
A Semiquantitative Scoring System for Histopathological and Immunohistochemical Assessment of Lesions and Tissue Tropism in Avian Influenza

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. AIV Infection Experiments
2.2. Histopathological and Immunohistochemical Examination
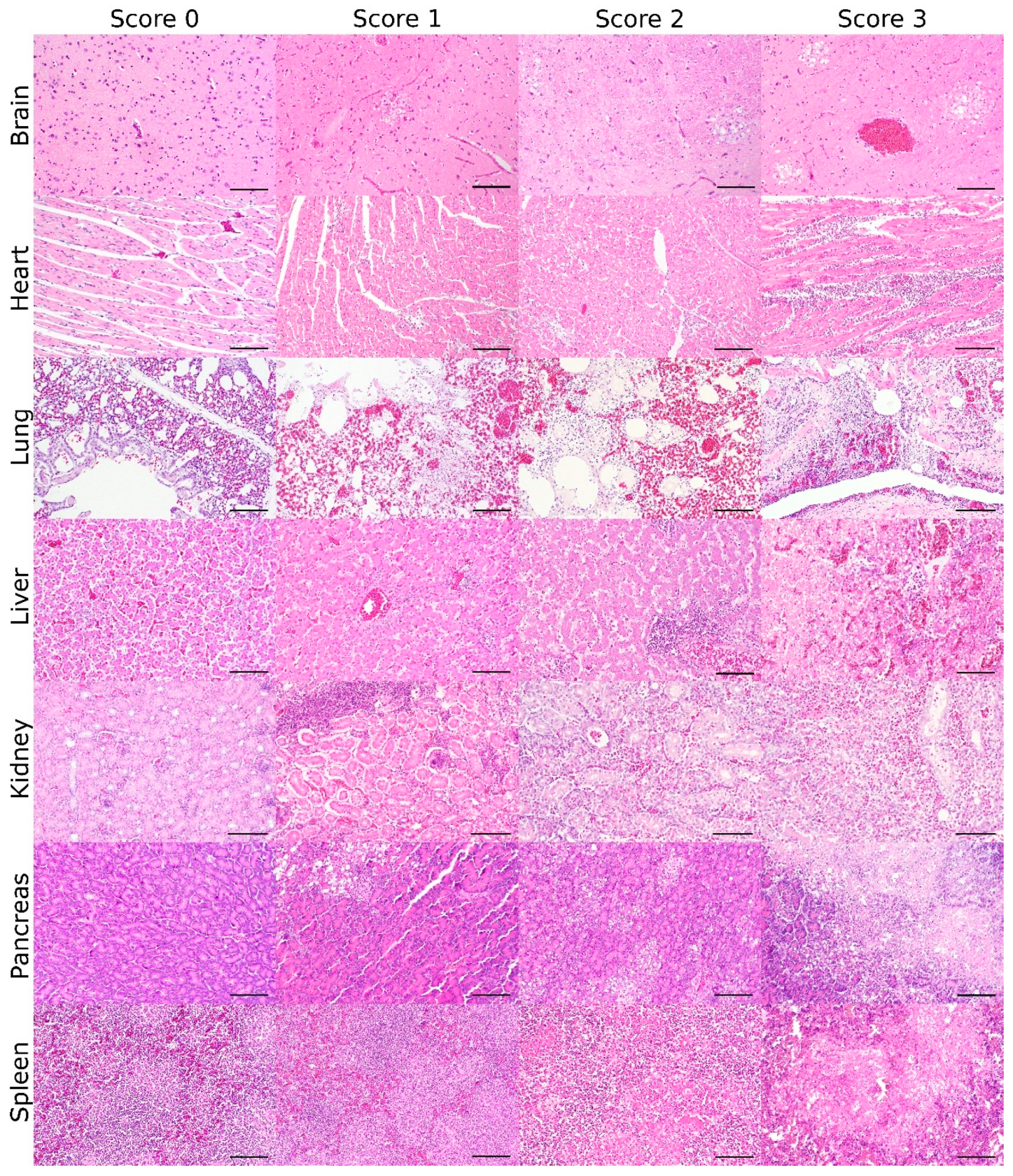
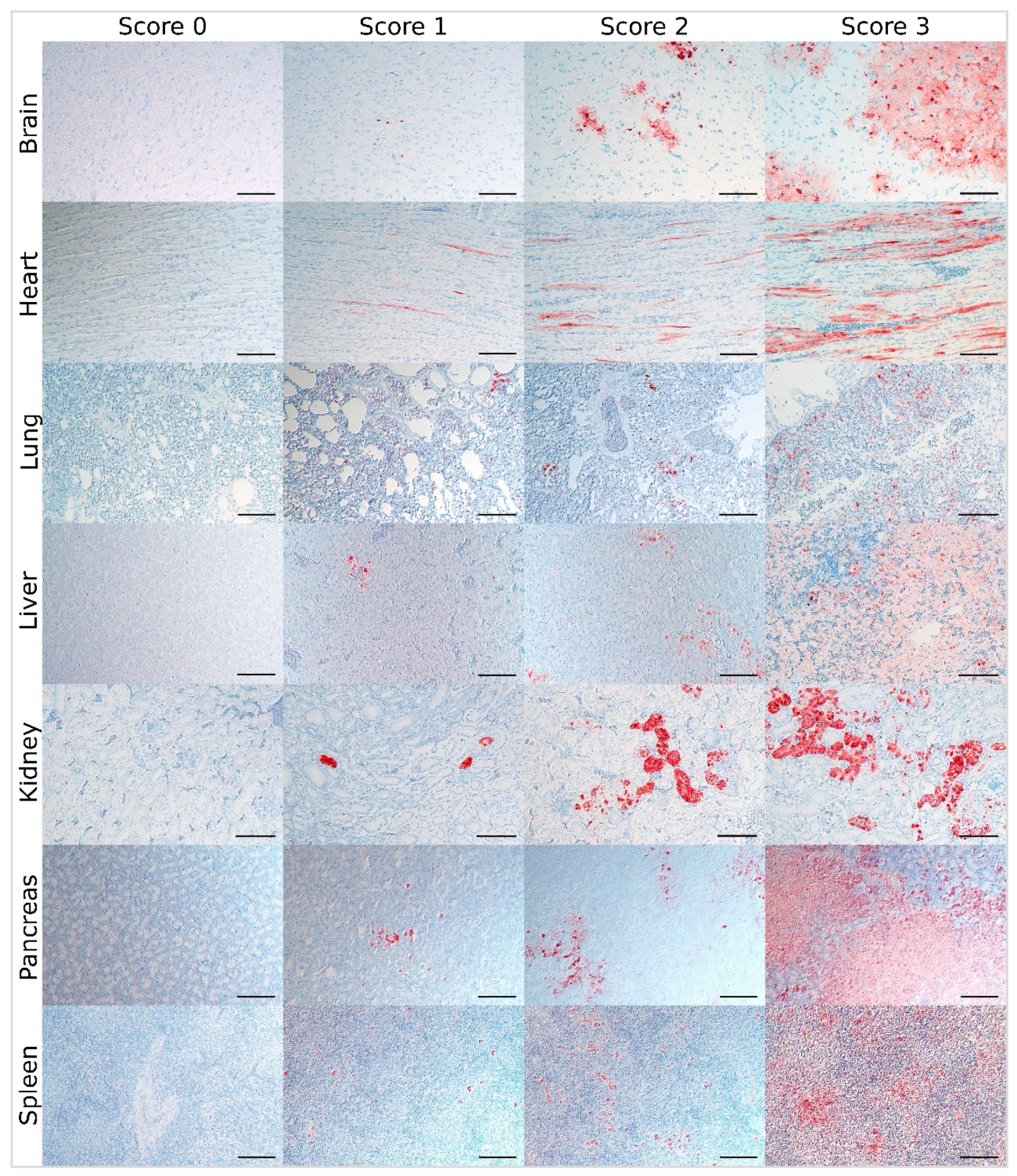
2.3. Development of a Semiquantitative Scoring System
2.4. Comparative Meta-Analysis between Selected Studies
2.5. Quantitative Real-Time RT-PCR
2.6. Statistical Analysis
3. Results
3.1. Lesion Score: Histopathologic Scoring for the Grade of AIV-Induced Lesions
3.2. Virus Score: Immunohistochemical Scoring for the Distribution of AIV Antigen
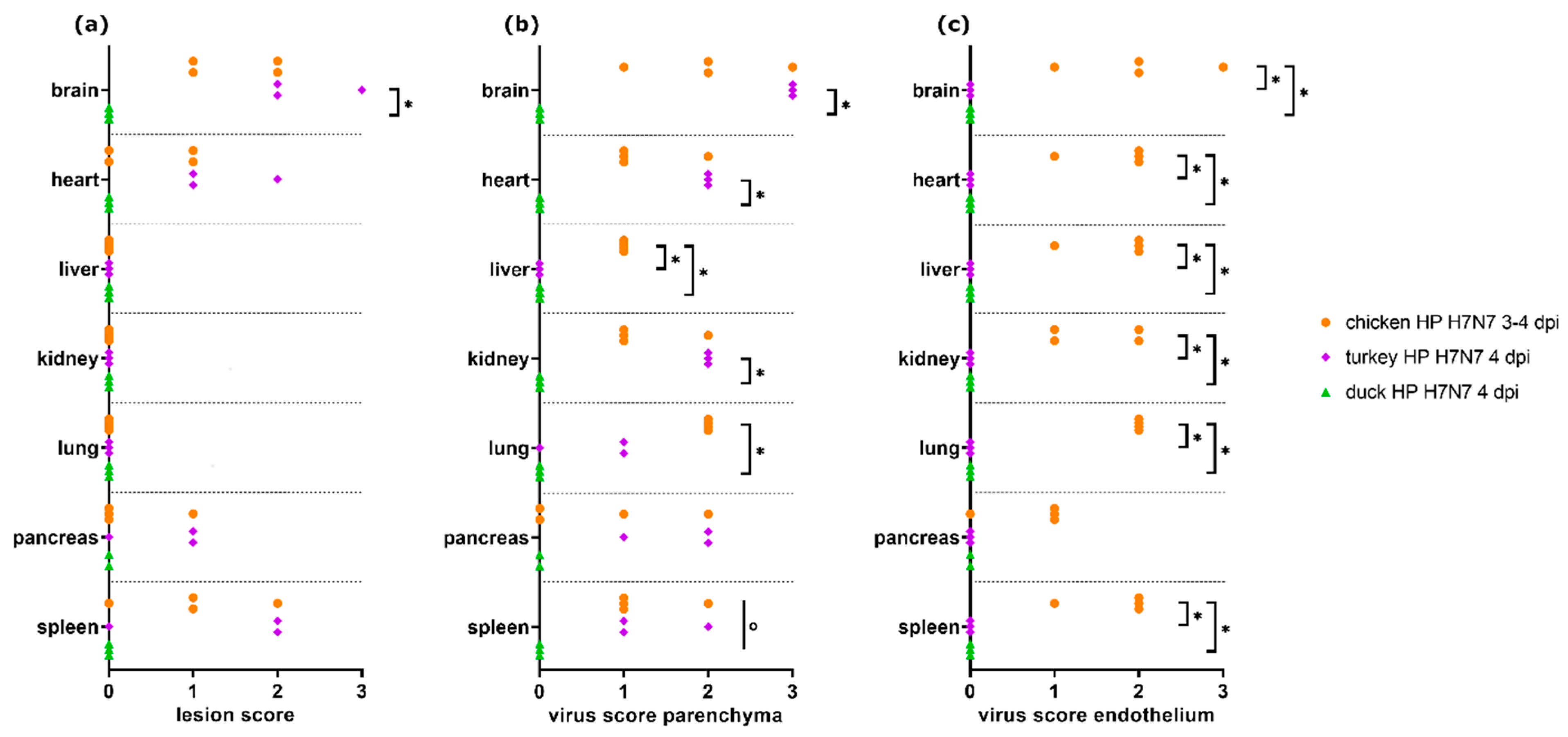
3.3. Comparison of H7N7-Infected Chickens, Turkeys, and Ducks
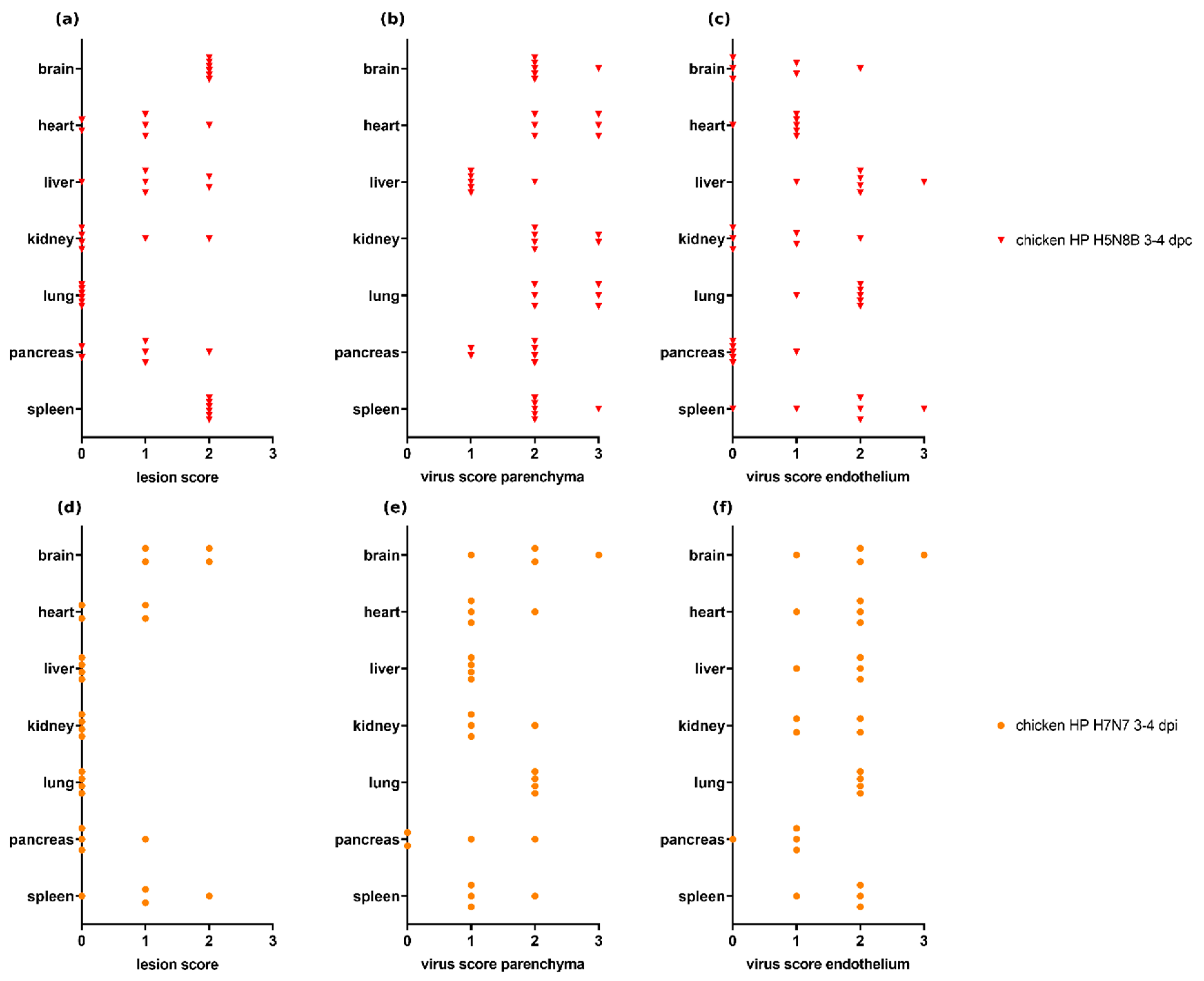
3.4. Scoring of H7N7- and H5N8B-Infected Chickens
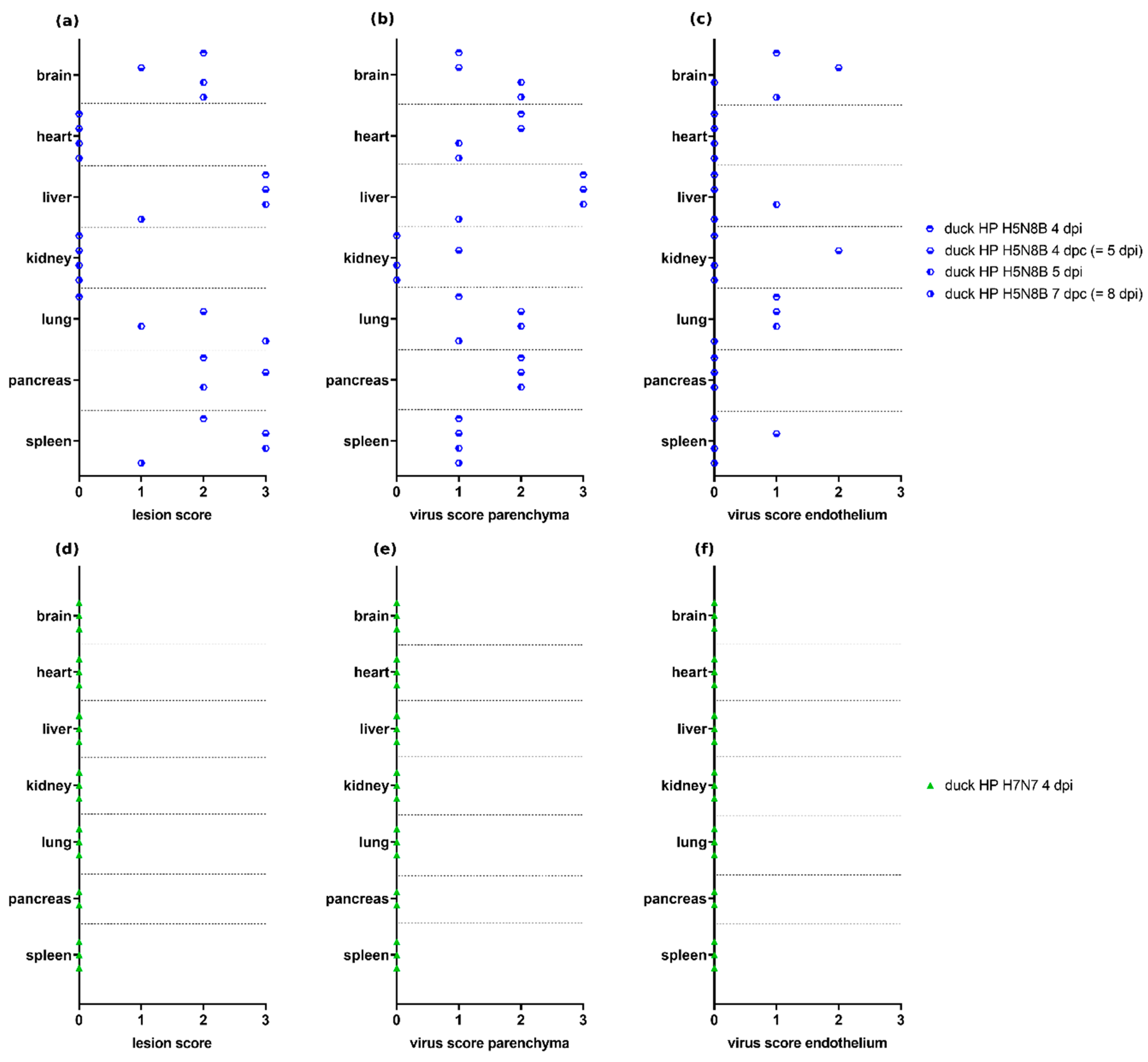
3.5. Scoring of H7N7- and H5N8B-Infected Ducks
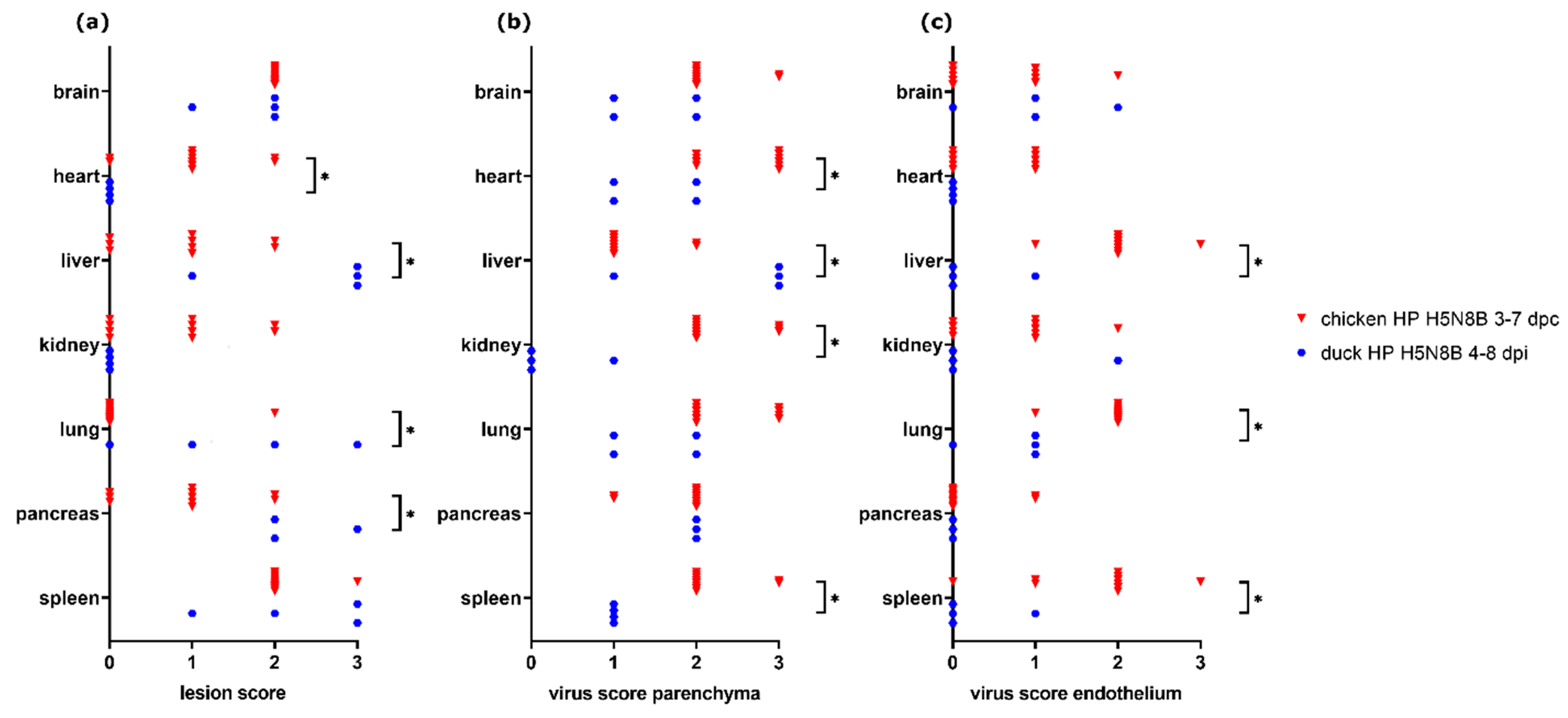
3.6. Comparison of H5N8B-Infected Chickens and Ducks
3.7. Comparison of Different Time Points within H7N7-Infected Chickens
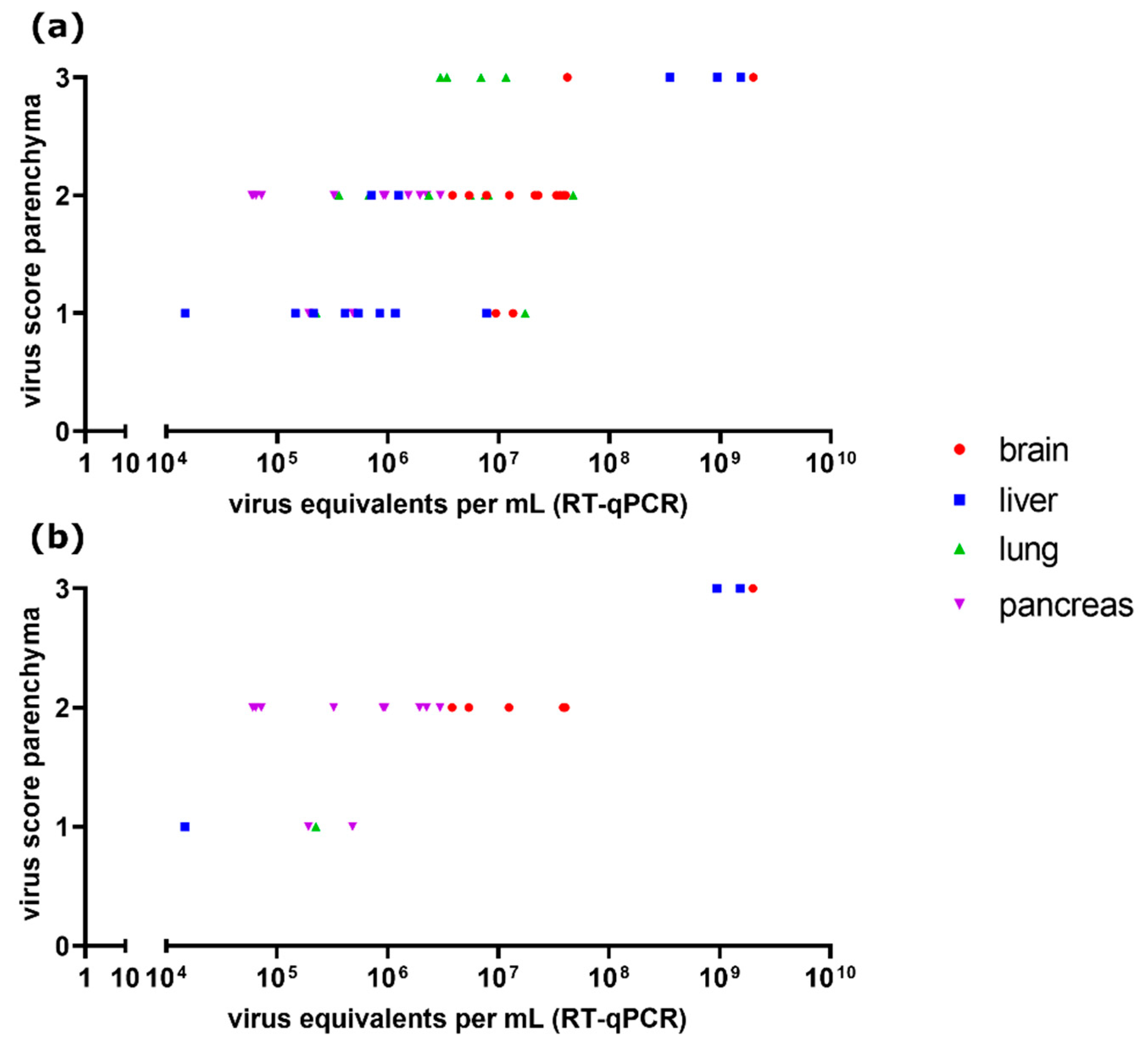
3.8. Comparison of Parenchyma Antigen Score and Virus Quantification
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medina, R.A.; García-Sastre, A. Influenza A viruses: New research developments. Nat. Rev. Microbiol. 2011, 9, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, E.C. Influenza Virus. Trends Microbiol. 2018, 26, 809–810. [Google Scholar] [CrossRef]
- Mostafa, A.; Abdelwhab, E.M.; Mettenleiter, T.C.; Pleschka, S. Zoonotic potential of Influenza A viruses: A comprehensive overview. Viruses 2018, 10, 497. [Google Scholar] [CrossRef] [Green Version]
- Cattoli, G.; Milani, A.; Temperton, N.; Zecchin, B.; Buratin, A.; Molesti, E.; Aly, M.M.; Arafa, A.; Capua, I. Antigenic drift in H5N1 avian influenza virus in poultry is driven by mutations in major antigenic sites of the hemagglutinin molecule analogous to those for human influenza virus. J. Virol. 2011, 85, 8718–8724. [Google Scholar] [CrossRef] [Green Version]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef]
- Horimoto, T.; Kawaoka, Y. Influenza: Lessons from past pandemics, warnings from current incidents. Nat. Rev. Microbiol. 2005, 3, 591–600. [Google Scholar] [CrossRef]
- Jourdain, E.; van Riel, D.; Munster, V.J.; Kuiken, T.; Waldenström, J.; Olsen, B.; Ellström, P. The pattern of influenza virus attachment varies among wild bird species. PLoS ONE 2011, 6, e24155. [Google Scholar] [CrossRef] [Green Version]
- Wood, G.W.; McCauley, J.W.; Bashiruddin, J.B.; Alexander, D.J. Deduced amino acid sequences at the haemagglutinin cleavage site of avian influenza A viruses of H5 and H7 subtypes. Arch. Virol. 1993, 130, 209–217. [Google Scholar] [CrossRef]
- Steinhauer, D.A. Role of hemagglutinin cleavage for the pathogenicity of influenza virus. Virology 1999, 258, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Antarasena, C.; Sirimujalin, R.; Prommuang, P.; Blacksell, S.D.; Promkuntod, N.; Prommuang, P. Tissue tropism of a Thailand strain of high-pathogenicity avian influenza virus (H5N1) in tissues of naturally infected native chickens (Gallus gallus), Japanese quail (Coturnix coturnix japonica) and ducks (Anas spp.). Avian Pathol. 2006, 35, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Perkins, L.E.; Swayne, D.E. Pathobiology of A/chicken/Hong Kong/220/97 (H5N1) avian influenza virus in seven gallinaceous species. Vet. Pathol. 2001, 38, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-W.; Suarez, D.L.; Tumpey, T.M.; Sung, H.-W.; Kwon, Y.-K.; Lee, Y.-J.; Choi, J.-G.; Joh, S.-J.; Kim, M.-C.; Lee, E.-K.; et al. Characterization of highly pathogenic H5N1 avian influenza A viruses isolated from South Korea. J. Virol. 2005, 79, 3692–3702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swayne, D.E. Pathobiology of H5N2 Mexican avian influenza virus infections of chickens. Vet. Pathol. 1997, 34, 557–567. [Google Scholar] [CrossRef]
- Swayne, D.E. Understanding the complex pathobiology of high pathogenicity avian influenza viruses in birds. Avian Dis. 2007, 51, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Chaves, A.J.; Busquets, N.; Valle, R.; Rivas, R.; Vergara-Alert, J.; Dolz, R.; Ramis, A.; Darji, A.; Majó, N. Neuropathogenesis of a highly pathogenic avian influenza virus (H7N1) in experimentally infected chickens. Vet. Res. 2011, 42, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, Y.L.; Swayne, D.E. Comparative pathobiology of low and high pathogenicity H7N3 Chilean avian influenza viruses in chickens. Avian Dis. 2004, 48, 119–128. [Google Scholar] [CrossRef]
- van Riel, D.; van den Brand, J.M.A.; Munster, V.J.; Besteboer, T.M.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Pathology and virus distribution in chickens naturally infected with highly pathogenic avian influenza A virus (H7N7) during the 2003 outbreak in The Netherlands. Vet. Pathol. 2009, 46, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Bingham, J.; Green, D.J.; Lowther, S.; Klippel, J.; Burggraaf, S.; Anderson, D.E.; Wibawa, H.; Hoa, D.M.; Long, N.T.; Vu, P.P.; et al. Infection studies with two highly pathogenic avian influenza strains (Vietnamese and Indonesian) in Pekin ducks (Anas platyrhynchos), with particular reference to clinical disease, tissue tropism and viral shedding. Avian Pathol. 2009, 38, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.D.; Stallknecht, D.E.; Beck, J.R.; Suarez, D.L.; Swayne, D.E. Susceptibility of North American ducks and gulls to H5N1 highly pathogenic avian influenza viruses. Emerg. Infect. Dis. 2006, 12, 1663–1670. [Google Scholar] [CrossRef]
- Perkins, L.E.L.; Swayne, D.E. Pathogenicity of a Hong Kong-origin H5N1 highly pathogenic avian influenza virus for emus, geese, ducks, and pigeons. Avian Dis. 2002, 46, 53–63. [Google Scholar] [CrossRef]
- Kwon, Y.K.; Thomas, C.; Swayne, D.E. Variability in pathobiology of South Korean H5N1 high-pathogenicity avian influenza virus infection for 5 species of migratory waterfowl. Vet. Pathol. 2010, 47, 495–506. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J.; Swayne, D.E. Pathobiology of Asian highly pathogenic avian influenza H5N1 virus infections in ducks. Avian Dis. 2007, 51, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Ellis, T.M.; Bousfield, R.B.; Bissett, L.A.; Dyrting, K.C.; Luk, G.S.M.; Tsim, S.T.; Sturm-Ramirez, K.; Webster, R.G.; Guan, Y.; Malik Peiris, J.S. Investigation of outbreaks of highly pathogenic H5N1 avian influenza in waterfowl and wild birds in Hong Kong in late 2002. Avian Pathol. 2004, 33, 492–505. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.D.; Stallknecht, D.E.; Swayne, D.E. Experimental infection of swans and geese with highly pathogenic avian influenza virus (H5N1) of Asian lineage. Emerg. Infect. Dis. 2008, 14, 136–142. [Google Scholar] [CrossRef]
- Kalthoff, D.; Breithaupt, A.; Teifke, J.P.; Globig, A.; Harder, T.; Mettenleiter, T.C.; Beer, M. Highly pathogenic avian influenza virus (H5N1) in experimentally infected adult mute swans. Emerg. Infect. Dis. 2008, 14, 1267–1270. [Google Scholar] [CrossRef]
- Pasick, J.; Berhane, Y.; Embury-Hyatt, C.; Copps, J.; Kehler, H.; Handel, K.; Babiuk, S.; Hooper-McGrevy, K.; Li, Y.; Mai Le, Q.; et al. Susceptibility of Canada Geese (Branta canadensis) to highly pathogenic avian influenza virus (H5N1). Emerg. Infect. Dis. 2007, 13, 1821–1827. [Google Scholar] [CrossRef]
- Short, K.R.; Kroeze, E.J.B.V.; Reperant, L.A.; Richard, M.; Kuiken, T. Influenza virus and endothelial cells: A species specific relationship. Front. Microbiol. 2014, 5, 653:1–653:11. [Google Scholar] [CrossRef] [Green Version]
- Schafer, K.A.; Eighmy, J.; Fikes, J.D.; Halpern, W.G.; Hukkanen, R.R.; Long, G.G.; Meseck, E.K.; Patrick, D.J.; Thibodeau, M.S.; Wood, C.E.; et al. Use of severity grades to characterize histopathologic changes. Toxicol. Pathol. 2018, 46, 256–265. [Google Scholar] [CrossRef]
- Shackelford, C.; Long, G.G.; Wolf, J.; Okerberg, C.; Herbert, R. Qualitative and quantitative analysis of nonneoplastic lesions in toxicology studies. Toxicol. Pathol. 2002, 30, 93–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Brand, J.M.A.; Krone, O.; Wolf, P.U.; van de Bildt, M.W.G.; van Amerongen, G.; Osterhaus, A.D.M.E.; Kuiken, T. Host-specific exposure and fatal neurologic disease in wild raptors from highly pathogenic avian influenza virus H5N1 during the 2006 outbreak in Germany. Vet. Res. 2015, 46, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelwhab, E.-S.M.; Veits, J.; Breithaupt, A.; Gohrbandt, S.; Ziller, M.; Teifke, J.P.; Stech, J.; Mettenleiter, T.C. Prevalence of the C-terminal truncations of NS1 in avian influenza A viruses and effect on virulence and replication of a highly pathogenic H7N1 virus in chickens. Virulence 2016, 7, 546–557. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Horimoto, T.; Kawaoka, Y.; Alexander, D.J.; Itakura, C. Pathological studies of chickens experimentally infected with two highly pathogenic avian influenza viruses. Avian Pathol. 1996, 25, 285–304. [Google Scholar] [CrossRef] [Green Version]
- Klopfleisch, R.; Werner, O.; Mundt, E.; Harder, T.; Teifke, J.P. Neurotropism of highly pathogenic avian influenza virus A/chicken/Indonesia/2003 (H5N1) in experimentally infected pigeons (Columbia livia f. domestica). Vet. Pathol. 2006, 43, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Höfle, U.; van de Bildt, M.W.G.; Leijten, L.M.; van Amerongen, G.; Verhagen, J.H.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Tissue tropism and pathology of natural influenza virus infection in black-headed gulls (Chroicocephalus ridibundus). Avian Pathol. 2012, 41, 547–553. [Google Scholar] [CrossRef]
- Samir, M.; Vidal, R.O.; Abdallah, F.; Capece, V.; Seehusen, F.; Geffers, R.; Hussein, A.; Ali, A.A.H.; Bonn, S.; Pessler, F. Organ-specific small non-coding RNA responses in domestic (Sudani) ducks experimentally infected with highly pathogenic avian influenza virus (H5N1). RNA Biol. 2020, 17, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Gibson-Corley, K.N.; Olivier, A.K.; Meyerholz, D.K. Principles for valid histopathologic scoring in research. Vet. Pathol. 2013, 50, 1007–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klopfleisch, R. Multiparametric and semiquantitative scoring systems for the evaluation of mouse model histopathology—A systematic review. BMC Vet. Res. 2013, 9, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrie, A.; Watson, P. Statistics for Veterinary and Animal Science, 3rd ed.; Wiley-Blackwell: Oxford, UK, 2013; pp. 165–166, 171–173, 336. ISBN 0470670754. [Google Scholar]
- Blaurock, C.; Scheibner, D.; Landmann, M.; Vallbracht, M.; Ulrich, R.; Böttcher-Friebertshäuser, E.; Mettenleiter, T.C.; Abdelwhab, E.M. Non-basic amino acids in the hemagglutinin proteolytic cleavage site of a European H9N2 avian influenza virus modulate virulence in turkeys. Sci. Rep. 2020, 10, 21226. [Google Scholar] [CrossRef]
- Gischke, M.; Ulrich, R.; Fatola, O.I.; Scheibner, D.; Salaheldin, A.H.; Crossley, B.; Böttcher-Friebertshäuser, E.; Veits, J.; Mettenleiter, T.C.; Abdelwhab, E.M. Insertion of basic amino acids in the hemagglutinin cleavage site of H4N2 avian influenza virus (AIV)-reduced virus fitness in chickens is restored by reassortment with highly pathogenic H5N1 AIV. Int. J. Mol. Sci. 2020, 21, 2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graaf, A.; Ulrich, R.; Maksimov, P.; Scheibner, D.; Koethe, S.; Abdelwhab, E.M.; Mettenleiter, T.C.; Beer, M.; Harder, T. A viral race for primacy: Co-infection of a natural pair of low and highly pathogenic H7N7 avian influenza viruses in chickens and embryonated chicken eggs. Emerg. Microbes Infect. 2018, 7, 204:1–204:12. [Google Scholar] [CrossRef] [Green Version]
- Grund, C.; Hoffmann, D.; Ulrich, R.; Naguib, M.M.; Schinköthe, J.; Hoffmann, B.; Harder, T.; Saenger, S.; Zscheppang, K.; Tönnies, M.; et al. A novel European H5N8 influenza A virus has increased virulence in ducks but low zoonotic potential. Emerg. Microbes Infect. 2018, 7, 132:1–132:14. [Google Scholar] [CrossRef]
- Koethe, S.; Ulrich, L.; Ulrich, R.; Amler, S.; Graaf, A.; Harder, T.C.; Grund, C.; Mettenleiter, T.C.; Conraths, F.J.; Beer, M.; et al. Modulation of lethal HPAIV H5N8 clade 2.3.4.4B infection in AIV pre-exposed mallards. Emerg. Microbes Infect. 2020, 9, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Scheibner, D.; Ulrich, R.; Fatola, O.I.; Graaf, A.; Gischke, M.; Salaheldin, A.H.; Harder, T.C.; Veits, J.; Mettenleiter, T.C.; Abdelwhab, E.M. Variable impact of the hemagglutinin polybasic cleavage site on virulence and pathogenesis of avian influenza H7N7 virus in chickens, turkeys and ducks. Sci. Rep. 2019, 9, 11556:1–11556:13. [Google Scholar] [CrossRef] [PubMed]
- Naguib, M.M.; Ulrich, R.; Kasbohm, E.; Eng, C.L.P.; Hoffmann, D.; Grund, C.; Beer, M.; Harder, T.C. Natural reassortants of potentially zoonotic avian influenza viruses H5N1 and H9N2 from Egypt display distinct pathogenic phenotypes in experimentally infected chickens and ferrets. J. Virol. 2017, 91, e01300-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvin, R.; Schinkoethe, J.; Grund, C.; Ulrich, R.; Bönte, F.; Behr, K.P.; Voss, M.; Samad, M.A.; Hassan, K.E.; Luttermann, C.; et al. Comparison of pathogenicity of subtype H9 avian influenza wild-type viruses from a wide geographic origin expressing mono-, di-, or tri-basic hemagglutinin cleavage sites. Vet. Res. 2020, 51, 48. [Google Scholar] [CrossRef] [Green Version]
- Krone, O.; Globig, A.; Ulrich, R.; Harder, T.; Schinköthe, J.; Herrmann, C.; Gerst, S.; Conraths, F.J.; Beer, M. White-tailed sea eagle (Haliaeetus albicilla) die-off due to infection with highly pathogenic avian influenza virus, subtype H5N8, in Germany. Viruses 2018, 10, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breithaupt, A.; Kalthoff, D.; Dale, J.; Bairlein, F.; Beer, M.; Teifke, J.P. Neurotropism in blackcaps (Sylvia atricapilla) and red-billed queleas (Quelea quelea) after highly pathogenic avian influenza virus H5N1 infection. Vet. Pathol. 2011, 48, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Dietze, K.; Graaf, A.; Homeier-Bachmann, T.; Grund, C.; Forth, L.; Pohlmann, A.; Jeske, C.; Wintermann, M.; Beer, M.; Conraths, F.J.; et al. From low to high pathogenicity-Characterization of H7N7 avian influenza viruses in two epidemiologically linked outbreaks. Transbound. Emerg. Dis. 2018, 65, 1576–1587. [Google Scholar] [CrossRef]
- Hoffmann, B.; Harder, T.; Lange, E.; Kalthoff, D.; Reimann, I.; Grund, C.; Oehme, R.; Vahlenkamp, T.W.; Beer, M. New real-time reverse transcriptase polymerase chain reactions facilitate detection and differentiation of novel A/H1N1 influenza virus in porcine and human samples. Berl. Munch. Tierarztl. Wochenschr. 2010, 123, 286–292. [Google Scholar] [CrossRef]
- Meyerholz, D.K.; Beck, A.P. Principles and approaches for reproducible scoring of tissue stains in research. Lab. Investig. 2018, 98, 844–855. [Google Scholar] [CrossRef]
- Brisson, A.R.; Matsui, D.; Rieder, M.J.; Fraser, D.D. Translational research in pediatrics: Tissue sampling and biobanking. Pediatrics 2012, 129, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Kayser, K.; Schultz, H.; Goldmann, T.; Görtler, J.; Kayser, G.; Vollmer, E. Theory of sampling and its application in tissue based diagnosis. Diagn. Pathol. 2009, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Pantin-Jackwood, M.J.; Swayne, D.E. Pathogenesis and pathobiology of avian influenza virus infection in birds. Rev. Sci. Tech. 2009, 28, 113–136. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J.; Stephens, C.B.; Bertran, K.; Swayne, D.E.; Spackman, E. The pathogenesis of H7N8 low and highly pathogenic avian influenza viruses from the United States 2016 outbreak in chickens, turkeys and mallards. PLoS ONE 2017, 12, e0177265. [Google Scholar] [CrossRef]
- Löndt, B.Z.; Nunez, A.; Banks, J.; Nili, H.; Johnson, L.K.; Alexander, D.J. Pathogenesis of highly pathogenic avian influenza A/turkey/Turkey/1/2005 H5N1 in Pekin ducks (Anas platyrhynchos) infected experimentally. Avian Pathol. 2008, 37, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Perkins, L.E.L.; Swayne, D.E. Comparative susceptibility of selected avian and mammalian species to a Hong Kong-origin H5N1 high-pathogenicity avian influenza virus. Avian Dis. 2003, 47, 956–967. [Google Scholar] [CrossRef]
- Kim, H.-R.; Kwon, Y.-K.; Jang, I.; Lee, Y.-J.; Kang, H.-M.; Lee, E.-K.; Song, B.-M.; Lee, H.-S.; Joo, Y.-S.; Lee, K.-H.; et al. Pathologic changes in wild birds infected with highly pathogenic avian influenza A(H5N8) viruses, South Korea, 2014. Emerg. Infect. Dis. 2015, 21, 775–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spackman, E.; Pantin-Jackwood, M.J.; Kapczynski, D.R.; Swayne, D.E.; Suarez, D.L. H5N2 Highly Pathogenic Avian Influenza Viruses from the US 2014-2015 outbreak have an unusually long pre-clinical period in turkeys. BMC Vet. Res. 2016, 12, 260:1–260:9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelissen, J.B.W.J.; Vervelde, L.; Post, J.; Rebel, J.M.J. Differences in highly pathogenic avian influenza viral pathogenesis and associated early inflammatory response in chickens and ducks. Avian Pathol. 2013, 42, 347–364. [Google Scholar] [CrossRef] [Green Version]
- van den Brand, J.M.A.; Verhagen, J.H.; Veldhuis Kroeze, E.J.B.; van de Bildt, M.W.G.; Bodewes, R.; Herfst, S.; Richard, M.; Lexmond, P.; Bestebroer, T.M.; Fouchier, R.A.M.; et al. Wild ducks excrete highly pathogenic avian influenza virus H5N8 (2014-2015) without clinical or pathological evidence of disease. Emerg. Microbes Infect. 2018, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Berhane, Y.; Kobasa, D.; Embury-Hyatt, C.; Pickering, B.; Babiuk, S.; Joseph, T.; Bowes, V.; Suderman, M.; Leung, A.; Cottam-Birt, C.; et al. Pathobiological characterization of a novel reassortant highly pathogenic H5N1 virus isolated in British Columbia, Canada, 2015. Sci. Rep. 2016, 6, 23380:1–23380:15. [Google Scholar] [CrossRef] [Green Version]
- Pantin-Jackwood, M.J.; Costa-Hurtado, M.; Shepherd, E.; DeJesus, E.; Smith, D.; Spackman, E.; Kapczynski, D.R.; Suarez, D.L.; Stallknecht, D.E.; Swayne, D.E. Pathogenicity and transmission of H5 and H7 highly pathogenic avian influenza viruses in mallards. J. Virol. 2016, 90, 9967–9982. [Google Scholar] [CrossRef] [Green Version]
- Pantin-Jackwood, M.J.; Costa-Hurtado, M.; Bertran, K.; DeJesus, E.; Smith, D.; Swayne, D.E. Infectivity, transmission and pathogenicity of H5 highly pathogenic avian influenza clade 2.3.4.4 (H5N8 and H5N2) United States index viruses in Pekin ducks and Chinese geese. Vet. Res. 2017, 48, 33:1–33:14. [Google Scholar] [CrossRef] [Green Version]
- Pantin-Jackwood, M.; Swayne, D.E.; Smith, D.; Shepherd, E. Effect of species, breed and route of virus inoculation on the pathogenicity of H5N1 highly pathogenic influenza (HPAI) viruses in domestic ducks. Vet. Res. 2013, 44, 62:1–62:11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Deng, G.; Ma, S.; Zeng, X.; Yin, X.; Li, M.; Zhang, B.; Cui, P.; Chen, Y.; Yang, H.; et al. Rapid evolution of H7N9 highly pathogenic viruses that emerged in China in 2017. Cell Host Microbe 2018, 24, 558–568. [Google Scholar] [CrossRef] [Green Version]
- Capua, I.; Mutinelli, F. Mortality in Muscovy ducks (Cairina moschata) and domestic geese (Anser anser var. domestica) associated with natural infection with a highly pathogenic avian influenza virus of H7N1 subtype. Avian Pathol. 2001, 30, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Aldous, E.W.; Seekings, J.M.; McNally, A.; Nili, H.; Fuller, C.M.; Irvine, R.M.; Alexander, D.J.; Brown, I.H. Infection dynamics of highly pathogenic avian influenza and virulent avian paramyxovirus type 1 viruses in chickens, turkeys and ducks. Avian Pathol. 2010, 39, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Pantin-Jackwood, M.J.; Suarez, D.L.; Spackman, E.; Swayne, D.E. Age at infection affects the pathogenicity of Asian highly pathogenic avian influenza H5N1 viruses in ducks. Virus Res. 2007, 130, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Wasilenko, J.L.; Arafa, A.M.; Selim, A.A.; Hassan, M.K.; Aly, M.M.; Ali, A.; Nassif, S.; Elebiary, E.; Balish, A.; Klimov, A.; et al. Pathogenicity of two Egyptian H5N1 highly pathogenic avian influenza viruses in domestic ducks. Arch. Virol. 2011, 156, 37–51. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Score 0 | Score 1 | Score 2 | Score 3 |
|---|---|---|---|---|
| all | no | mild | moderate | severe |
| parenchymal necrotic/necrotizing inflammatory lesions | ||||
| brain | no | mild | moderate | severe |
| necrotizing polioencephalitis/meningoencephalitis with/without gliosis and with/without perivascular lymphohistiocytic infiltration | ||||
| heart | no | mild | moderate | severe |
| necrotizing myocarditis/myocardial necrosis with/without lymphohistiocytic infiltration | ||||
| lung | no | mild | moderate | severe |
| (fibrino)necrotizing (broncho-)pneumonia/necrosis of parenchymal cells | ||||
| liver | no | mild | moderate | severe |
| necrotizing hepatitis/hepatocellular necrosis with/without lobular/perivascular pattern and with/without lymphohistiocytic infiltration | ||||
| kidney | no | mild | moderate | severe |
| necrosis of tubular epithelium with/without lymphohistiocytic infiltration | ||||
| pancreas | no | mild | moderate | severe |
| necrotizing pancreatitis/pancreatic necrosis | ||||
| spleen | no | mild | moderate | severe |
| necrosis/apoptosis of lymphoid and/or reticuloendothelial cells and/or lymphoid depletion | ||||
| Organ | Parenchymal Cells |
|---|---|
| all | functional cells of organ (e.g., epithelia) |
| brain | neurons, glial cells |
| heart | cardiomyocytes |
| lung | bronchiolar and parabronchiolar epithelia, pneumocytes, alveolar macrophages * |
| liver | hepatocytes |
| kidney | tubular epithelium |
| pancreas | exocrine pancreatocytes |
| spleen | reticular cells, round cells (lymphocytes, dendritic cells, macrophages) |
| Organ | Score 0 | Score 1 | Score 2 | Score 3 |
|---|---|---|---|---|
| all | no | focal to oligofocal | multifocal | coalescing to diffuse |
| antigen in parenchymal cells/necrotic parenchymal areas | ||||
| further criteria for orientation and classification of inconclusive cases | ||||
| brain | - | less than 5% immunoreactive area and up to 0.75 clusters/low-power field * | less than 5% immunoreactive area and more than 0.75 and up to 5 clusters/low-power field * or 5%–20% immunoreactive area and up to 5 clusters/low-power field * | more than 20% immunoreactive area and/or more than 5 clusters/low-power field * |
| heart | - | less than 10% immunoreactive area and less than 2 immunoreactive cells/ high-power field * | less than 10% immunoreactive area and 2–10 immunoreactive cells/ high-power field * | at least 10% immunoreactive area and/or more than 10 immunoreactive cells/ high-power field * |
| lung | - | less than 5% immunoreactive area and less than 3 immunoreactive cells/ high-power field * | less than 5% immunoreactive area and at least 3 immunoreactive cells/ high-power field * | at least 5% immunoreactive area |
| liver | ||||
| - necrotic areas immunoreactive | - | less than 5% immunoreactive area | 5%–14% immunoreactive area | at least 15% immunoreactive area |
| - mainly single cells immunoreactive | - | less than 3 immunoreactive cells/ high-power field * | 3–10 immunoreactive cells/ high-power field * | more than 10 immunoreactive cells/ high-power field * |
| kidney | - | less than 2 clusters/ low-power field * and up to 2% immunoreactive area | at least 2 clusters/ low-power field * and up to 5% immunoreactive area with single clusters | 5% immunoreactive area with coalescing clusters or more than 5% immunoreactive area |
| pancreas | - | less than 5% immunoreactive area and up to 1 cluster/ low-power field * | less than 5% immunoreactive area and more than 1 cluster/ low-power field * or 5%–24% immunoreactive area | at least 25% immunoreactive area |
| spleen | ||||
| - necrotic areas immunoreactive | - | less than 2% immunoreactive area | 2%–14% immunoreactive area | at least 15% immunoreactive area |
| - mainly single cells immunoreactive | - | up to 15 immunoreactive cells/ high-power field * | 16–40 immunoreactive cells/ high-power field * | more than 40 immunoreactive cells/ high-power field * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landmann, M.; Scheibner, D.; Graaf, A.; Gischke, M.; Koethe, S.; Fatola, O.I.; Raddatz, B.; Mettenleiter, T.C.; Beer, M.; Grund, C.; et al. A Semiquantitative Scoring System for Histopathological and Immunohistochemical Assessment of Lesions and Tissue Tropism in Avian Influenza. Viruses 2021, 13, 868. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050868
Landmann M, Scheibner D, Graaf A, Gischke M, Koethe S, Fatola OI, Raddatz B, Mettenleiter TC, Beer M, Grund C, et al. A Semiquantitative Scoring System for Histopathological and Immunohistochemical Assessment of Lesions and Tissue Tropism in Avian Influenza. Viruses. 2021; 13(5):868. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050868
Chicago/Turabian StyleLandmann, Maria, David Scheibner, Annika Graaf, Marcel Gischke, Susanne Koethe, Olanrewaju I. Fatola, Barbara Raddatz, Thomas C. Mettenleiter, Martin Beer, Christian Grund, and et al. 2021. "A Semiquantitative Scoring System for Histopathological and Immunohistochemical Assessment of Lesions and Tissue Tropism in Avian Influenza" Viruses 13, no. 5: 868. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050868