
Influence of SARS-CoV-2 Variant B.1.1.7, Vaccination, and Public Health Measures on the Spread of SARS-CoV-2

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Statistical Model
2.2. Study Population
2.3. SARS-CoV-2 RNA Detection and Sequencing
3. Results
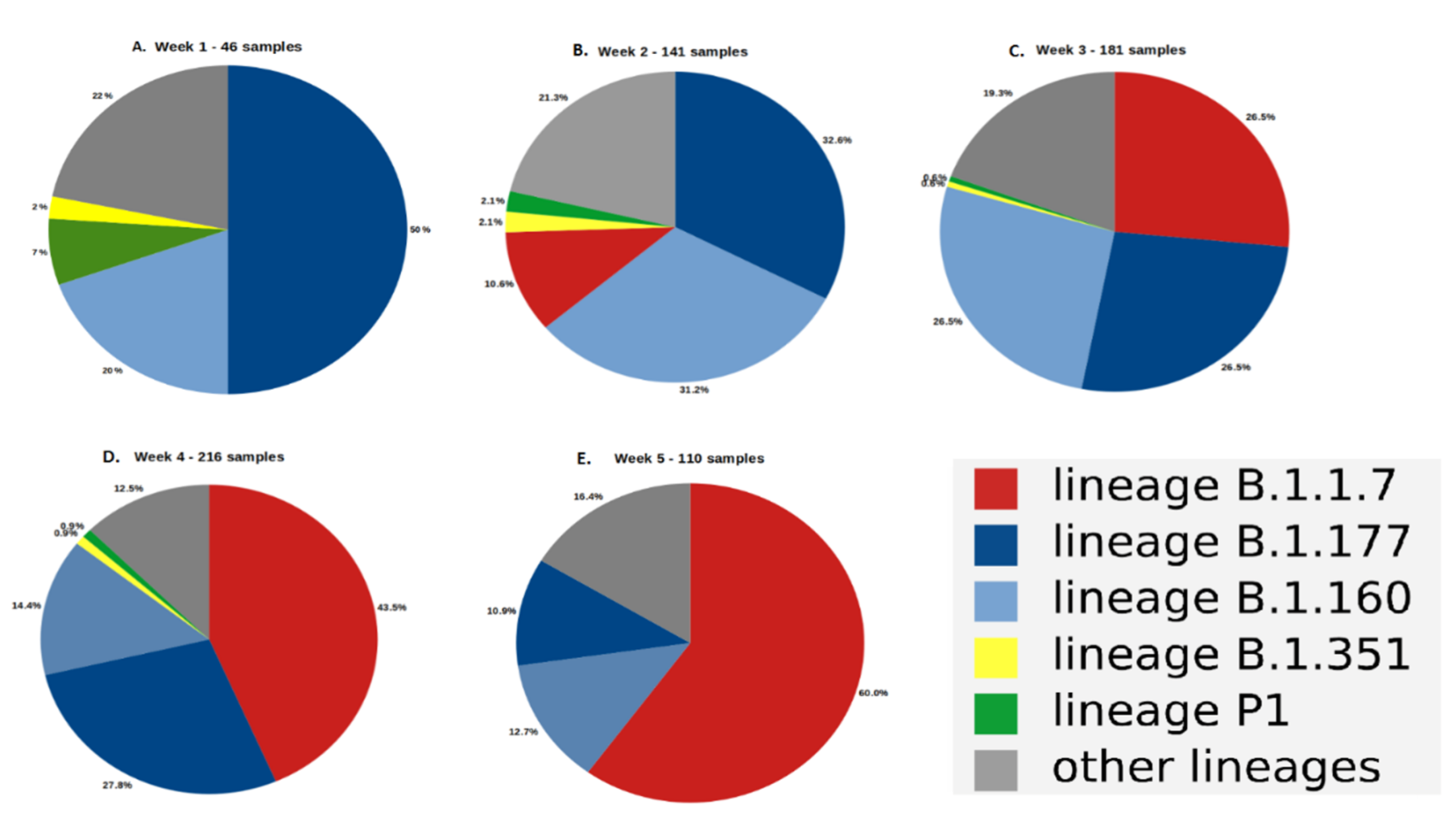
3.1. Evolution of the Proportion of the SARS-CoV-2 Variants
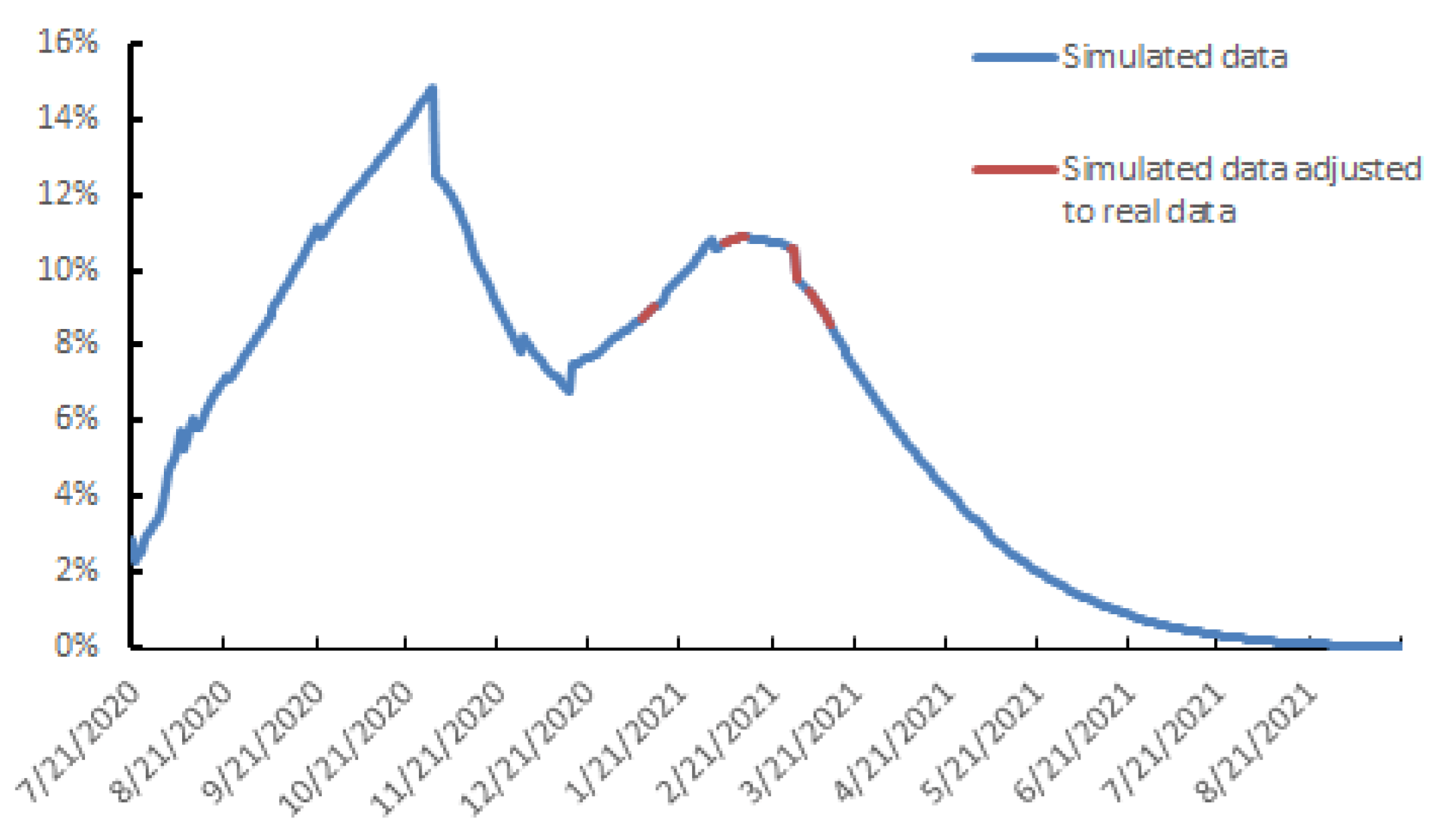
3.2. Outlook for the Evolution of the SARS-CoV-2 Dynamics in the Urban Area of Toulouse
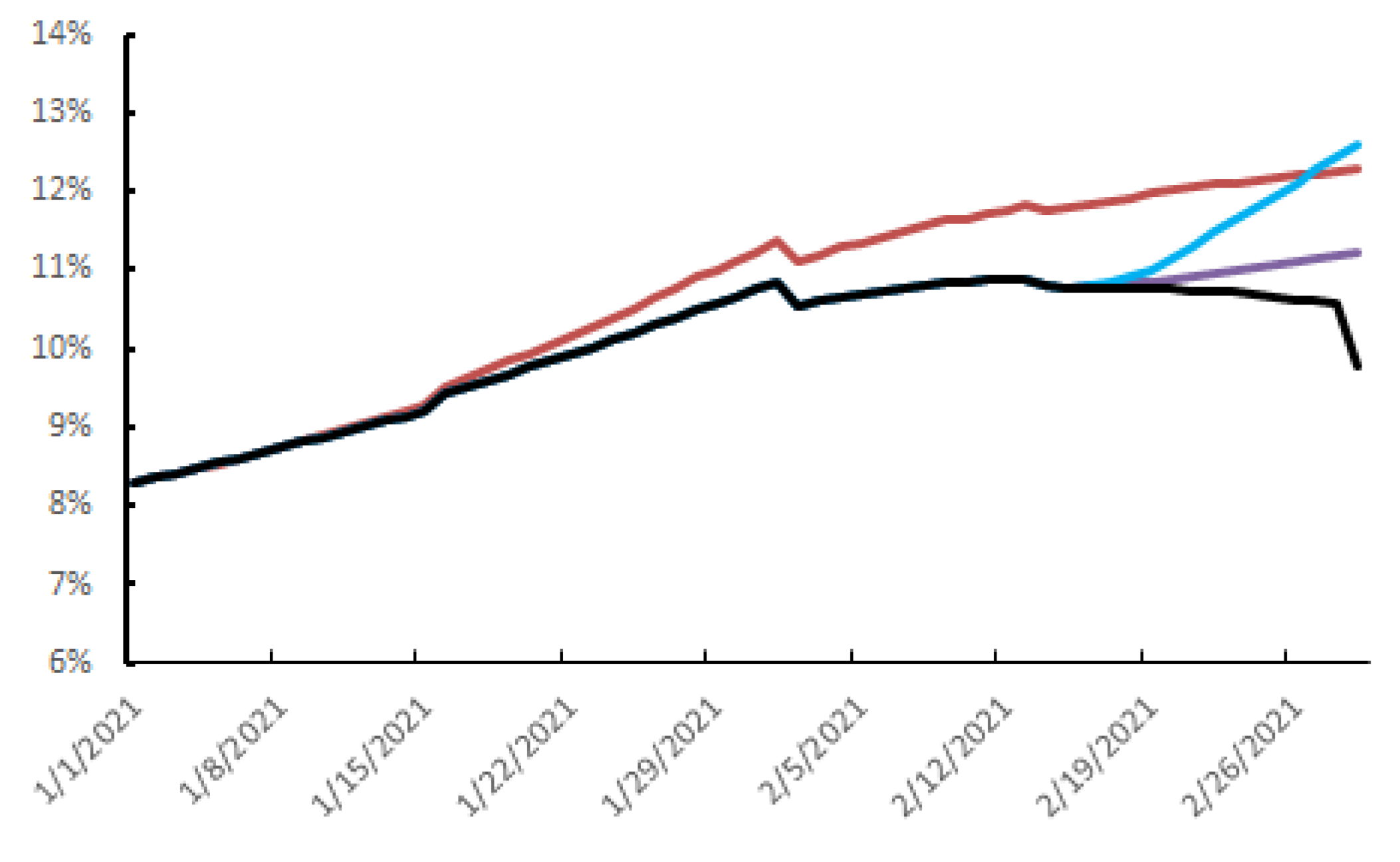
3.3. The Parameters Influencing the Spread of SARS-CoV-2
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perlman, S. Another Decade, Another Coronavirus. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- WHO Virtual press conference on COVID-19—11 March 2020. Available online: https://www.who.int/docs/default-source/coronaviruse/transcripts/who-audio-emergencies-coronavirus-press-conference-full-and-final-11mar2020.pdf?sfvrsn=cb432bb3_2 (accessed on 26 April 2021).
- Dimeglio, C.; Loubes, J.M.; Mansuy, J.M.; Izopet, J. Quantifying the impact of public health protection measures on the spread of SARS-CoV-2. J. Infect. 2021, 82, 414–451. [Google Scholar] [CrossRef]
- EU Strategy for COVID-19 Vaccines. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?qid=1597339415327&uri=CELEX:52020DC0245 (accessed on 26 April 2021).
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomized controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 9, 99–111. [Google Scholar] [CrossRef]
- Baden, L.R.; Rubin, E.J. Covid-19—The Search for Effective Therapy. N. Engl. J. Med. 2020, 382, 1851–1852. [Google Scholar] [CrossRef] [PubMed]
- Emary, K.R.W.; Golubchik, T.; Aley, P.K.; Ariani, C.V.; Angus, B.J.; Bibi, S.; Blane, B.; Bonsall, D.; Cicconi, P.; Charlton, S.; et al. Efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 VOC 202012/01 (B.1.1.7). Lancet 2021, 397, 1351–1362. [Google Scholar] [CrossRef]
- Volz, E.; Mishra, S.; Chand, M.; Barrett, J.C.; Johnson, R.; Geidelberg, L.; Hinsley, W.R.; Laydon, D.J.; Darbrera, G.; O’Toole, A.; et al. Transmission of SARS-CoV-2 Lineage, B.1.1.7 in England: Insights from linking epidemiological and genetic data. medRxiv 2021. [Google Scholar] [CrossRef]
- Kirby, T. New variant of SARS-CoV-2 in IK causes surge of COVID-19. Lancet Respir. Med. 2021, 9, e20–e21. [Google Scholar] [CrossRef]
- Leung, K.; Shum, M.H.; Leung, G.M.; Lam, T.T.; Wu, J.T. Early transmissibility assessment of the N501Y mutant strains of SARS-CoV-2 in the United Kingdom, October to November 2020. Euro Surveill. 2021, 26, 2002106. [Google Scholar] [CrossRef]
- Wu, J.T.; Leung, K. Nowcasting and forecasting the potential domestic and international spread of the 2019-ncov outbreak originating in wuhan, china: A modelling study. Lancet 2020, 395, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.M.; Rui, J. A mathematical model for simulating the phase-based transmissibility of a novel coronavirus. Infect. Dis. Poverty 2020, 9, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kermack, W.O.; McKendrick, A.G. A Contribution to the Mathematical Theory of Epidemics. Proc. Roy. Soc. Lond. A 1927, 115, 700–721. [Google Scholar]
- Cooper, I.; Mondal, A. Antonopoulos CG. A SIR model assumption for the spread of COVID-19 in different communities. Chaos Solitons Fractals 2020, 139, 110057. [Google Scholar] [CrossRef] [PubMed]
- Estrada, E. COVID-19 and SARS-CoV-2. Modeling the present, looking at the future. Phys. Rep. 2020, 10, 1–51. [Google Scholar]
- Walsh, K.A.; Jordan, K.; Clyne, B.; Rohde, D.; Drummond, L.; Byrne, P.; Ahern, S.; Carty, P.G.; O’Brien, K.K.; O’Murchu, E.; et al. SARS-CoV-2 detection, viral load and infectivity over the course of an infection. J. Infect. 2020, 81, 357–371. [Google Scholar] [CrossRef]
- Lescure, F.X.; Bouadma, L.; Nguyen, D.; Parisey, M.; Wicky, P.-H.; Behillil, S.; Gaymard, A.; Bouscambert-Duchamp, M.; Donati, F.; Le Hingrat, Q.; et al. Clinical and virological data of the first cases of COVID-19 in Europe: A case series. Lancet Infect. Dis. 2020, 20, 697–706. [Google Scholar] [CrossRef] [Green Version]
- Jaafar, R.; Aherfi, S.; Wurtz, N.; Grimaldier, C.; Hoang, V.T.; Colson, P.; Raoult, D.; La Scola, B. Correlation between 3790 qPCR positives samples and positive cell cultures including 1941 SARS-CoV-2 isolates. Clin. Infect. Dis. 2020, 28, ciaa1491. [Google Scholar] [CrossRef]
- Dimeglio, C.; Loubes, J.-M.; Deporte, B.; Dubois, M.; Latour, J.; Mansuy, J.-M.; Izopet, J. The SARS-CoV-2 seroprevalence is the key factor for deconfinement in France. J. Infect. 2020, 81, 318–356. [Google Scholar] [CrossRef]
- Santé Publique France (2021) COVID-19: Epidemiological update in Occitania on 11 March 2021. Available online: https://www.santepubliquefrance.fr/regions/occitanie/documents/bulletin-regional/2021/covid-19-point-epidemiologique-en-occitanie-du-11-mars-2021 (accessed on 26 April 2021).
- Salje, H.; Tran Kiem, C.; Lefrancq, N.; Courtejoie, N.; Bosetti, P.; Paireau, J.; Andronico, A.; Hozé, N.; Richet, J.; Dubost, C.-L.; et al. Estimating the burden of SARS-CoV-2 in France. Science 2020. [Google Scholar] [CrossRef]
- WHO Report of the WHO-China Joint Mission on Coronavirus Disease. (COVID-19). Available online: https://www.who.int/docs/default-source/coronaviruse/who-china-joint-mission-on-Covid-19-final-report.pdf (accessed on 26 April 2021).
- WHO SARS-CoV-2 Variant—United Kingdom of Great Britain and Northern Ireland. Disease Outbreak News 21 December 2020. Available online: https://www.who.int/csr/don/21-december-2020-sars-cov2-variant-united-kingdom/en/ (accessed on 26 April 2021).
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England. Science 2021, 9. [Google Scholar] [CrossRef]
- INSERM Press Release (October 2020). Available online: https://presse.inserm.fr/premiers-resultats-des-enquetes-de-sante-publique-de-linserm-sur-la-Covid-19-facteurs-de-risque-individuels-et-sociaux/41032/ (accessed on 26 April 2021).
- Buitrago-Garcia, D.; Egli-Gany, D.; Counotte, M.J.; Hossmann, S.; Imeri, H.; Ipekci, A.M.; Salanti, G.; Low, N. Occurrence and transmission potential of asymptomatic and presymptomatic SARS-CoV-2 infections: A living systematic review and meta-analysis. PLoS Med. 2020, 17, e1003346. [Google Scholar] [CrossRef] [PubMed]
- Meyerowitz, E.A.; Richterman, A.; Bogoch, I.I.; Low, N.; Cevik, M. Towards an accurate and systematic characterisation of persistently asymptomatic infection with SARS-CoV-2. Lancet Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.S.; Msomi, N.; et al. Emergence of a SARS-CoV-2 variant of concern with mutations in spike glycoprotein. Nature 2021. [Google Scholar] [CrossRef] [PubMed]
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, D.d.S.; Mishra, S.; Crispim, M.A.E.; Sales, F.C.; Hawryluk, I.; McCrone, J.T.; et al. Genomics and epidemiology of a novel SARS-CoV-2 lineage in Manaus, Brazil. medRxiv 2021. [Google Scholar] [CrossRef]
- Dimeglio, C.; Miedougé, M.; Loubes, J.M.; Mansuy, J.M.; Izopet, J. Side effect of a 6 p.m curfew for preventing the spread of SARS-CoV-2: A modeling study from Toulouse, France. J. Infect. 2021. [Google Scholar] [CrossRef]
- Graham, M.S.; Sudre, C.H.; May, A.; Antonelli, M.; Murray, B.; Varsavsky, T.; Klaser, K.; Canas, L.S.; Molteni, E.; Modat, M.; et al. Changes in symptomatology, reinfection, and transmissibility associated with the SARS-CoV-2 variant B.1.1.7: An ecological study. Lancet Public Health 2021. [Google Scholar] [CrossRef]
- Kostaki, E.G.; Tseti, I.; Tsiodras, S.; Pavlakis, G.N.; Sfikakis, P.P.; Paraskevis, D. Temporal Dominance of B.1.1.7 over B.1.354 SARS-CoV-2 Variant: A Hypothesis Based on Areas of Variant Co-Circulation. Life 2021, 11, 375. [Google Scholar]
- Dimeglio, C.; Miedougé, M.; Loubes, J.M.; Mansuy, J.M.; Izopet, J. Estimating the impact of public health strategies on the spread of SARS-CoV-2: Epidemiological modelling for Toulouse, France. Rev. Med. Virol. 2021, 13, e2224. [Google Scholar] [CrossRef]
- Dagan, N.; Barda, N.; Kepten, E.; Miron, O.; Perchik, S.; Katz, M.A.; Hernán, M.A.; Lipsitch, M.; Reis, B.; Balicer, R.D. BNT162b2 mRNA Covid-19 Vaccine in a Nationwide Mass Vaccination Setting. N. Engl. J. Med. 2021, 384, 1412–1423. [Google Scholar] [CrossRef]
- Amit, S.; Regev-Yochay, G.; Afek, A.; Kreiss, Y.; Leshem, E. Early rate reductions of SARS-CoV-2 infection and COVID-19 in BNT162b2 vaccine recipients. Lancet 2021, 397, 875–877. [Google Scholar] [CrossRef]
- Levine-Tiefenbrun, M.; Yelin, I.; Katz, R.; Herzel, E.; Golan, Z.; Schreiber, L.; Wolf, T.; Nadler, V.; Ben-Tov, A.; Kuint, J.; et al. Decreased SARS-CoV-2 viral load following vaccination. medRxiv 2021. [Google Scholar] [CrossRef]
- Petter, E.; Mor, O.; Zuckerman, N.; Oz-Levi, D.; Younger, A.; Aran, D.; Erlich, Y. Initial real world evidence for lower transmissibility of individuals who have been vaccinated by BNT162b2. medRxiv 2021. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Age Group | % In Toulouse Population. (Source INSEE) | A (Based on Toulouse Data) | |
|---|---|---|---|
| <15 years old | 14.8% | 10.2% | 20 |
| 15–60 years old | 68.2% | 61.58% | 20 |
| 60–74 years old | 10.4% | 10.68% | 20 |
| >74 years old | 5.5% | 17.54% | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimeglio, C.; Milhes, M.; Loubes, J.-M.; Ranger, N.; Mansuy, J.-M.; Trémeaux, P.; Jeanne, N.; Latour, J.; Nicot, F.; Donnadieu, C.; et al. Influence of SARS-CoV-2 Variant B.1.1.7, Vaccination, and Public Health Measures on the Spread of SARS-CoV-2. Viruses 2021, 13, 898. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050898
Dimeglio C, Milhes M, Loubes J-M, Ranger N, Mansuy J-M, Trémeaux P, Jeanne N, Latour J, Nicot F, Donnadieu C, et al. Influence of SARS-CoV-2 Variant B.1.1.7, Vaccination, and Public Health Measures on the Spread of SARS-CoV-2. Viruses. 2021; 13(5):898. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050898
Chicago/Turabian StyleDimeglio, Chloé, Marine Milhes, Jean-Michel Loubes, Noémie Ranger, Jean-Michel Mansuy, Pauline Trémeaux, Nicolas Jeanne, Justine Latour, Florence Nicot, Cécile Donnadieu, and et al. 2021. "Influence of SARS-CoV-2 Variant B.1.1.7, Vaccination, and Public Health Measures on the Spread of SARS-CoV-2" Viruses 13, no. 5: 898. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050898