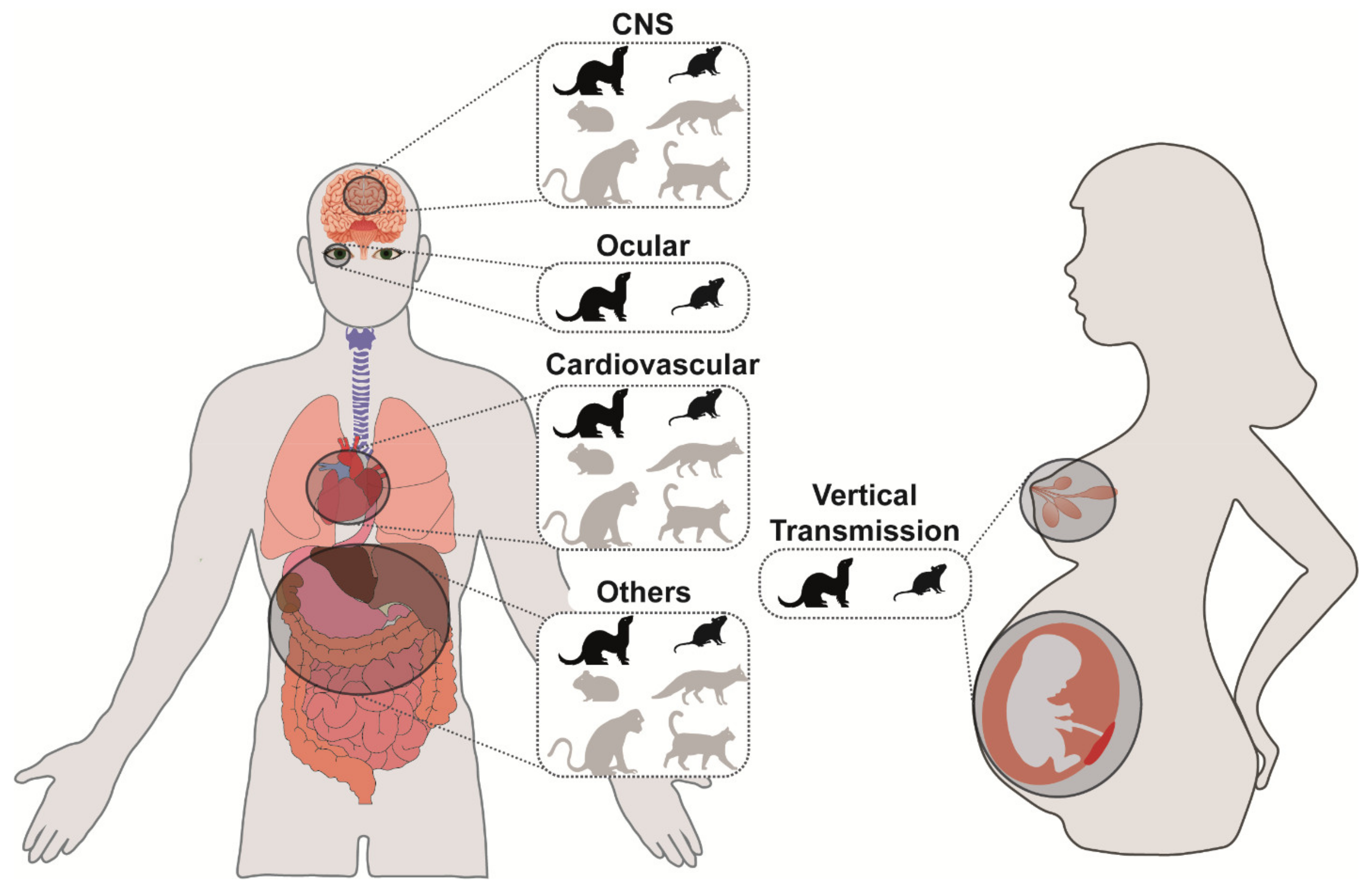
2. CNS Disease

3. Cardiovascular Disease
4. Other Diseases Involving the Eye, Placenta, Fetus, Lacteal Gland, Liver, Pancreas, Intestinal Tract, and Lymphoid Tissues
4.1. Ocular Disease
4.2. Disease Involving Pregnancy, Placenta, Fetus, and Lacteal Gland
4.3. Hematogenous Spread to Other Organs
| Extra-respiratory spread of IAVs in birds Influenza A virus (IAV) inoculation of birds does not represent an ideal in vivo model to study the extra-respiratory systemic pathogenesis of influenza in humans. There are substantial differences in the pathogenesis of influenza between birds and mammals, and also among different bird species, which compromises comparison. However, this does not imply that studying the pathogenesis of influenza in birds cannot lead to findings that can be extrapolated to the mammalian situation. Intravenous inoculation in six-week-old chickens is still used for the determination of the intravenous pathogenicity index (IVPI) that categorizes the pathogenicity of IAVs. Here, we will in short describe the pathogenesis of IAV infections in birds to highlight the differences with humans and other mammals. Within birds, IAV infection is associated with a different pattern of disease in water birds compared to poultry. Wild ducks and geese are considered the main natural hosts of IAVs and typically do not show signs of disease [87]. Like in humans, epithelial cells are the main target of infection in these birds; however, the virus mainly infects the intestinal epithelium, and less frequently the respiratory epithelium [88]. Poultry, and several wild bird species, can be infected by low pathogenic avian influenza viruses (LPAIVs) and by highly pathogenic avian influenza viruses (HPAIVs) [89]. Infection with LPAIVs in poultry typically produces limited clinical signs, and the virus preferentially targets epithelium of the respiratory tract. Infection with HPAIVs of the H5 and H7 subtypes is associated with severe systemic disease in poultry. HPAIVs target endothelial cells of multiple organs, such as lung, liver, heart, kidney, spleen, pancreas, bursa, and brain. The endotheliotropism of HPAIVs in poultry, which is not observed in humans, is associated with oedema formation and hemorrhages, coagulation disturbances, and induction of acute pro-inflammatory cytokines (cytokine storm), which may lead to acute mortality [90]. In wild birds, HPAIVs predominantly infect epithelial cells of the respiratory tract but not endothelial cells. Infection may lead to infection of parenchymal cells of other organ systems as well, associated with multi-organ necrosis and inflammation [91]. Amongst wild birds, the black swan is an exception because HPAIVs target systemic endothelial cells [92]. |
5. Discussion & Recommendations
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sellers, S.A.; Hagan, R.S.; Hayden, F.G.; Fischer, W.A. The hidden burden of influenza: A review of the extra-pulmonary complications of influenza infection. Influ. Other Respir. Viruses 2017, 11, 372–393. [Google Scholar] [CrossRef] [PubMed]
- Siegers, J.Y.; Van De Bildt, M.W.G.; Lin, Z.; Leijten, L.M.; Lavrijssen, R.A.M.; Bestebroer, T.; Spronken, M.I.J.; De Zeeuw, C.I.; Gao, Z.; Schrauwen, E.J.A.; et al. Viral Factors Important for Efficient Replication of Influenza A Viruses in Cells of the Central Nervous System. J. Virol. 2019, 93, e02273-18. [Google Scholar] [CrossRef] [Green Version]
- Martínez, A.; Soldevila, N.; Romero-Tamarit, A.; Torner, N.; Godoy, P.; Rius, C.; Jané, M.; Domínguez, À.; Surveillance of Hospitalized Cases of Severe Influenza in Catalonia Working Group. Risk factors associated with severe outcomes in adult hospitalized patients according to influenza type and subtype. PLoS ONE 2019, 14, e0210353. [Google Scholar] [CrossRef] [PubMed]
- Glaser, C.A.; Winter, K.; DuBray, K.; Harriman, K.; Uyeki, T.M.; Sejvar, J.; Gilliam, S.; Louie, J.K. A Population-Based Study of Neurologic Manifestations of Severe Influenza A(H1N1)pdm09 in California. Clin. Infect. Dis. 2012, 55, 514–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutleša, M.; Santini, M.; Krajinovic, V.; Raffanelli, D.; Baršić, B. Acute Motor Axonal Neuropathy Associated with Pandemic H1N1 Influenza A Infection. Neurocritical Care 2010, 13, 98–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-J.; Smith, D.S.; Rao, V.A.; Siegel, R.D.; Kosek, J.; Glaser, C.A.; Flint, A.C. Fatal H1N1-Related Acute Necrotizing Encephalopathy in an Adult. Case Rep. Crit. Care 2011, 2011, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Pula, J.H.; Issawi, A.; Desanto, J.R.; Kattah, J.C. Cortical Vision Loss as a Prominent Feature of H1N1 Encephalopathy. J. Neuro-Ophthalmol. 2012, 32, 48–50. [Google Scholar] [CrossRef]
- Gonzalez-Duarte, A.; Zamora, L.M.; Brito, C.C.; Garcia-Ramos, G. Hypothalamic abnormalities and Parkinsonism associated with H1N1 influenza infection. J. Neuroinflamm. 2010, 7, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amano, H.; Miyamoto, N.; Shimura, H.; Sato, D.K.; Fujihara, K.; Ueno, S.; Nakamura, R.; Ueno, Y.; Watanabe, M.; Hattori, N.; et al. Influenza-associated MOG antibody-positive longitudinally extensive transverse myelitis: A case report. BMC Neurol. 2014, 14, 224. [Google Scholar] [CrossRef] [Green Version]
- Landau, Y.E.; Grisaru-Soen, G.; Reif, S.; Fattal-Valevski, A. Pediatric Neurologic Complications Associated with Influenza A H1N. Pediatr. Neurol. 2011, 44, 47–51. [Google Scholar] [CrossRef]
- Kuiken, T.; Taubenberger, J.K. Pathology of human influenza revisited. Vaccine 2008, 26, D59–D66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Riel, D.; Verdijk, R.; Kuiken, T. The olfactory nerve: A shortcut for influenza and other viral diseases into the central nervous system. J. Pathol. 2015, 235, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Shinya, K.; Makino, A.; Hatta, M.; Watanabe, S.; Kim, J.H.; Hatta, Y.; Gao, P.; Ozawa, M.; Le, Q.M.; Kawaoka, Y. Subclinical Brain Injury Caused by H5N1 Influenza Virus Infection. J. Virol. 2011, 85, 5202–5207. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.; Boltz, D.; Sturm-Ramirez, K.; Shepherd, K.R.; Jiao, Y.; Webster, R.; Smeyne, R.J. Highly pathogenic H5N1 influenza virus can enter the central nervous system and induce neuroinflammation and neurodegeneration. Proc. Natl. Acad. Sci. USA 2009, 106, 14063–14068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrauwen, E.; Herfst, S.; Leijten, L.; Van Run, P.; Bestebroer, T.; Linster, M.; Bodewes, R.; Kreijtz, J.; Rimmelzwaan, G.; Osterhaus, A.; et al. The Multibasic Cleavage Site in H5N1 Virus Is Critical for Systemic Spread along the Olfactory and Hematogenous Routes in Ferrets. J. Virol. 2012, 86, 3975–3984. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Bingham, J.; Payne, J.; Rookes, J.; Lowther, S.; Haining, J.; Robinson, R.; Johnson, D.; Middleton, D. Multiple routes of invasion of wild-type Clade 1 highly pathogenic avian influenza H5N1 virus into the central nervous system (CNS) after intranasal exposure in ferrets. Acta Neuropathol. 2012, 124, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Plourde, J.R.; Pyles, J.A.; Layton, R.C.; Vaughan, S.E.; Tipper, J.L.; Harrod, K.S. Neurovirulence of H5N1 Infection in Ferrets Is Mediated by Multifocal Replication in Distinct Permissive Neuronal Cell Regions. PLoS ONE 2012, 7, e46605. [Google Scholar] [CrossRef] [Green Version]
- Kwon, D.; Shin, K.; Kim, S.; Ha, Y.; Choi, J.-H.; Yang, J.S.; Lee, J.-Y.; Chae, C.; Oh, H.-B.; Kang, C. Replication and pathogenesis of the pandemic (H1N1) 2009 influenza virus in mammalian models. J. Microbiol. 2010, 48, 657–662. [Google Scholar] [CrossRef]
- Iwasaki, T.; Itamura, S.; Nishimura, H.; Sato, Y.; Tashiro, M.; Hashikawa, T.; Kurata, T. Productive infection in the murine central nervous system with avian influenza virus A (H5N1) after intranasal inoculation. Acta Neuropathol. 2004, 108, 485–492. [Google Scholar] [CrossRef]
- Mori, I.; Goshima, F.; Imai, Y.; Kohsaka, S.; Sugiyama, T.; Yoshida, T.; Yokochi, T.; Nishiyama, Y.; Kimura, Y. Olfactory receptor neurons prevent dissemination of neurovirulent influenza A virus into the brain by undergoing virus-induced apoptosis. J. Gen. Virol. 2002, 83, 2109–2116. [Google Scholar] [CrossRef]
- Leyva-Grado, V.H.; Churchill, L.; Harding, J.; Krueger, J.M. The olfactory nerve has a role in the body temperature and brain cytokine responses to influenza virus. Brain Behav. Immun. 2010, 24, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majde, J.A.; Bohnet, S.G.; Ellis, G.A.; Churchill, L.; Leyva-Grado, V.; Wu, M.; Szentirmai, É.; Rehman, A.; Krueger, J.M. Detection of mouse-adapted human influenza virus in the olfactory bulbs of mice within hours after intranasal infection. J. NeuroVirol. 2007, 13, 399–409. [Google Scholar] [CrossRef]
- Park, C.H.; Ishinaka, M.; Takada, A.; Kida, H.; Kimura, T.; Ochiai, K.; Umemura, T. The invasion routes of neurovirulent A/Hong Kong/483/97 (H5N1) influenza virus into the central nervous system after respiratory infection in mice. Arch. Virol. 2002, 147, 1425–1436. [Google Scholar] [CrossRef] [PubMed]
- De Wit, E.; Siegers, J.Y.; Cronin, J.M.; Weatherman, S.; Brand, J.M.V.D.; Leijten, L.M.; Van Run, P.; Begeman, L.; Ham, H.-J.V.D.; Andeweg, A.C.; et al. 1918 H1N1 Influenza Virus Replicates and Induces Proinflammatory Cytokine Responses in Extrarespiratory Tissues of Ferrets. J. Infect. Dis. 2018, 217, 1237–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinya, K.; Shimada, A.; Ito, T.; Otsuki, K.; Morita, T.; Tanaka, H.; Takada, A.; Kida, H.; Umemura, T. Avian influenza virus intranasally inoculated infects the central nervous system of mice through the general visceral afferent nerve. Arch. Virol. 2000, 145, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Park, C.H.; Sunden, Y.; Kimura, T.; Ochiai, K.; Kida, H.; Umemura, T. The Vagus Nerve is One Route of Transneural Invasion for Intranasally Inoculated Influenza A Virus in Mice. Veter. Pathol. 2004, 41, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Park, C.-H.; Ninomiya, A.; Ozaki, H.; Takada, A.; Umemura, T.; Kida, H. Neurotropism of the 1997 Hong Kong H5N1 influenza virus in mice. Veter. Microbiol. 2003, 95, 1–13. [Google Scholar] [CrossRef]
- Van Riel, D.; Leijten, L.M.; Verdijk, R.M.; GeurtsvanKessel, C.; Van Der Vries, E.; Van Rossum, A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Evidence for Influenza Virus CNS Invasion Along the Olfactory Route in an Immunocompromised Infant. J. Infect. Dis. 2014, 210, 419–423. [Google Scholar] [CrossRef]
- Brand, J.M.A.V.D.; Stittelaar, K.J.; Van Amerongen, G.; Reperant, L.; De Waal, L.; Osterhaus, A.D.M.E.; Kuiken, T. Comparison of Temporal and Spatial Dynamics of Seasonal H3N2, Pandemic H1N1 and Highly Pathogenic Avian Influenza H5N1 Virus Infections in Ferrets. PLoS ONE 2012, 7, e42343. [Google Scholar] [CrossRef] [Green Version]
- Shinya, K.; Makino, A.; Tanaka, H.; Hatta, M.; Watanabe, T.; Le, M.Q.; Imai, H.; Kawaoka, Y. Systemic Dissemination of H5N1 Influenza A Viruses in Ferrets and Hamsters after Direct Intragastric Inoculation. J. Virol. 2011, 85, 4673–4678. [Google Scholar] [CrossRef] [Green Version]
- Zitzow, L.A.; Rowe, T.; Morken, T.; Shieh, W.-J.; Zaki, S.; Katz, J.M. Pathogenesis of Avian Influenza A (H5N1) Viruses in Ferrets. J. Virol. 2002, 76, 4420–4429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodewes, R.; Kreijtz, J.H.; van Amerongen, G.; Fouchier, R.A.; Osterhaus, A.D.; Rimmelzwaan, G.F.; Kuiken, T. Pathogenesis of Influenza A/H5N1 Virus Infection in Ferrets Differs between Intranasal and Intratracheal Routes of Inoculation. Am. J. Pathol. 2011, 179, 30–36. [Google Scholar] [CrossRef]
- Imai, M.; Watanabe, T.; Kiso, M.; Nakajima, N.; Yamayoshi, S.; Iwatsuki-Horimoto, K.; Hatta, M.; Yamada, S.; Ito, M.; Sakai-Tagawa, Y.; et al. A Highly Pathogenic Avian H7N9 Influenza Virus Isolated from A Human Is Lethal in Some Ferrets Infected via Respiratory Droplets. Cell Host Microbe 2017, 22, 615–626.e8. [Google Scholar] [CrossRef] [Green Version]
- Kalthoff, D.; Bogs, J.; Grund, C.; Tauscher, K.; Teifke, J.P.; Starick, E.; Harder, T.; Beer, M. Avian Influenza H7N9/13 and H7N7/13: A Comparative Virulence Study in Chickens, Pigeons, and Ferrets. J. Virol. 2014, 88, 9153–9165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lednicky, J.A.; Hamilton, S.B.; Tuttle, R.S.; Sosna, W.A.; Daniels, D.E.; Swayne, D.E. Ferrets develop fatal influenza after inhaling small particle aerosols of highly pathogenic avian influenza virus A/Vietnam/1203/2004 (H5N1). Virol. J. 2010, 7, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyva-Grado, V.H.; Churchill, L.; Wu, M.; Williams, T.J.; Taishi, P.; Majde, J.A.; Krueger, J.M. Influenza virus- and cytokine-immunoreactive cells in the murine olfactory and central autonomic nervous systems before and after illness onset. J. Neuroimmunol. 2009, 211, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronsson, F.; Robertson, B.; Ljunggren, H.-G.; Kristensson, K. Invasion and Persistence of the Neuroadapted Influenza Virus A/WSN/33 in the Mouse Olfactory System. Viral Immunol. 2003, 16, 415–423. [Google Scholar] [CrossRef]
- Hosseini, S.; Wilk, E.; Michaelsen-Preusse, K.; Gerhauser, I.; Baumgärtner, W.; Geffers, R.; Schughart, K.; Korte, M. Long-Term Neuroinflammation Induced by Influenza A Virus Infection and the Impact on Hippocampal Neuron Morphology and Function. J. Neurosci. 2018, 38, 3060–3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belser, J.A.; Lu, X.; Maines, T.R.; Smith, C.; Li, Y.; Donis, R.O.; Katz, J.M.; Tumpey, T.M. Pathogenesis of Avian Influenza (H7) Virus Infection in Mice and Ferrets: Enhanced Virulence of Eurasian H7N7 Viruses Isolated from Humans. J. Virol. 2007, 81, 11139–11147. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Iwatsuki-Horimoto, K.; Kiso, M.; Nakajima, N.; Takahashi, K.; Lopes, T.J.D.S.; Ito, M.; Fukuyama, S.; Hasegawa, H.; Kawaoka, Y. Experimental infection of Cynomolgus Macaques with highly pathogenic H5N1 influenza virus through the aerosol route. Sci. Rep. 2018, 8, 4801. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, Y.; Suguitan, A.; Orandle, M.; Paskel, M.; Boonnak, K.; Gardner, D.J.; Feldmann, F.; Feldmann, H.; Marino, M.; Jin, H.; et al. African Green Monkeys Recapitulate the Clinical Experience with Replication of Live Attenuated Pandemic Influenza Virus Vaccine Candidates. J. Virol. 2014, 88, 8139–8152. [Google Scholar] [CrossRef] [Green Version]
- Fujiyuki, T.; Yoneda, M.; Yasui, F.; Kuraishi, T.; Hattori, S.; Kwon, H.-J.; Munekata, K.; Kiso, Y.; Kida, H.; Kohara, M.; et al. Experimental Infection of Macaques with a Wild Water Bird-Derived Highly Pathogenic Avian Influenza Virus (H5N1). PLoS ONE 2013, 8, e83551. [Google Scholar] [CrossRef]
- Marriott, A.C.; Dennis, M.; Kane, J.A.; Gooch, K.E.; Hatch, G.; Sharpe, S.; Prevosto, C.; Leeming, G.; Zekeng, E.-G.; Staples, K.J.; et al. Influenza A Virus Challenge Models in Cynomolgus Macaques Using the Authentic Inhaled Aerosol and Intra-Nasal Routes of Infection. PLoS ONE 2016, 11, e0157887. [Google Scholar] [CrossRef] [Green Version]
- Kobasa, D.; Jones, S.M.; Shinya, K.; Kash, J.C.; Copps, J.; Ebihara, H.; Hatta, Y.; Kim, J.H.; Halfmann, P.; Hatta, M.; et al. Aberrant innate immune response in lethal infection of macaques with the 1918 influenza virus. Nat. Cell Biol. 2007, 445, 319–323. [Google Scholar] [CrossRef]
- Iwatsuki-Horimoto, K.; Nakajima, N.; Kiso, M.; Takahashi, K.; Ito, M.; Inoue, T.; Horiuchi, M.; Okahara, N.; Sasaki, E.; Hasegawa, H.; et al. The Marmoset as an Animal Model of Influenza: Infection With A(H1N1)pdm09 and Highly Pathogenic A(H5N1) Viruses via the Conventional or Tracheal Spray Route. Front. Microbiol. 2018, 9, 844. [Google Scholar] [CrossRef]
- Reperant, L.A.; Van Amerongen, G.; Van De Bildt, M.W.; Rimmelzwaan, G.F.; Dobson, A.P.; Osterhaus, A.D.; Kuiken, T. Highly Pathogenic Avian Influenza Virus (H5N1) Infection in Red Foxes Fed Infected Bird Carcasses. Emerg. Infect. Dis. 2008, 14, 1835–1841. [Google Scholar] [CrossRef]
- Rimmelzwaan, G.F.; Van Riel, D.; Baars, M.; Bestebroer, T.M.; Van Amerongen, G.; Fouchier, R.A.; Osterhaus, A.D.; Kuiken, T. Influenza A Virus (H5N1) Infection in Cats Causes Systemic Disease with Potential Novel Routes of Virus Spread within and between Hosts. Am. J. Pathol. 2006, 168, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Reperant, L.; Van De Bildt, M.; Van Amerongen, G.; Leijten, L.; Watson, S.; Palser, A.; Kellam, P.; Eissens, A.C.; Frijlink, H.W.; Osterhaus, A.D.M.E.; et al. Marked Endotheliotropism of Highly Pathogenic Avian Influenza Virus H5N1 following Intestinal Inoculation in Cats. J. Virol. 2011, 86, 1158–1165. [Google Scholar] [CrossRef] [Green Version]
- Madjid, M.; Miller, C.C.; Zarubaev, V.V.; Marinich, I.G.; Kiselev, O.I.; Lobzin, Y.V.; Filippov, A.E.; Casscells, S.W. Influenza epidemics and acute respiratory disease activity are associated with a surge in autopsy-confirmed coronary heart disease death: Results from 8 years of autopsies in 34 892 subjects. Eur. Heart J. 2007, 28, 1205–1210. [Google Scholar] [CrossRef] [Green Version]
- Corrales-Medina, V.F.; Madjid, M.; Musher, D.M. Role of acute infection in triggering acute coronary syndromes. Lancet Infect. Dis. 2010, 10, 83–92. [Google Scholar] [CrossRef]
- Warren-Gash, C.; Hayward, A.C.; Hemingway, H.; Denaxas, S.; Thomas, S.L.; Timmis, A.D.; Whitaker, H.J.; Smeeth, L. Influenza Infection and Risk of Acute Myocardial Infarction in England and Wales: A CALIBER Self-Controlled Case Series Study. J. Infect. Dis. 2012, 206, 1652–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naghavi, M.; Barlas, Z.; Siadaty, S.; Naguib, S.; Madjid, M.; Casscells, W. Association of Influenza Vaccination and Reduced Risk of Recurrent Myocardial Infarction. Circulation 2000, 102, 3039–3045. [Google Scholar] [CrossRef] [Green Version]
- Haidari, M.; Wyde, P.R.; Litovsky, S.; Vela, D.; Ali, M.; Casscells, S.W.; Madjid, M. Influenza virus directly infects, inflames, and resides in the arteries of atherosclerotic and normal mice. Atherosclerosis 2010, 208, 90–96. [Google Scholar] [CrossRef]
- Gao, P.; Watanabe, S.; Ito, T.; Goto, H.; Wells, K.; McGregor, M.; Cooley, A.J.; Kawaoka, Y. Biological Heterogeneity, Including Systemic Replication in Mice, of H5N1 Influenza A Virus Isolates from Humans in Hong Kong. J. Virol. 1999, 73, 3184–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogiwara, H.; Yasui, F.; Munekata, K.; Takagi-Kamiya, A.; Munakata, T.; Nomura, N.; Shibasaki, F.; Kuwahara, K.; Sakaguchi, N.; Sakoda, Y.; et al. Histopathological Evaluation of the Diversity of Cells Susceptible to H5N1 Virulent Avian Influenza Virus. Am. J. Pathol. 2014, 184, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Fislová, T.; Gocník, M.; Sládková, T.; Ďurmanová, V.; Rajčáni, J.; Varečková, E.; Mucha, V.; Kostolanský, F. Multiorgan distribution of human influenza A virus strains observed in a mouse model. Arch. Virol. 2009, 154, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.-Y.; Yamada, H.; Chida, J.; Wang, S.; Yano, M.; Yao, M.; Zhu, J.; Kido, H. Up-regulation of ectopic trypsins in the myocardium by influenza A virus infection triggers acute myocarditis. Cardiovasc. Res. 2010, 89, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, H.; Itamura, S.; Iwasaki, T.; Kurata, T.; Tashiro, M. Characterization of human influenza A (H5N1) virus infection in mice: Neuro-, pneumo- and adipotropic infection. J. Gen. Virol. 2000, 81, 2503–2510. [Google Scholar] [CrossRef] [Green Version]
- Kotaka, M.; Kitaura, Y.; Deguchi, H.; Kawamura, K. Experimental influenza A virus myocarditis in mice. Light and electron microscopic, virologic, and hemodynamic study. Am. J. Pathol. 1990, 136, 409–419. [Google Scholar] [PubMed]
- Kenney, A.D.; McMichael, T.M.; Imas, A.; Chesarino, N.M.; Zhang, L.; Dorn, L.E.; Wu, Q.; Alfaour, O.; Amari, F.; Chen, M.; et al. IFITM3 protects the heart during influenza virus infection. Proc. Natl. Acad. Sci. USA 2019, 116, 18607–18612. [Google Scholar] [CrossRef] [Green Version]
- Siegers, J.Y.; Novakovic, B.; Hulme, K.D.; Marshall, R.J.; Bloxham, C.J.; Thomas, W.G.; Reichelt, M.E.; Leijten, L.; Van Run, P.; Knox, K.; et al. A High-Fat Diet Increases Influenza A Virus-Associated Cardiovascular Damage. J. Infect. Dis. 2020, 222, 820–831. [Google Scholar] [CrossRef]
- Filgueiras-Rama, D.; Vasilijevic, J.; Jalife, J.; Noujaim, S.F.; Alfonso, J.M.; A Nicolas-Avila, J.; Gutierrez, C.; Zamarreño, N.; Hidalgo, A.; Bernabé, A.; et al. Human influenza A virus causes myocardial and cardiac-specific conduction system infections associated with early inflammation and premature death. Cardiovasc. Res. 2021, 117, 876–889. [Google Scholar] [CrossRef]
- Walters, K.-A.; D’Agnillo, F.; Sheng, Z.-M.; Kindrachuk, J.; Schwartzman, L.M.; Keustner, R.E.; Chertow, D.S.; Golding, B.T.; Taubenberger, J.K.; Kash, J.C. 1918 pandemic influenza virus and Streptococcus pneumoniae co-infection results in activation of coagulation and widespread pulmonary thrombosis in mice and humans. J. Pathol. 2016, 238, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koupenova, M.; Corkrey, H.A.; Vitseva, O.; Manni, G.; Pang, C.J.; Clancy, L.; Yao, C.; Rade, J.; Levy, D.; Wang, J.P.; et al. The role of platelets in mediating a response to human influenza infection. Nat. Commun. 2019, 10, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskin, C.R.; Bielefeldt-Ohmann, H.; Tumpey, T.M.; Sabourin, P.J.; Long, J.P.; García-Sastre, A.; Tolnay, A.-E.; Albrecht, R.; Pyles, J.A.; Olson, P.H.; et al. Early and sustained innate immune response defines pathology and death in nonhuman primates infected by highly pathogenic influenza virus. Proc. Natl. Acad. Sci. USA 2009, 106, 3455–3460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimmelzwaan, G.F.; Kuiken, T.; Van Amerongen, G.; Bestebroer, T.M.; Fouchier, R.A.M.; Osterhaus, A.D.M.E. Pathogenesis of Influenza A (H5N1) Virus Infection in a Primate Model. J. Virol. 2001, 75, 6687–6691. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Pak, C.; Han, Y.; Ahlenius, H.; Zhang, Z.; Chanda, S.; Marro, S.; Patzke, C.; Acuna, C.; Covy, J.; et al. Rapid Single-Step Induction of Functional Neurons from Human Pluripotent Stem Cells. Neuron 2013, 78, 785–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Bao, L.; Deng, W.; Qin, C. Highly pathogenic avian influenza H5N1 virus could partly be evacuated by pregnant BALB/c mouse during abortion or preterm delivery. Virol. J. 2011, 8, 342. [Google Scholar] [CrossRef] [Green Version]
- Jansen, A.J.G.; Spaan, T.; Low, H.Z.; Di Iorio, D.; Brand, J.V.D.; Tieke, M.; Barendrecht, A.; Rohn, K.; Van Amerongen, G.; Stittelaar, K.; et al. Influenza-induced thrombocytopenia is dependent on the subtype and sialoglycan receptor and increases with virus pathogenicity. Blood Adv. 2020, 4, 2967–2978. [Google Scholar] [CrossRef]
- Belser, J.A.; Rota, P.A.; Tumpey, T.M. Ocular Tropism of Respiratory Viruses. Microbiol. Mol. Biol. Rev. 2013, 77, 144–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belser, J.A.; Wadford, D.A.; Xu, J.; Katz, J.M.; Tumpey, T.M. Ocular Infection of Mice with Influenza A (H7) Viruses: A Site of Primary Replication and Spread to the Respiratory Tract. J. Virol. 2009, 83, 7075–7084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belser, J.A.; Zeng, H.; Katz, J.M.; Tumpey, T.M. Ocular Tropism of Influenza A Viruses: Identification of H7 Subtype-Specific Host Responses in Human Respiratory and Ocular Cells. J. Virol. 2011, 85, 10117–10125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belser, J.A.; Gustin, K.M.; Maines, T.R.; Pantin-Jackwood, M.J.; Katz, J.M.; Tumpey, T.M. Influenza Virus Respiratory Infection and Transmission Following Ocular Inoculation in Ferrets. PLoS Pathog. 2012, 8, e1002569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honce, R.; Wohlgemuth, N.; Meliopoulos, V.A.; Short, K.R.; Schultz-Cherry, S. Influenza in High-Risk Hosts—Lessons Learned from Animal Models. Cold Spring Harb. Perspect. Med. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Paquette, S.G.; Banner, D.; Huang, S.S.H.; Almansa, R.; Leon, A.; Xu, L.; Bartoszko, J.; Kelvin, D.J.; Kelvin, A.A. Influenza Transmission in the Mother-Infant Dyad Leads to Severe Disease, Mammary Gland Infection, and Pathogenesis by Regulating Host Responses. PLOS Pathog. 2015, 11, e1005173. [Google Scholar] [CrossRef] [PubMed]
- Brand, J.V.D.; Haagmans, B.; van Riel, D.; Osterhaus, A.; Kuiken, T. The Pathology and Pathogenesis of Experimental Severe Acute Respiratory Syndrome and Influenza in Animal Models. J. Comp. Pathol. 2014, 151, 83–112. [Google Scholar] [CrossRef] [Green Version]
- Cilloniz, C.; Pantin-Jackwood, M.J.; Ni, C.; Goodman, A.G.; Peng, X.; Proll, S.C.; Carter, V.S.; Rosenzweig, E.R.; Szretter, K.J.; Katz, J.M.; et al. Lethal Dissemination of H5N1 Influenza Virus Is Associated with Dysregulation of Inflammation and Lipoxin Signaling in a Mouse Model of Infection. J. Virol. 2010, 84, 7613–7624. [Google Scholar] [CrossRef] [Green Version]
- Capua, I.; Mercalli, A.; Romero-Tejeda, A.; Pizzuto, M.S.; Kasloff, S.; Sordi, V.; Marzinotto, I.; Lampasona, V.; Vicenzi, E.; De Battisti, C.; et al. Study of 2009 H1N1 Pandemic Influenza Virus as a Possible Causative Agent of Diabetes. J. Clin. Endocrinol. Metab. 2018, 103, 4343–4356. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-X.; Zhao, L.-Z.; Tang, S.-J.; Weng, T.-H.; Wu, W.-G.; Yao, S.-H.; Wu, H.-B.; Cheng, L.-F.; Wang, J.; Hu, F.-Y.; et al. Novel pathogenic characteristics of highly pathogenic avian influenza virus H7N9: Viraemia and extrapulmonary infection. Emerg. Microbes Infect. 2020, 9, 962–975. [Google Scholar] [CrossRef] [Green Version]
- Munster, V.J.; De Wit, E.; Van Riel, D.; Beyer, W.E.P.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E.; Kuiken, T.; Fouchier, R.A.M. The Molecular Basis of the Pathogenicity of the Dutch Highly Pathogenic Human Influenza A H7N7 Viruses. J. Infect. Dis. 2007, 196, 258–265. [Google Scholar] [CrossRef] [Green Version]
- Garigliany, M.-M.; Habyarimana, A.; Lambrecht, B.; Van De Paar, E.; Cornet, A.; Berg, T.V.D.; Desmecht, D. Influenza A Strain-Dependent Pathogenesis in Fatal H1N1 and H5N1 Subtype Infections of Mice. Emerg. Infect. Dis. 2010, 16, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Edenborough, K.M.; Lowther, S.; Laurie, K.; Yamada, M.; Long, F.; Bingham, J.; Payne, J.; Harper, J.; Haining, J.; Arkinstall, R.; et al. Predicting Disease Severity and Viral Spread of H5N1 Influenza Virus in Ferrets in the Context of Natural Exposure Routes. J. Virol. 2015, 90, 1888–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrauwen, E.J.; Herfst, S.; Chutinimitkul, S.; Bestebroer, T.M.; Rimmelzwaan, G.F.; Osterhaus, A.D.; Kuiken, T.; Fouchier, R.A. Possible Increased Pathogenicity of Pandemic (H1N1) 2009 Influenza Virus upon Reassortment. Emerg. Infect. Dis. 2011, 17, 200–208. [Google Scholar] [CrossRef]
- Lipatov, A.S.; Kwon, Y.K.; Pantin-Jackwood, M.J.; Swayne, D.E. Pathogenesis of H5N1 Influenza Virus Infections in Mice and Ferret Models Differs According to Respiratory Tract or Digestive System Exposure. J. Infect. Dis. 2009, 199, 717–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegers, J.Y.; Brand, J.M.V.D.; Leijten, L.M.; Van De Bildt, M.M.W.; Van Run, P.R.; Van Amerongen, G.; Stittelaar, K.J.; Koopmans, M.P.; Osterhaus, A.D.M.E.; Kuiken, T.; et al. Vaccination Is More Effective Than Prophylactic Oseltamivir in Preventing CNS Invasion by H5N1 Virus via the Olfactory Nerve. J. Infect. Dis. 2016, 214, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.-H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Pantin-Jackwook, M.; Swayne, D. Pathogenesis and pathobiology of avian influenza virus infection in birds. Rev. Sci. Tech. l’OIE 2009, 28, 113–136. [Google Scholar] [CrossRef]
- Short, K.R.; Kroeze, E.J.B.V.; Fouchier, R.A.M.; Kuiken, T. Pathogenesis of influenza-induced acute respiratory distress syndrome. Lancet Infect. Dis. 2014, 14, 57–69. [Google Scholar] [CrossRef]
- Lycett, S.J.; Pohlmann, A.; Staubach, C.; Caliendo, V.; Woolhouse, M.; Beer, M.; Kuiken, T. Genesis and spread of multiple reassortants during the 2016/2017 H5 avian influenza epidemic in Eurasia. Proc. Natl. Acad. Sci. USA 2020, 117, 20814–20825. [Google Scholar] [CrossRef]
- Burggraaf, S.; Karpala, A.J.; Bingham, J.; Lowther, S.; Selleck, P.; Kimpton, W.; Bean, A.G. H5N1 infection causes rapid mortality and high cytokine levels in chickens compared to ducks. Virus Res. 2014, 185, 23–31. [Google Scholar] [CrossRef]
- Caliendo, V.; Leijten, L.; Begeman, L.; Poen, M.J.; Fouchier, R.A.M.; Beerens, N.; Kuiken, T. Enterotropism of highly pathogenic avian influenza virus H5N8 from the 2016/2017 epidemic in some wild bird species. Veter. Res. 2020, 51, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.D.; Stallknecht, D.E.; Swayne, D.E. Experimental Infection of Swans and Geese with Highly Pathogenic Avian Influenza Virus (H5N1) of Asian Lineage. Emerg. Infect. Dis. 2008, 14, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Van Der Vries, E.; Stittelaar, K.J.; Van Amerongen, G.; Kroeze, E.J.B.V.; De Waal, L.; Fraaij, P.L.A.; Meesters, R.J.; Luider, T.M.; Van Der Nagel, B.; Koch, B.; et al. Prolonged Influenza Virus Shedding and Emergence of Antiviral Resistance in Immunocompromised Patients and Ferrets. PLoS Pathog. 2013, 9, e1003343. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veldhuis Kroeze, E.; Bauer, L.; Caliendo, V.; van Riel, D. In Vivo Models to Study the Pathogenesis of Extra-Respiratory Complications of Influenza A Virus Infection. Viruses 2021, 13, 848. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050848
Veldhuis Kroeze E, Bauer L, Caliendo V, van Riel D. In Vivo Models to Study the Pathogenesis of Extra-Respiratory Complications of Influenza A Virus Infection. Viruses. 2021; 13(5):848. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050848
Chicago/Turabian StyleVeldhuis Kroeze, Edwin, Lisa Bauer, Valentina Caliendo, and Debby van Riel. 2021. "In Vivo Models to Study the Pathogenesis of Extra-Respiratory Complications of Influenza A Virus Infection" Viruses 13, no. 5: 848. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050848