
Comparison of the Virome of Quarantined Sugarcane Varieties and the Virome of Grasses Growing near the Quarantine Station

 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Quarantined Sugarcane Sampling
2.2. Wild Poaceae Sampling
2.3. Virion-Associated Nucleic Acid-Based Viral Metagenomics
2.4. Partial Genomic Characterization of a Novel Sugarcane Umbravirus
2.5. Phylogenetic Analyses
3. Results
3.1. Analysis of the Virome of Quarantined Sugarcane and of Wild Poaceae Growing Outside the Sugarcane Quarantine Glasshouse
3.2. Phylogenetic Relationships of Plant Viral Sequences Assigned to the Closteroviridae, Luteoviridae, and Potyviridae Families
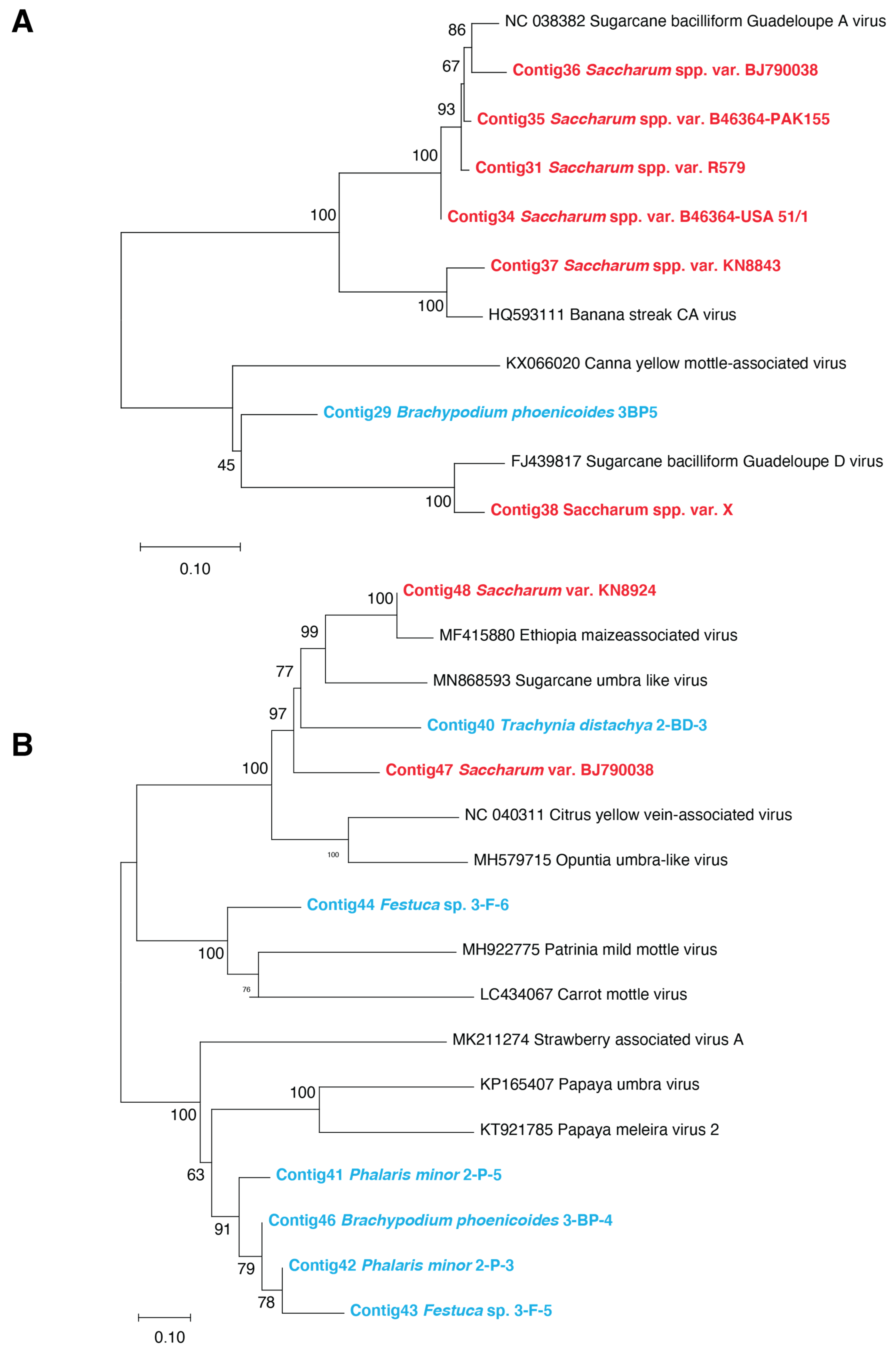
3.3. Phylogenetic Relationships of Plant Viral Sequences Assigned to the Mastrevirus, Badnavirus, and Umbravirus Genera
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin, R.R.; Constable, F.; Tzanetakis, I.E. Quarantine regulations and the impact of modern detection methods. Annu. Rev. Phytopathol. 2016, 54, 189–205. [Google Scholar] [CrossRef] [Green Version]
- Rott, P.; Bousquet, J.-F.; Muller, E.; Chatenet, M. La quarantaine de canne à sucre du CIRAD à Montpellier. Agric. Dév. 1997, 13, 22–28. [Google Scholar]
- Guinet-Brial, I.; Girard, J.-C.; Roumagnac, A.; Daugrois, J.-H.; Fernandez, E.; Rott, P. Visacane: An innovative quarantine tool for the exchange of pest and disease-free sugarcane germplasm. In International Society of Sugar Cane Technologists, Proceedings of the XXVIIIth Congress, São Paulo, Brazil, 24–27 June 2013; Sociedade dos Técnicos Açucareiros e Alcooleiros do Brasil (STAB): São Paulo, Brazil; The XXVIIIth ISSCT Organising Committee: São Paulo, Brazil, 2013; pp. 890–896. [Google Scholar]
- Fernandez, E.; Ferdinand, R.; Filloux, D.; Guinet, I.; Julian, C.; Rott, P.; Roumagnac, P.; Daugrois, J.-H. Improvements in virus detection at CIRAD’s sugarcane quarantine using both viral metagenomics and PCR-Based approaches. Sugar Tech 2020, 22, 498–503. [Google Scholar] [CrossRef]
- Candresse, T.; Filloux, D.; Muhire, B.; Julian, C.; Galzi, S.; Fort, G.; Bernardo, P.; Daugrois, J.-H.; Fernandez, E.; Martin, D.P.; et al. Appearances can be deceptive: Revealing a hidden viral infection with deep sequencing in a plant quarantine context. PLoS ONE 2014, 9, e102945. [Google Scholar] [CrossRef] [Green Version]
- Roumagnac, P.; Cirad, F.; Mollov, D.; Daugrois, J.-H.; Filloux, D.; Usda-Ars, U. Viral Metagenomics and Sugarcane Pathogens. In Understanding the Behaviour and Improving the Welfare of Dairy Cattle; Burleigh Dodds Science Publishing: Cambridge, UK, 2018; Volume 2, pp. 183–200. [Google Scholar]
- Filloux, D.; Fernandez, E.; Comstock, J.C.; Mollov, D.; Roumagnac, P.; Rott, P. Viral metagenomic-based screening of sugarcane from florida reveals occurrence of six sugarcane-infecting viruses and high prevalence of sugarcane yellow leaf virus. Plant Dis. 2018, 102, 2317–2323. [Google Scholar] [CrossRef] [Green Version]
- Rott, P.; Bailey, R.A.; Comstock, J.C.; Croft, B.J.; Saumtally, A.S. A Guide to Sugarcane Diseases; Cirad: Montpellier France, 2000. [Google Scholar]
- Tahir, M.N.; Bolus, S.; Grinstead, S.C.; McFarlane, S.A.; Mollov, D. A new virus of the family Tombusviridae infecting sugarcane. Arch. Virol. 2021, 166, 961–965. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.A.C.; Coutts, B.A. Spread of introduced viruses to new plants in natural ecosystems and the threat this poses to plant biodiversity. Mol. Plant Pathol. 2015, 16, 541–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redinbaugh, M.G.; Zambrano, J.L. Control of Virus Diseases in Maize. Adv. Clin. Chem. 2014, 90, 391–429. [Google Scholar] [CrossRef]
- Sun, S.-R.; Chen, J.-S.; Yang, J.; Huang, X.-C.; Huang, M.-T.; Gao, S.-J. First Report of Maize yellow mosaic virus Infecting Sugarcane in China. Plant Dis. 2019, 103, 2482. [Google Scholar] [CrossRef]
- Yahaya, A.; Dangora, D.B.; Alabi, O.J.; Zongoma, A.M.; Kumar, P.L. Detection and diversity of maize yellow mosaic virus infecting maize in Nigeria. VirusDisease 2019, 30, 538–544. [Google Scholar] [CrossRef]
- Grisham, M.P. Strains of sorghum mosaic virus causing sugarcane mosaic in Louisiana. Plant Dis. 1994, 78, 729–732. [Google Scholar] [CrossRef]
- Chen, J.; Chen, J. Sugarcane mosaic disease in Zhejiang Province was caused by sorghum mosaic virus and sugarcane mosaic virus. Bing Du Xue Bao 2002, 18, 362–366. [Google Scholar]
- Chalupniková, J.; Kundu, J.K.; Singh, K.; Bartaková, P.; Beoni, E. Wheat streak mosaic virus: Incidence in field crops, potential reservoir within grass species and uptake in winter wheat cultivars. J. Integr. Agric. 2017, 16, 523–531. [Google Scholar] [CrossRef]
- Fargette, D.; Konaté, G.; Fauquet, C.; Muller, E.; Peterschmitt, M.; Thresh, J.M. Molecular Ecology and Emergence of Tropical Plant Viruses. Annu. Rev. Phytopathol. 2006, 44, 235–260. [Google Scholar] [CrossRef] [PubMed]
- Claverie, S.; Ouattara, A.; Hoareau, M.; Filloux, D.; Varsani, A.; Roumagnac, P.; Martin, D.P.; Lett, J.-M.; Lefeuvre, P. Exploring the diversity of Poaceae-infecting mastreviruses on Reunion Island using a viral metagenomics-based approach. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ellis, M.H.; Rebetzke, G.J.; Moore, C.S.; Hyles, J.E.; Kelman, W.M. Detection of Wheat streak mosaic virus in four pasture grass species in Australia. Plant Pathol. 2004, 53, 239. [Google Scholar] [CrossRef]
- Malmstrom, C.M.; McCullough, A.J.; Johnson, H.A.; Newton, L.A.; Borer, E.T. Invasive annual grasses indirectly increase virus incidence in California native perennial bunchgrasses. Oecologia 2005, 145, 153–164. [Google Scholar] [CrossRef]
- Kumar, A.; Murthy, S.; Kapoor, A. Evolution of selective-sequencing approaches for virus discovery and virome analysis. Virus Res. 2017, 239, 172–179. [Google Scholar] [CrossRef]
- Roossinck, M.J.; Martin, D.P.; Roumagnac, P. Plant Virus metagenomics: Advances in virus discovery. Phytopathology 2015, 105, 716–727. [Google Scholar] [CrossRef] [Green Version]
- Boukari, W.; Alcalá-Briseño, R.I.; Kraberger, S.; Fernandez, E.; Filloux, D.; Daugrois, J.-H.; Comstock, J.C.; Lett, J.-M.; Martin, D.P.; Varsani, A.; et al. Occurrence of a novel mastrevirus in sugarcane germplasm collections in Florida, Guadeloupe and Réunion. Virol. J. 2017, 14, 146. [Google Scholar] [CrossRef]
- François, S.; Bernardo, P.; Filloux, D.; Roumagnac, P.; Yaverkovski, N.; Froissart, R.; Ogliastro, M. A novel itera-like densovirus isolated by viral metagenomics from the Sea Barley Hordeum marinum. Genome Announc. 2014, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraberger, S.; Farkas, K.; Bernardo, P.; Booker, C.; Argüello-Astorga, G.R.; Mesléard, F.; Martin, D.P.; Roumagnac, P.; Varsani, A. Identification of novel Bromus- and Trifolium-associated circular DNA viruses. Arch. Virol. 2015, 160, 1303–1311. [Google Scholar] [CrossRef]
- Bernardo, P.; Charles-Dominique, T.; Barakat, M.; Ortet, P.; Fernandez, E.; Filloux, D.; Hartnady, P.A.; Rebelo, T.; Cousins, S.R.; Mesleard, F.; et al. Geometagenomics illuminates the impact of agriculture on the distribution and prevalence of plant viruses at the ecosystem scale. ISME J. 2018, 12, 173–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- François, S.; Filloux, D.; Fernandez, E.; Ogliastro, M.; Roumagnac, P. Viral metagenomics approaches for high-resolution screening of multiplexed arthropod and plant viral communities. Breast Cancer 2018, 1746, 77–95. [Google Scholar] [CrossRef]
- van der Valk, T.; Vezzi, F.; Ormestad, M.; Dalén, L.; Guschanski, K. Estimating the rate of index hopping on the Illumina HiSeq X platform. bioRxiv 2018, 179028. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Manber, U. A fast approximate pattern-matching tool. In Proceedings of the Usenix Winter 1992 Technical Conference, San Francisco, CA, USA, 20–24 January 1992; pp. 153–162. [Google Scholar]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet 2011, 17, 10. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Wylie, S.J.; Adams, M.; Chalam, C.; Kreuze, J.; López-Moya, J.J.; Ohshima, K.; Praveen, S.; Rabenstein, F.; Stenger, D.; Wang, A.; et al. ICTV Virus Taxonomy Profile: Potyviridae. J. Gen. Virol. 2017, 98, 352–354. [Google Scholar] [CrossRef] [PubMed]
- Muhire, B.; Martin, D.P.; Brown, J.K.; Navas-Castillo, J.; Moriones, E.; Zerbini, F.M.; Rivera-Bustamante, R.; Malathi, V.G.; Briddon, R.W.; Varsani, A. A genome-wide pairwise-identity-based proposal for the classification of viruses in the genus Mastrevirus (family Geminiviridae). Arch. Virol. 2013, 158, 1411–1424. [Google Scholar] [CrossRef] [Green Version]
- Muller, E.; Dupuy, V.; Blondin, L.; Bauffe, F.; Daugrois, J.-H.; Nathalie, L.; Iskra-Caruana, M.-L. High molecular variability of sugarcane bacilliform viruses in Guadeloupe implying the existence of at least three new species. Virus Res. 2011, 160, 414–419. [Google Scholar] [CrossRef]
- Roossinck, M.J.; Saha, P.; Wiley, G.B.; Quan, J.; White, J.D.; Lai, H.; Chavarría, F.; Shen, G.; Roe, B.A. Ecogenomics: Using massively parallel pyrosequencing to understand virus ecology. Mol. Ecol. 2010, 19, 81–88. [Google Scholar] [CrossRef]
- Sastry, K.S.; Mandal, B.; Hammond, J.; Scott, S.W.; Briddon, R.W. Encyclopedia of Plant Viruses and Viroids; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Diaz-Lara, A.; Mosier, N.J.; Keller, K.E.; Martin, R.R. A variant of Rubus yellow net virus with altered genomic organization. Virus Genes 2014, 50, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Basso, M.F.; Fajardo, T.V.M.; Saldarelli, P. Grapevine virus diseases:economic impact and current advances in viral prospection and management. Rev. Bras. Frutic. 2017, 39, e-411. [Google Scholar] [CrossRef] [Green Version]
- Rumbou, A.; Candresse, T.; Marais, A.; Theil, S.; Langer, J.; Jalkanen, R.; Büttner, C. A novel badnavirus discovered from Betula sp. affected by birch leaf-roll disease. PLoS ONE 2018, 13, e0193888. [Google Scholar] [CrossRef]
- Minafra, A.; Chiumenti, M.; Elbeaino, T.; Digiaro, M.; Bottalico, G.; Pantaleo, V.; Martelli, G. Occurrence of fig Badnavirus 1 in fig trees from different countries and in symptomless seedlings. J. Plant Pathol. 2012, 94, 4–105. [Google Scholar]
- Wu, X.; Alabi, O.J.; Damaj, M.B.; Sun, S.; Mirkov, T.E.; Fu, H.; Chen, R.; Gao, S. Prevalence and RT/RNase H Genealogy of Sugarcane Bacilliform Virus Isolates from China. J. Phytopathol. 2016, 164, 595–607. [Google Scholar] [CrossRef]
- Agindotan, B.O.; Domier, L.L.; Bradley, C.A. Detection and characterization of the first North American mastrevirus in switchgrass. Arch. Virol. 2015, 160, 1313–1317. [Google Scholar] [CrossRef]
- Hadfield, J.; Martin, D.P.; Stainton, D.; Kraberger, S.; Owor, B.E.; Shepherd, D.N.; Lakay, F.; Markham, P.G.; Greber, R.S.; Briddon, R.W.; et al. Bromus catharticus striate mosaic virus: A new mastrevirus infecting Bromus catharticus from Australia. Arch. Virol. 2010, 156, 335–341. [Google Scholar] [CrossRef]
- Oluwafemi, S.; Kraberger, S.; Shepherd, D.N.; Martin, D.P.; Varsani, A. A high degree of African streak virus diversity within Nigerian maize fields includes a new mastrevirus from Axonopus compressus. Arch. Virol. 2014, 159, 2765–2770. [Google Scholar] [CrossRef] [PubMed]
- Manurung, B.; Witsack, W.; Mehner, S.; Grüntzig, M.; Fuchs, E. The epidemiology of Wheat dwarf virus in relation to occurrence of the leafhopper Psammotettix alienus in Middle-Germany. Virus Res. 2004, 100, 109–113. [Google Scholar] [CrossRef]
- Achon, M.A.; Serrano, L.; Ratti, C.; Rubies-Autonell, C. First Detection of Wheat dwarf virus in Barley in Spain Associated with an Outbreak of Barley Yellow Dwarf. Plant Dis. 2006, 90, 970. [Google Scholar] [CrossRef] [PubMed]
- Lemmetty, A.; Huusela-Veistola, E. First report of wheat dwarf virus in winter wheat in finland. Plant Dis. 2005, 89, 912. [Google Scholar] [CrossRef]
- Schubert, J.; Habekuß, A.; Kazmaier, K.; Jeske, H. Surveying cereal-infecting geminiviruses in Germany—Diagnostics and direct sequencing using rolling circle amplification. Virus Res. 2007, 127, 61–70. [Google Scholar] [CrossRef]
- Adams, I.P.; Braidwood, L.A.; Stomeo, F.; Phiri, N.; Uwumukiza, B.; Feyissa, B.; Mahuku, G.; Wangi, A.; Smith, J.; Mumford, R.; et al. Characterising maize viruses associated with maize lethal necrosis symptoms in sub Saharan Africa. bioRxiv 2017, 161489. [Google Scholar]
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Dietzgen, R.G.; Mann, K.S.; Johnson, K.N. Plant virus–insect vector interactions: Current and potential future research directions. Viruses 2016, 8, 303. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Quarantined Sugarcane | Wild Poaceae from Flat Stone Land | Wild Poaceae from Sloped Land | Wild Poaceae from Grassland | |||||
|---|---|---|---|---|---|---|---|---|
| Variety | Number of Reads | Species | Number of Reads | Species | Number of Reads | Species | Number of Reads | |
| KN88260 | 169,914 | Aegilops triuncialis (3) a | 1,228,780 | Aegilops triuncialis (3) | 455,952 | Avena sterilis (2) | 376,016 | |
| R579 | 337,700 | Aegilops geniculata (3) | 1,051,536 | Aegilops geniculata (3) | 425,570 | Bothriochloa barbinodis (4) | 1,190,788 | |
| KN88104 | 364,446 | Anisantha madritensis (6) | 1,661,880 | Avena sterilis (2) | 249,804 | Brachypodium phoenicoides (5) | 885,284 | |
| Q112 | 189,526 | Avena sterilis (2) | 800,428 | Bothriochloa barbinodis (3) | 981,718 | Bromopsis erecta (3) | 1,028,474 | |
| B46364-USA51/1 | 301,906 | Bothriochloa ischaemum (6) | 931,920 | Trachynia distachya (2) | 374,270 | Cynodon dactylon (2) | 447,196 | |
| KN88147 | 483,278 | Trachynia distachya (3) | 1,088,638 | Brachypodium phoenicoides (2) | 859,170 | Festuca sp. (5) | 728,278 | |
| BJ79038 | 244,194 | Bromopsis erecta (2) | 1,102,196 | Bromopsis erecta (3) | 152,906 | Gastridium ventricosum (6) | 1,065,580 | |
| KN8924 | 625,654 | Bromus lanceolatus (3) | 516,490 | Catapodium rigidum (2) | 681,122 | Helictochloa bromoides (1) | 150,384 | |
| KN8843 | 391,108 | Catapodium rigidum (2) | 922,976 | Cynodon dactylon (2) | 539,866 | |||
| GT9 | 596,222 | Cynodon dactylon (2) | 52,6150 | Phalaris minor (6) | 1,161,018 | |||
| B46364-PAK155 | 386,752 | Elytrigia intermedia (6) | 1,270,630 | |||||
| X | 269,742 | Hainardia cylindrical (6) | 998,856 | |||||
| USDA | 598,072 | Holcus lanatus (2) | 386,482 | |||||
| LF653661 | 189,262 | Hordeum murinum subsp. Leporinum (6) | 806,796 | |||||
| NA021668 | 236,036 | Lolium rigidum (6) | 1,353,330 | |||||
| SP701284 | 267,242 | Melica ciliata (6) | 520,800 | |||||
| CR9821 | 186,898 | Piptatherum miliaceum (1) | 284,228 | |||||
| Q140 | 218,214 | Rostraria cristata (3) | 602,560 | |||||
| FG087484 | 128,726 | Vulpia ciliata (6) | 570,698 | |||||
| FR95433 | 125,826 | |||||||
| Total (samples) reads | ||||||||
| (20) | 6,310,178 | (74) | 12,561,594 | (28) | 4,388,287 | (28) | 5,872,000 | |
| Putative Virus | Virus Host * | Wild Poaceae | Quarantined Sugarcane | ||
|---|---|---|---|---|---|
| Number of Samples | Number of Reads | Number of Samples | Number of Reads | ||
| Geminiviridae | Plant | 2 | 48,512 | 8 | 833,570 |
| Potyviridae | Plant | 2 | 15,029 | 3 | 414,737 |
| Tombusviridae | Plant | 13 | 3578 | 2 | 1339 |
| Luteoviridae | Plant | 2 | 270 | 2 | 83,338 |
| Closteroviridae | Plant | 1 | 368 | 7 | 10,260 |
| Alphasatellitidae | Plant | 3 | 140,222 | ||
| Amalgaviridae | Plant | 4 | 657 | ||
| Bromoviridae | Plant | 1 | 25 | ||
| Aspiviridae | Plant | 1 | 19 | ||
| Sobemovirus | Plant | 1 | 20 | ||
| Trifolium-ass. DNA virus | Plant | 3 | 587 | ||
| Caulimoviridae | Plant/insect | 2 | 44 | 9 | 37,192 |
| Partitiviridae | Plant/fungus | 89 | 89,391 | 5 | 210 |
| Endornaviridae | Plant/fungus/oomycete | 2 | 66 | ||
| Rhabdoviridae | Plant/vertebrate/invertebrate | 4 | 493 | ||
| Reoviridae | Plant/fungus/vertebrate/invertebrate | 1 | 413 | ||
| Chrysoviridae | Fungus | 29 | 4197 | ||
| Narnaviridae | Fungus | 1 | 15 | ||
| Hypoviridae | Fungus | 6 | 560 | ||
| Gammaflexiviridae | Fungus | 2 | 477 | ||
| Metaviridae | Fungus | 2 | 125 | ||
| Botybirnavirus | Fungus | 21 | 2554 | ||
| Tymovirales (unclassified) | Fungus | 2 | 75 | ||
| Unclassified fungal viruses | Fungus | 31 | 35,671 | ||
| Totiviridae | Fungus/protozoan | 40 | 60,090 | ||
| Genomoviridae | Fungus/human/mammal/bird | 30 | 6678 | 1 | 15 |
| Microviridae | Bacteria/spiroplasma | 26 | 35,410 | ||
| Myoviridae | Bacteria/archaea | 5 | 1419 | 1 | 13 |
| Podoviridae | Bacteria/archaea | 5 | 6673 | ||
| Siphoviridae | Bacteria/archaea | 5 | 152,598 | ||
| Unclassified bacterial viruses | Bacteria | 3 | 108 | ||
| Iflaviridae | Insect | 1 | 220 | ||
| Bidnaviridae | Insect | 1 | 32 | ||
| Parvoviridae | Insect/vertebrate | 10 | 3271 | ||
| Baculoviridae | Insect/decapod | 1 | 19 | 1 | 18 |
| Peribunyaviridae | Insect/rodent | 1 | 16 | ||
| Unclassified arthropod viruses | Arthropod | 18 | 20,812 | ||
| Retroviridae | Vertebrate | 2 | 70 | ||
| Circoviridae | Bird/mammal | 26 | 8208 | ||
| Iridoviridae | Amphibia/fish/invertebrate | 1 | 23 | ||
| Phycodnaviridae | Alga | 15 | 450 | 1 | 19 |
| Mimiviridae | Amoebae | 10 | 277 | ||
| Nimaviridae | Crustacean | 1 | 15 | ||
| Arenaviridae | Rodent | 1 | 19 | ||
| unclassified | Vertebrate/invertebrate/crustacean | 11 | 3915 | ||
| unclassified | environmental | 8 | 96,793 | ||
| unclassified | unknown | 6 | 33,824 | ||
| Total reads | 634,566 | 1,520,933 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daugrois, J.H.; Filloux, D.; Julian, C.; Claude, L.; Ferdinand, R.; Fernandez, E.; Fontes, H.; Rott, P.C.; Roumagnac, P. Comparison of the Virome of Quarantined Sugarcane Varieties and the Virome of Grasses Growing near the Quarantine Station. Viruses 2021, 13, 922. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050922
Daugrois JH, Filloux D, Julian C, Claude L, Ferdinand R, Fernandez E, Fontes H, Rott PC, Roumagnac P. Comparison of the Virome of Quarantined Sugarcane Varieties and the Virome of Grasses Growing near the Quarantine Station. Viruses. 2021; 13(5):922. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050922
Chicago/Turabian StyleDaugrois, Jean H., Denis Filloux, Charlotte Julian, Lisa Claude, Romain Ferdinand, Emmanuel Fernandez, Hugo Fontes, Philippe C. Rott, and Philippe Roumagnac. 2021. "Comparison of the Virome of Quarantined Sugarcane Varieties and the Virome of Grasses Growing near the Quarantine Station" Viruses 13, no. 5: 922. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050922