
Phage Biocontrol of Pseudomonas aeruginosa in Water
by
 , , , ,
, , , ,
Ari Kauppinen
1,*,† ,
,
Sallamaari Siponen
1,2 ,
,
Tarja Pitkänen
1,3,
Karin Holmfeldt
4,
Anna Pursiainen
1,
Eila Torvinen
2 and
Ilkka T. Miettinen
1 1
Expert Microbiology Unit, Department of Health Security, Finnish Institute for Health and Welfare, FI-70701 Kuopio, Finland
2
Department of Environmental and Biological Sciences, University of Eastern Finland, FI-70211 Kuopio, Finland
3
Department of Food Hygiene and Environmental Health, Faculty of Veterinary Medicine, University of Helsinki, FI-00014 Helsinki, Finland
4
Centre for Ecology and Evolution in Microbial Model Systems (EEMiS), Department of Biology and Environmental Sciences, Linnaeus University, SE-39231 Kalmar, Sweden
*
Author to whom correspondence should be addressed.
†
Current address: Virology Unit, Laboratory and Research Division, Finnish Food Authority, FI-00027 Helsinki, Finland.
Viruses 2021, 13(5), 928; https://0-doi-org.brum.beds.ac.uk/10.3390/v13050928
Submission received: 10 April 2021
/
Revised: 7 May 2021
/
Accepted: 11 May 2021
/
Published: 17 May 2021
(This article belongs to the Special Issue Viruses in the Environment)
Abstract
:Bacteriophage control of harmful or pathogenic bacteria has aroused growing interest, largely due to the rise of antibiotic resistance. The objective of this study was to test phages as potential agents for the biocontrol of an opportunistic pathogen Pseudomonas aeruginosa in water. Two P. aeruginosa bacteriophages (vB_PaeM_V523 and vB_PaeM_V524) were isolated from wastewater and characterized physically and functionally. Genomic and morphological characterization showed that both were myoviruses within the Pbunavirus genus. Both had a similar latent period (50–55 min) and burst size (124–134 PFU/infected cell), whereas there was variation in the host range. In addition to these environmental phages, a commercial Pseudomonas phage, JG003 (DSM 19870), was also used in the biocontrol experiments. The biocontrol potential of the three phages in water was tested separately and together as a cocktail against two P. aeruginosa strains; PAO1 and the environmental strain 17V1507. With PAO1, all phages initially reduced the numbers of the bacterial host, with phage V523 being the most efficient (>2.4 log10 reduction). For the environmental P. aeruginosa strain (17V1507), only the phage JG003 caused a reduction (1.2 log10) compared to the control. The cocktail of three phages showed a slightly higher decrease in the level of the hosts compared to the use of individual phages. Although no synergistic effect was observed in the host reduction with the use of the phage cocktail, the cocktail-treated hosts did not appear to acquire resistance as rapidly as hosts treated with a single phage. The results of this study provide a significant step in the development of bacteriophage preparations for the control of pathogens and harmful microbes in water environments.
1. Introduction
The control of harmful bacteria is an increasingly important and topical public health issue as antibiotic-resistant and disinfection-tolerant pathogenic strains become more common. Bacteriophages, which are viruses that exclusively infect bacteria, play an important role in biological processes by controlling the natural balance in an ecosystem, and hold great potential for targeting harmful bacteria. For biocontrol applications, promising candidates are broad-host-range lytic phages, which destroy the infected cell after immediate replication of the virion. One of the benefits of using bacteriophages over antibiotics or disinfectants is their specificity, i.e., they can target just the pathogenic bacteria of interest, rather than all bacteria. Currently, bacteriophages have mainly been tested in clinical settings [1,2] and environmental applications are still rare. However, the interest in phages has recently increased, e.g., in agriculture [3], aquaculture [3,4] and the food industry [5,6,7]. Biocontrol using bacteriophages may also have potential in several applications in water environments, such as for the mitigation of pathogens, including antibiotic-resistant strains, as well as harmful microbes responsible for causing problematic corrosion and biofouling [8,9,10,11,12,13].
The prevalence of antibiotic resistance (AR) has increased and become more versatile in past years, and currently AR is one of the most significant health threats to modern society [14,15]. The environment may play an important role in the spread of AR and water can act as a vector, enabling the spread of AR properties [16,17]. Currently, antibiotic-resistant bacterial strains have been described for several opportunistic pathogens, including multidrug-resistant P. aeruginosa. This Gram-negative bacterium is an opportunistic pathogen that is capable of causing acute and chronic infections of wounds, eyes, ears and respiratory and urinary tracts, particularly among immunocompromised people [18]. Metabolic versatility, the ability to form biofilms and drug/chemical resistance are characteristics enabling the success of P. aeruginosa in clinical and environmental settings, including in water [19,20,21].
The role of P. aeruginosa in outbreaks related to contaminated tap or bottled water has been well described [22,23,24,25,26]. Moreover, P. aeruginosa has been frequently isolated in the water of swimming pools and spas [27,28], and several outbreaks have been described [29,30]. P. aeruginosa benefits from the warm and moist conditions present in swimming pools and spas, and it can tolerate chlorination (<1 mg/L) to some extent [30]. Although chlorination is a widely used disinfection method in water environments, the excessive chlorination required to control P. aeruginosa has several drawbacks, such as the formation of harmful disinfection by-products. Furthermore, chlorination is relatively ineffective against microbes protected by biofilms, and it may compromise system performance if used, e.g., in biological water treatment processes [12]. Moreover, previous studies have indicated that chlorine exposure may promote the development of AR in bacteria [31,32]. Therefore, new control options are needed and biocontrol using bacteriophages is one of the promising alternatives.
In this study, three P. aeruginosa phages were tested against two P. aeruginosa strains in order to determine their potential as biocontrol agents in water. The results of this study provide insights into potential future applications in water treatment and reuse.
2. Materials and Methods
2.1. P. aeruginosa Strains and Growth Conditions
A P. aeruginosa PAO1 (DSM 19880) strain, purchased from the Leibniz Institute DSMZ (German Collection of Microorganisms and Cell Cultures GmbH, Braunschweig, Germany), and the environmental strain P. aeruginosa 17V1507, isolated from swimming pool water, were used for this study. P. aeruginosa 17V1507 was extracted from a swimming pool water sample according to EN ISO 16266:2008 and confirmed using an API 20 NE test (Biomerieux, Marcy-l’Étoile, France). P. aeruginosa was propagated in a tryptic soy broth (TSB) and agar (TSA) for 21 ± 3 h, at 36 °C ± 2 °C.
2.2. Phages and Growth Conditions
A Pseudomonas phage JG003 isolate (DSM 19870), purchased from the Leibniz Institute DSMZ (German Collection of Microorganisms and Cell Cultures GmbH, Braunschweig, Germany), was used for this study. In addition, two environmental strains (Pseudomonas phage vB_PaeM_V523 and Pseudomonas phage vB_PaeM_V524) isolated from wastewater using P. aeruginosa PAO1 (DSM 19880) as a host (see isolation procedures below) were exploited. P. aeruginosa phages were enumerated using a double agar layer (DAL) method [33] from serially diluted samples using PAO1 (DSM 19880) as a host. DAL-plates (1.5% TSA bottom agar with 0.7% TSA top agar) were incubated for 21 ± 3 h at 36 °C ± 2 °C.
2.3. Phage Isolation and Purification
Two environmental P. aeruginosa phage strains (Pseudomonas phage vB_PaeM_V523 and Pseudomonas phage vB_PaeM_V524) were isolated from the influent of a municipal wastewater treatment plant (Kuopio, Finland). A 100 mL wastewater sample was centrifuged at 9000× g for 10 min at 4 °C and the supernatant was filtered through 0.45-µm (Acrodisc, Pall Corporation, New York, NY, USA) and 0.22-µm (Minisart, Sartorius, Göttingen, Germany) syringe filters. Plaques were produced from the filtered sample using the DAL method. Three rounds of plaque purification were conducted for each plaque with a protocol slightly modified from that of Azeredo et al. [34]. Briefly, the plaques were isolated by picking them with a pipette tip and placing them in a saline magnesium (SM) buffer and then centrifuging them at 13,000× g for 5 min. The supernatant was analyzed using the DAL method for the production of the next round of purification. Finally, plaques were eluted from the plates in the SM buffer with gentle agitation (90 rpm) for 5 h at 4 °C, and were then centrifuged at 9000× g for 10 min at 4 °C and filtered through a 0.22-µm (Minisart, Sartorius, Göttingen, Germany) syringe filter. The filtered JG003 isolate (DSM 19870) phage stock was produced using the DAL method and elution as described above. The phage stocks were stored at 4 °C until use.
2.4. Transmission Electron Microscopy (TEM)
Transmission electron microscopy (TEM) was used for the determination of the phage morphology. A filtered high-titer (about 1011 PFU/mL) phage stock solution was deposited on a Formvar- and carbon-coated grid (Agar Scientific, Essex, UK) and incubated for 30 s. The excess liquid was removed with a filter paper before the grid was negative stained with phosphor tungstic acid (5%) for 30 s. Phages were examined using a JEM-2100F TEM (JEOL, Tokyo, Japan) in SIB Labs, University of Eastern Finland, Finland.
2.5. Phage Host Range
The host range was determined for the three phages (V523, V524 and JG003) using a spot test on a double agar overlay with undiluted phage stock and up to 108 serial diluted phage suspensions (with titers about 103 to 1011 PFU/mL). In total, 36 P. aeruginosa strains, which included ten strains isolated from clinical samples, 23 strains from environmental samples (three strains from wastewater samples, eight from swimming pool samples and twelve from drinking water tanks) and three reference strains (DSM 19880, DSM 50071 and ATCC 27853) were tested. In addition, non-P. aeruginosa bacteria were tested using reference strains of Pseudomonas fluorescens (ATCC 49642), Enterobacter aerogenes (DSM 30053), Enterobacter cloacae (DSM 30054) and three Escherichia coli strains (DSM 1103, DSM 30083 and DSM 1576). Aliquots of 10 μL of phage stock or dilution were spotted with a pipette onto the surface of an agar overlay. The plates were incubated for 21 ± 3 h at 36 °C ± 2 °C. The susceptibility of each host to the different phages was evaluated for the appearance of lytic clearing zones, which were classified as clear, turbid or not present.
2.6. One-Step Growth Curve
One-step growth curve analyses were performed as previously described [35] to determine the latent periods and burst sizes of the phages. Here, a 20-mL log-phase PAO1 bacterial culture (optical density at 520 nm of 0.24 = 108 CFU/mL) was combined with a phage suspension (105 PFU/mL) to obtain a multiplicity of infection (MOI) of 0.001. The phages were allowed to adsorb for 10 min at 36 °C ± 2 °C, after which the mixture was diluted to 10−3. The diluted mixture was incubated at 36 °C ± 2 °C with shaking at 100 rpm. Triplicate 1 mL samples were taken over one hour at 5 min intervals and titrated using the DAL method. The burst size was calculated as the difference between the final and the initial phage titer divided by the initial phage titer.
2.7. Phage Genome Sequencing and Assembly
The phage genome was extracted from a 200 μL volume using the High Pure Viral Nucleic Acid Kit according to the manufacturer’s instructions (Roche Molecular Biochemicals Ltd., Mannheim, Germany). The extracted nucleic acids were stored at ≤−75 °C.
The purified DNA was sequenced at SciLife/NGI (Solna, Sweden). A Nextera XT Kit (Illumina Inc., San Diego, CA, USA) was used for the library preparation. Paired-end (2 × 125 bp) sequencing was performed on a HiSeq 2500 instrument (Illumina Inc., San Diego, CA, USA). Raw reads were trimmed to remove adapters and poor quality reads (Trimmomatic version 0.30, settings: -PE –threads 2 -phred33 ILLUMINACLIP:nextera_linkers.txt:2:30:10 LEADING:3 TRAILING:3 SLIDINGWINDOW:4:15 MINLEN:30) [36] and quality was evaluated with FastQC [37]. Trimmed reads were then assembled into contigs using Spades (version 3.6.0, settings: --careful –t 8 --pe1-1 --pe1-2 -o) with a k-mer length of 99 [38]. A first comparison of the genomes to known, previously isolated viruses was conducted with an online basic local alignment search tool, nucleotide, (blastn) against an NCBI nucleotide collection (nr) using the default settings. Given the high similarity of the genomes with phages within the genus Pbunavirus, the type-species within this genus, as well as the two phages that V523 and V524 shared the highest similarity with, were downloaded from NCBI for further comparisons (Supplementary Table S1).
An investigation of the taxonomic affiliation of the viruses was conducted using the VIRIDIC [39] and VICTOR [40] online tools with default settings, in which V523 and V524 were compared to the downloaded Pbunavirus phages.
Open reading frames (ORFs) were called using GenMark.hmm with Heuristic models [41]. The ORFs were validated through comparison with their closest relatives available in NCBI. Predicted proteins were annotated for function through comparison with the NCBI viral database (ViralDB) created from all viral genomes in the NCBI RefSeq database (July 2019) using BLASTp (v 2.7.1, e-value cut-off 0.001). In addition, hmm searches were conducted against the PFAM database (Nov 2020) using the online search tool. The functional annotation, as well as the similarity of V523 and V524 to a closely related Pbunavirus phage, were displayed using Easyfig [42].
2.8. Phage Biocontrol of P. aeruginosa in Water
Phage biocontrol tests were carried out in 500-mL Erlenmeyer flasks coated with aluminum foil to obtain dark conditions. The test water was the drinking water from the municipality of Siilinjärvi, Finland, which originates from a groundwater source and is disinfected with UV light, using no chlorine prior to distribution. The drinking water was autoclaved prior to tests. The general physical-chemical properties of the drinking water used in this study are presented in Table 1. Phage biocontrol tests were carried out with the P. aeruginosa PAO1 strain and the environmental P. aeruginosa strain 17V1507. A log-phase bacterial culture was washed twice prior to inoculation, first with a phosphate buffer and then with test water, pelleting the cells by means of centrifugation at 8000× g for 2 min. Triplicate 200-mL water samples were inoculated with a 200-µL log-phase P. aeruginosa culture (105 CFU/mL) and were kept in agitation (100–125 rpm) at 36 °C ± 2 °C. The phages were spiked in samples after a 24 h adaptation of the host to the test water with the aim of achieving about 108 PFU/mL (MOI of 1000). Triplicate 5 mL samples were taken after 0, 6, 24 (before spike), 24 (after spike), 25, 27, 48, 54 and 72 h from the beginning of the experimental work. Samples were kept in constant agitation (100–125 rpm) and shaken vigorously by hand for 15 s before sampling. Negative controls containing the host without the phage spike were performed for both P. aeruginosa strains. Phage controls without the host were also performed for each phage. The samples were immediately subjected to CFU determination of the host bacteria and PFU of phages.
2.9. Phage Resistance
The emergence of phage resistance was assessed in the biocontrol study conducted in water. Variation in colony morphologies and increased growth of hosts as the experiment progressed were considered to be due to resistance. To assess the persistence of phage resistance, three consecutive passages on TSA plates were performed for the PAO1 host treated with phage V524. A typical spherical colony was picked from the plate and cultivated on TSA agar plates three times. The resistance of this purified colony was tested against three phages (V523, V524 and JG003) using a spot test on a double agar overlay with undiluted phage stock.
2.10. Data Analysis
The statistical analyses were conducted using SPSS, version 25 software for Windows (SPSS Inc., Chicago, IL, USA). Log10 reductions were calculated as the average of three replicates. Differences between the average log10 reductions of hosts achieved with the individual phages and cocktail were tested using the related-samples Wilcoxon signed-rank test. The method detection limit values were used for the calculations when a below limit of detection result was obtained. Differences were considered significant if the p-value was <0.05.
3. Results
3.1. Phage Isolation and Physical Characterization
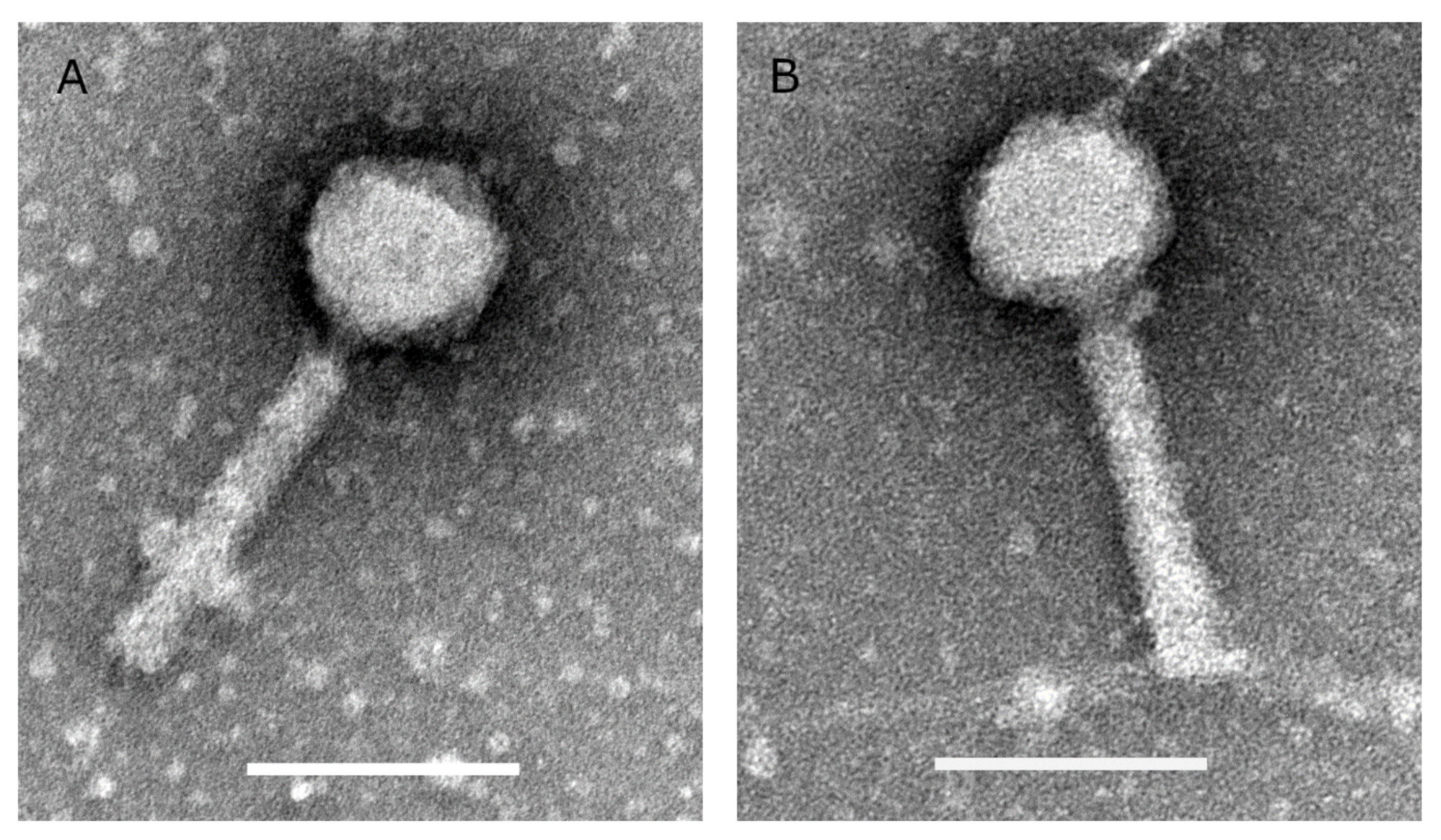
Two P. aeruginosa phages (V523 and V524) were isolated from wastewater and produced in high titers (1 × 1011–8 × 1011 PFU/mL). Both phages have isometric heads with a diameter of about 75 nm and contractile tails of about 140 nm (Figure 1).
3.2. Phage Host Range
All three phages showed a broad host range against tested P. aeruginosa strains (Table 2). Phage JG003 exhibited the broadest activity, lysing 97% of the strains, compared to 67% for phage V523 and 83% for phage V524. The proportions of complete clear lysis zones in spot tests were as follows: 33% for JG003, 14% for V523 and 42% for V524. To verify a positive result, spot tests in dilution series were performed and plaques were obtained in all cases. No cross-reactivity to non-P. aeruginosa strains was detected. Spot tests confirmed that all three phages were lytic against PAO1, and for the environmental strain 17V1507 phage V524 and JG003 were lytic.
3.3. One-Step Growth Curve
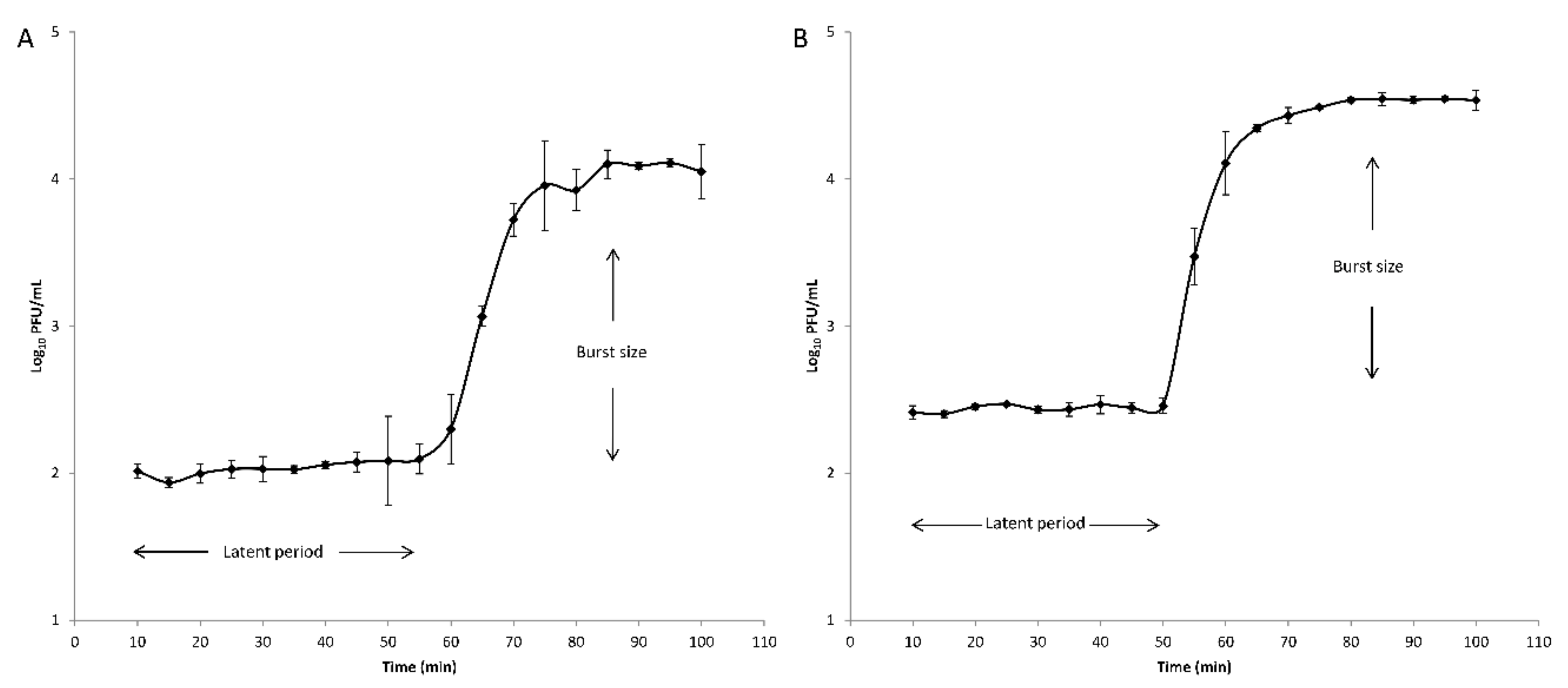
Latent periods and burst sizes were determined for phages V523 and V524 through one-step growth curve analyses using PAO1 as a host (Figure 2). The latent periods were 55 min and 50 min and the burst sizes were 124 PFU/infected cell and 134 PFU/infected cell for phages V523 and V524, respectively.
3.4. Phage Genome Characterization
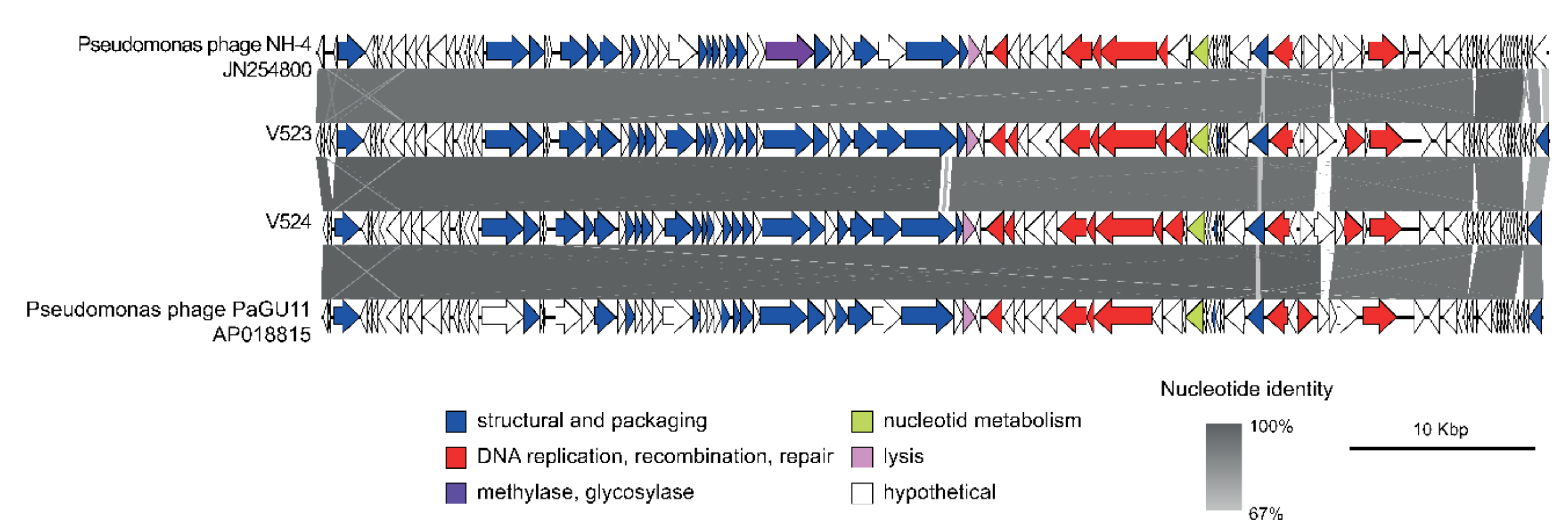
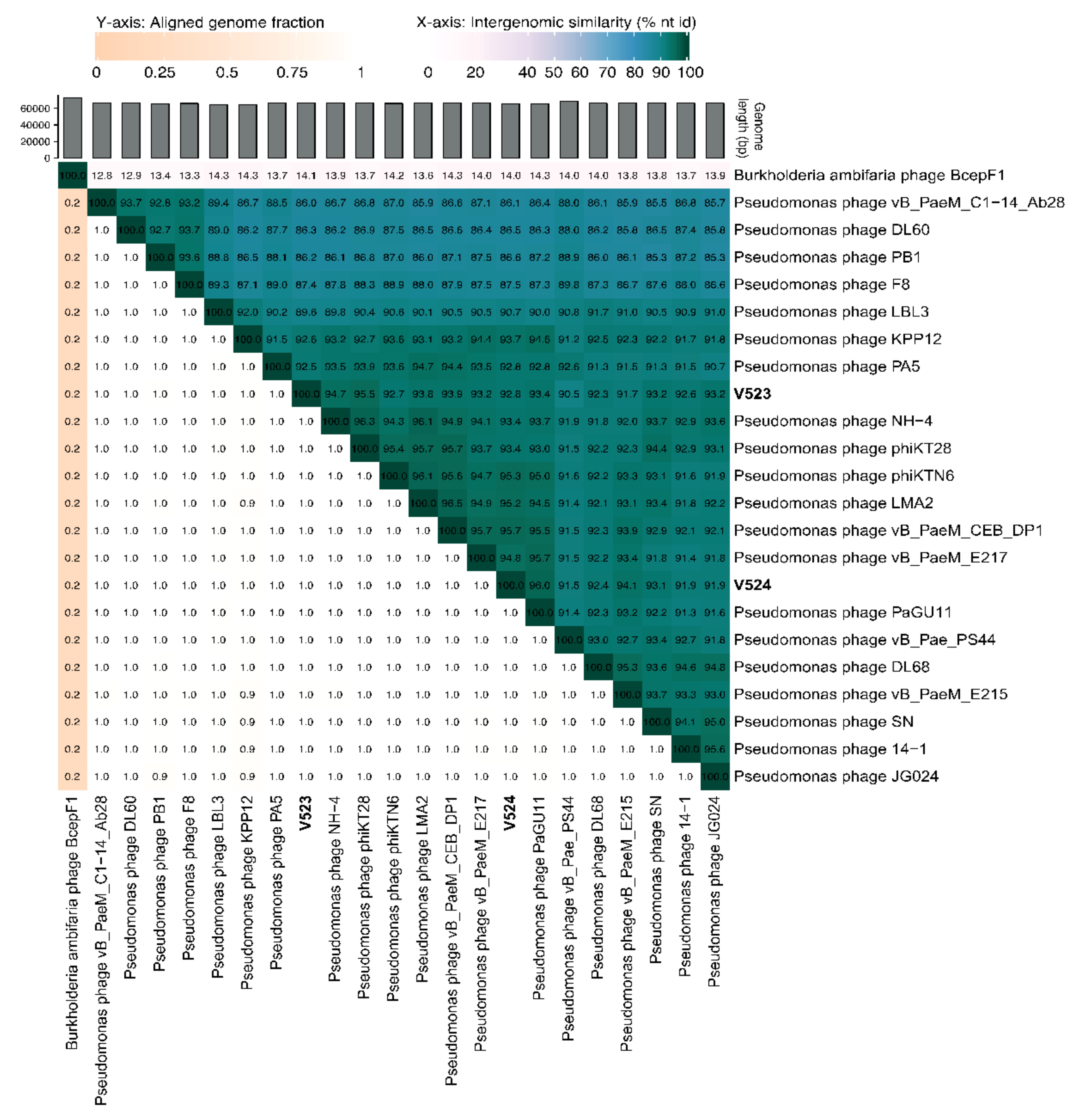
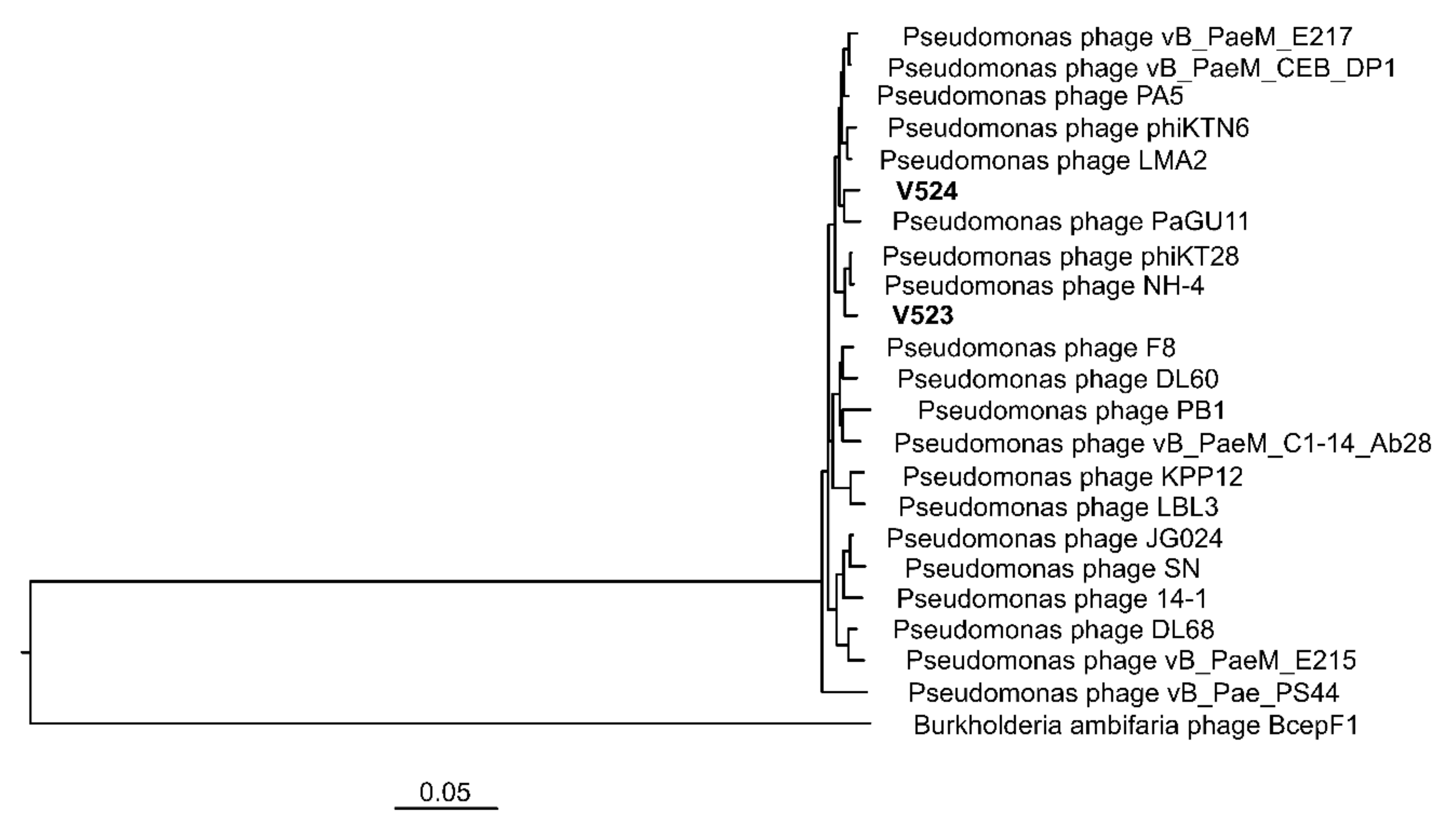
The phages V523 and V524 had genome sizes of 66,301 and 65,447 bp, respectively. They were both assembled into circular genomes, for which the start position of the genome was determined based on a comparison with other closely related phages (Figure 3). The two phages V523 and V524 shared an intergenomic nucleotide identity of 92.8%, identifying them as different species within the same genus (Figure 4). Both V523 and V524 aligned within the Pbunavirus genus (Figure 4 and Figure 5). Here, V523 was the most similar to Pseudomonas phage phiKT28, sharing an intergenomic nucleotide identity of 95.5%. In turn, V523 and the Pseudomonas phage phiKT28 shared 94.7% and 96.3% nucleotide identity, respectively, with Pseudomonas phage NH-4, which is the type phage for the species Pseudomonas virus NH4. Phage V524 shared 96.0% nucleotide identity with Pseudomonas phage PaGU11. Both V524 and Pseudomonas phage PaGU11 shared a high nucleotide identity with Pseudomonas phage vB_PaeM_E217 (94.8% and 95.7%, respectively) and Pseudomonas phage vB_PaeM_CEB_DP1 (95.7% and 95.5%, respectively).
For V523, 94 ORFs were detected and 90 ORFs were detected for V524. Of these, 38 encoded proteins could be annotated to a putative function within both genomes. These mainly included functions involved in DNA replication and proteins forming the structural virion (Figure 3). Both V523 and V524 contained a predicted thymidylate synthase, which is common among Pbunaviruses. No genes suggesting lysogenic replication, antibiotic resistance or toxin production were detected.
3.5. Phage Biocontrol of P. aeruginosa in Water
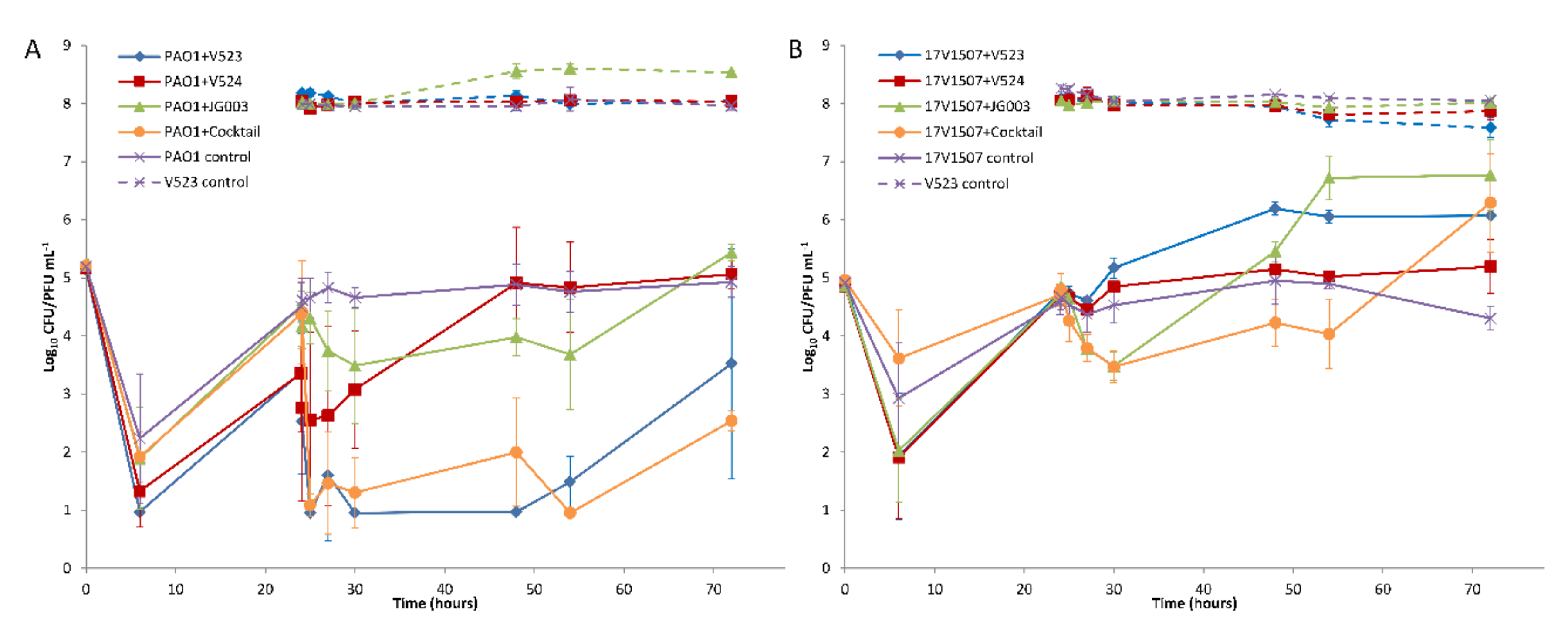
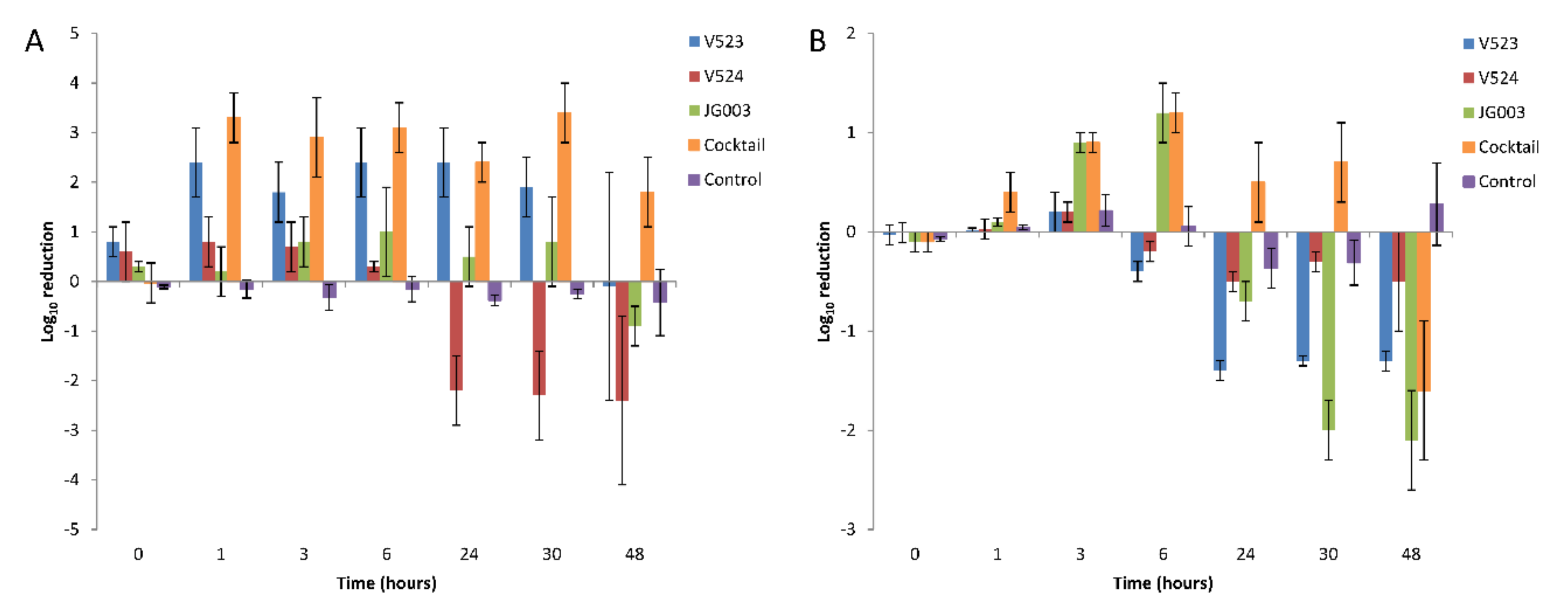
The hosts were allowed to adapt to the test water for 24 h prior to the addition of the phages. During this time, the numbers of both strains decreased and then recovered to near initial levels (Figure 6), except in tests done with PAO1 treated with phages V523 and V524, in which the levels remained about 1 log lower (Figure 6A). With both host strains, the effect of the phage addition was fast and could be seen within six hours after the application. With the PAO1 strain, phage V523 was the most efficient and produced a statistically significant >2.4 ± 0.7 log10 reduction compared to the control (p = 0.018, n = 7) (Figure 7A). The phage cocktail also produced a statistically significant reduction in the PAO1 strain compared to the control (p = 0.018, n = 7), with the highest log10 reduction of >3.4 ± 0.6 measured 30 h post-infection. The log10 reductions achieved with phage V523 and the cocktail did not differ statistically. The reductions observed with phage V524 and JG003 were restored and the phage counts returned to near-control levels within two days (Figure 6A).
With the environmental 17V1507 host, phage JG003 was the most efficient of the individual phages, showing a 1.2 ± 0.3 log10 reduction within six hours (Figure 6B and Figure 7B). The phage cocktail achieved an equal reduction within six hours (1.2 ± 0.2). The reductions in the 17V1507 strain observed with phage JG003 and the cocktail were restored, and the numbers ended up being higher compared to the control levels. The replication of the 17V1507 host was slower in the cocktail compared to JG003, as evidenced by the significant statistical difference between the log10 reductions of phage JG003 and the cocktail (p = 0.043, n = 7). The slight reductions observed with phage V523 and V524 were restored and the host counts ended up being higher compared to the control.
The PAO1 strain was more susceptible than the environmental 17V1507 strain to phage V523 (p = 0.018, n = 7) and the phage cocktail (p = 0.018, n = 7).
3.6. P. aeruginosa Resistance
The hosts treated with phages produced resistance during the experiment. Initially, the colonies produced on TSA plates showed variations in size and morphology (small colonies with irregular shapes) but became more regular in shape as the experiment progressed. The use of the phage cocktail reduced the development of resistance with both hosts compared to the use of individual phages (Figure 6 and Figure 7). The PAO1 resistance to phage V524 was stable after three consecutive passages on TSA plates. Resistance to phage V524 did not protect the PAO1 host against the other two phages (V523 and JG003).
4. Discussion
In this study, two P. aeruginosa phages were isolated from wastewater. The physical and genome characterization revealed that these phages belonged to the genus Pbunavirus, which constitute environmentally very widespread phages within the Myoviridae family [43]. The two isolated phages (V523 and V524) showed high similarity in their morphology, growth characteristics (latent period and burst size) and genome structure. The sequence identity between phages V523 and V524 was 92.8%. Phage V523 shared the highest nucleotide identity (95.5%) with Pseudomonas phage phiKT28 [44] and phage V524 had the highest identity with Pseudomonas phage PaGU11 (96%) (accession: AP018815). In turn, Pseudomonas phage phiKT28 and V524 and Pseudomonas phage PaGU11 shared >95% nucleotide identity with one and two phages, respectively, that represent an individual viral species within the Pbunavirus genus. A 95% nucleotide identity is set as the cut-off defining the species of the phages [45]. However, the micro-diversity among the Pseudomonas phages (Figure 4) makes it difficult to define which species V523 and V524 belong to, as several of the species share >95% nucleotide identity between each other. The genomes of V523 and V524, like the Pbunavirus phages we compared them to (Figure 3) [46], do not encode a recognizable integrase, toxin genes or antibiotic resistance genes, supporting their suitably for application in phage therapy experiments.
The studied phages were lytic against a broad range of P. aeruginosa hosts. The phage infectivity, tested with clinical and environmental P. aeruginosa strains (n = 36), revealed that 67% and 83% of the hosts were sensitive to phage V523 and V524, respectively. JG003 had the broadest host range of the three phages, lysing 97% of the strains. JG003 belongs to the phA group (phenotypically) and originates from sewage [47] (no genome data available). For the biocontrol applications, lytic phages with a broad host range are preferred and recommended. Moreover, growth characteristics are important determinants for efficient biocontrol, and in our study, a longer latent period (55 vs. 35 min), but a much higher burst size (124 vs. 65 PFU/infected cell) was observed with phage V523 in comparison to its closest relative phage KT28 [44]. Overall, the large burst size and relatively short latent period are desirable characteristics for a phage used in biocontrol applications [48].
This study describes the potential of using bacteriophages as biocontrol agents in water. The results showed that phages can control the growth and numbers of planktonic bacteria in water. The interaction was relatively fast and the highest log reductions were observed within 6 h. Differences in efficiency were noted between the phages against the two P. aeruginosa hosts in water. Of the three phages, V523 produced a significant >2.4 log10 reduction in the numbers of the PAO1 host during the 48-h challenge test. However, the reduction was reversed between 30 h and 48 h after the addition of V523. A similar loss of reduction was not observed with the phage cocktail. Phage JG003 caused a reduction in the numbers of both hosts (PAO1 and 17V1507, 1.0 and 1.2 log10 reductions, respectively) during the first 6 h after the spike, but the counts were restored during the following 48 h. Phage V524 had a poor effect in the biocontrol study. A cocktail of three phages showed a slightly higher decrease in the level of the hosts compared to the use of individual phages. The highest observed log10 reduction of >3.4 was achieved with a cocktail of three phages in the PAO1 host. The greater reduction of the PAO1 observed with the cocktail (>3.4 log) than with the phage V523 (>2.4) was probably due to the higher initial number of hosts before the spike in the test carried out with the cocktail. However, no clear synergistic effect was observed with the use of the cocktail, i.e., the cocktail did not produce higher reductions compared to the sum of the reductions of the three individual phages. Synergy has been described as one important reason for using cocktails in phage therapy [49].
It has been assumed that planktonic cells are easier targets for phage control than cells protected by biofilms [50,51]. A previous study showed that phages can eliminate up to 90% of P. aeruginosa strains (including PAO1) in a liquid medium [52]. More studies have been carried out with P. aeruginosa biofilms (reviewed in Chegini et al. [53]). Pires et al. [54] demonstrated that the administration of bacteriophages to bacterial biofilm cultures achieved a 2–3 log10 reduction in the P. aeruginosa cell number. Similarly, Magin et al. [52] showed a maximum reduction of 1.7 log10 in a biofilm, using viability-qPCR. These reductions are consistent with our results.
The reference strain PAO1 was more susceptible to the phages than the environmental strain 17V1507. This is consistent with the spot test results, in which PAO1 produced clear lysis zones for all three phages, and as with the 17V1507 strain, turbid plaques were observed with phage V524 and JG003. PAO1 also showed higher variation than 17V1507 between the replicates during the 72-h tests in water. These observations may be partly due to the different origin of the PAO1 strain (an infected wound) compared to the environmental strain 17V1507, isolated from swimming pool water. With strain 17V1507, better results could have been achieved with isolating phages with higher activity against this environmental isolate. Overall, the results clearly indicate that the careful selection of individual phages against a target host and the design of the phage cocktail are important. Moreover, the use of a same phage cocktail for different host strains is not recommended prior to further testing.
The results clearly show that bacteria can develop phage resistance in water. In this study, resistance was evident, since even though the colony counts of the bacterial hosts decreased (even to below detection-limit values) after phage addition, growth of the hosts was observed towards the end of the test period. This growth in spiked waters may be favored by the nutrients that came with the phage spike, since in the controls this growth was not observed. The emergence of phage resistance has been detected previously with P. aeruginosa [44,54,55]. Our study indicated that the resistance was stable after three consecutive passages on TSA plates. It was noteworthy that PAO1 resistance to phage V524 did not protect the host against other two phages (V523 and JG003). However, phage resistance usually comes at the cost of cell fitness [56,57]. In the present study, this was observed on the agar plates, which initially produced atypical colony morphologies (smaller colonies and/or a star-like shape), but later normal colonies (round) increased on the plates. In clinical settings, it is assumed that the host does not need to be killed totally since the immune system will take care of weakened pathogens. This is a prerequisite for a successful phage therapy because the total destruction of the host is rarely possible. Positive results have been documented, e.g., using phages in the treatment of antibiotic-resistant P. aeruginosa in chronic otitis [58]. There are also reports showing synergistic effects using phages and antibiotics in medical applications [59,60]. Similarly, in water environments, the phage-treated weakened pathogens are thought to be more susceptible to disinfectants [61]. This could allow the use of lower levels of chemicals and could enable the efficient disinfection of microbes which are tolerant of chemicals. The use of the phage cocktail clearly reduced the development of resistance. Therefore, the use of a cocktail is highly recommended in future applications in water.
5. Conclusions
The encouraging results obtained with the P. aeruginosa biocontrol in water emphasize the potential of using phages to control harmful bacteria. New control options are needed since antibiotic-resistant bacteria, such as multidrug-resistant P. aeruginosa, may spread in water environments and become a potential health threat if they are present in drinking water or recreational water such as in swimming pools and spas. Biocontrol in water may be sufficient to reduce the number of pathogenic bacteria below acceptable levels, at which there is no expected risk to health, and to inhibit their growth. However, more knowledge is needed in several respects related to phage use as a biocontrol agent in water, including the safety, efficiency, resistance, use and design of phage cocktails, the use of phage and disinfectants together and the role of matrices (water vs. biofilms). The main conclusions of this study are as follows:
- Lytic P. aeruginosa phages with a broad host range were isolated and characterized for biocontrol purposes in water.
- Tests conducted in water showed differences between the efficiency of phages and the sensitivity of hosts.
- A three-phage cocktail showed more consistent and efficient reduction of hosts compared to the use of a single phage.
- Phage resistance appeared rapidly and its control requires further studies.
- Careful design of the phage cocktail is important in order to achieve maximum reductions and to curtail possible phage resistance.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/v13050928/s1, Table S1: Information regarding the accession number and species affiliation for the Pbunavirus phages used for genome comparison in Figure 3, Figure 4 and Figure 5. Table S2: Information about the gene arrangement and functional annotation of the two phages, V523 and V524, as well as best match to viralDB and PFAM.
Author Contributions
Conceptualization, A.K., S.S., T.P., E.T. and I.T.M.; methodology, A.K. and K.H.; formal analysis, A.K. and K.H.; investigation, A.K., S.S. and A.P.; writing—original draft preparation, A.K. and K.H.; writing—review and editing, A.K., S.S., T.P., K.H., E.T. and I.T.M.; visualization, A.K. and K.H.; funding acquisition, E.T. and I.T.M. All authors have read and agreed to the published version of the manuscript.
Funding
The study was performed as part of the DWDSOME (Microbiome Dynamics in Drinking Water Distribution System) project, funded by the Academy of Finland (Number 275549/2014). This work was also supported by the Olvi foundation (grant number 201810730) and Maa- ja vesitekniikan tuki ry (grant number 41713) awarded to S.S.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are included in the article and the Supplementary Materials. The viral genomes were deposited in the NCBI GenBank with the accession numbers MW595220 and MW595221.
Acknowledgments
Ulla Kärkkäinen from the Eastern Finland Laboratory Centre Joint Authority Enterprise (ISLAB), is acknowledged for providing the bacterial strains for the host range study. We also acknowledge the help of the personnel from the Expert Microbiology unit of the Finnish Institute for Health and Welfare. Furthermore, we would like to acknowledge support from the National Genomics Infrastructure funded by the Science for Life Laboratory, the Knut and Alice Wallenberg Foundation and the Swedish Research Council, and the SNIC/Uppsala Multidisciplinary Center for Advanced Computational Science for assistance with massively parallel sequencing and access to the UPPMAX computational infrastructure. The data handling was enabled using resources provided by the Swedish National Infrastructure for Computing (SNIC) at UPPMAX, partially funded by the Swedish Research Council through grant agreement No. 2018-05973.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage treatment of human infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Calle, D.; Guimarães Benevides, R.; Góes-Neto, A.; Billington, C. Bacteriophages as Alternatives to Antibiotics in Clinical Care. Antibiotics 2019, 8, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dy, R.L.; Rigano, L.A.; Fineran, P.C. Phage-based biocontrol strategies and their application in agriculture and aquaculture. Biochem. Soc. Trans. 2018, 46, 1605–1613. [Google Scholar] [CrossRef] [PubMed]
- Soliman, W.S.; Shaapan, R.M.; Mohamed, L.A.; Gayed, S.S.R. Recent biocontrol measures for fish bacterial diseases, in particular to probiotics, bio-encapsulated vaccines, and phage therapy. Open Vet. J. 2019, 9, 190–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, J.W.; Park, S.C.; Wicklund, A.; Skurnik, M. Bacteriophages reduce Yersinia enterocolitica contamination of food and kitchenware. Int. J. Food Microbiol. 2018, 271, 33–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage Applications for Food Production and Processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, C.; Yamada, K.; Takeuchi, H.; Inokuchi, Y.; Kashiwagi, A.; Toba, T. A Lytic Bacteriophage for Controlling Pseudomonas lactis in Raw Cow’s Milk. Appl. Environ. Microbiol. 2018, 84, e00111-18. [Google Scholar] [CrossRef] [Green Version]
- Goldman, G.; Starosvetsky, J.; Armon, R. Inhibition of biofilm formation on UF membrane by use of specific bacteriophages. J. Membr. Sci. 2009, 342, 145–152. [Google Scholar] [CrossRef]
- Turki, Y.; Ouzari, H.; Mehri, I.; Ammar, A.B.; Hassen, A. Evaluation of a cocktail of three bacteriophages for the biocontrol of Salmonella of wastewater. Food Res. Int. 2012, 45, 1099–1105. [Google Scholar] [CrossRef]
- Malaeb, L.; Le-Clech, P.; Vrouwenvelder, J.S.; Ayoub, G.M.; Saikaly, P.E. Do biological-based strategies hold promise to biofouling control in MBRs? Water Res. 2013, 47, 5447–5463. [Google Scholar] [CrossRef]
- Zhang, Y.; Hunt, H.K.; Hu, Z. Application of bacteriophages to selectively remove Pseudomonas aeruginosa in water and wastewater filtration systems. Water Res. 2013, 47, 4507–4518. [Google Scholar] [CrossRef]
- Mathieu, J.; Yu, P.; Zuo, P.; Da Silva, M.L.B.; Alvarez, P.J.J. Going Viral: Emerging Opportunities for Phage-Based Bacterial Control in Water Treatment and Reuse. Acc. Chem. Res. 2019, 52, 849–857. [Google Scholar] [CrossRef]
- Reyneke, B.; Khan, S.; Fernández-Ibáñezc, P.; Khan, W. Podoviridae bacteriophage for the biocontrol of Pseudomonas aeruginosa in rainwater. Environ. Sci. Water Res. Technol. 2020, 6, 87–102. [Google Scholar] [CrossRef]
- Fondi, M.; Karkman, A.; Tamminen, M.V.; Bosi, E.; Virta, M.; Fani, R.; Alm, E.; McInerney, J.O. “Every gene is everywhere but the environment selects”: Global geolocalization of gene sharing in environmental samples through network analysis. Genome Biol. Evol. 2016, 8, 1388–1400. [Google Scholar] [CrossRef]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Baquero, F.; Martínez, J.L.; Cantón, R. Antibiotics and antibiotic resistance in water environments. Curr. Opin. Biotechnol. 2008, 19, 260–265. [Google Scholar] [CrossRef]
- Karkman, A.; Johnson, T.A.; Lyra, C.; Stedtfeld, R.D.; Tamminen, M.; Tiedje, J.M.; Virta, M. High-throughput quantification of antibiotic resistance genes from an urban wastewater treatment plant. FEMS Microbiol. Ecol. 2016, 92, fiw014. [Google Scholar] [CrossRef] [Green Version]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef]
- Mathee, K.; Narasimhan, G.; Valdes, C.; Qiu, X.; Matewish, J.M.; Koehrsen, M.; Rokas, A.; Yandava, C.N.; Engels, R.; Zeng, E.; et al. Dynamics of Pseudomonas aeruginosa genome evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 3100–3105. [Google Scholar] [CrossRef] [Green Version]
- Moritz, M.M.; Flemming, H.C.; Wingender, J. Integration of Pseudomonas aeruginosa and Legionella pneumophila in drinking water biofilms grown on domestic plumbing materials. Int. J. Hyg. Environ. Health 2010, 213, 190–197. [Google Scholar] [CrossRef]
- Valot, B.; Guyeux, C.; Rolland, J.Y.; Mazouzi, K.; Bertrand, X.; Hocquet, D. What it takes to be a Pseudomonas aeruginosa? the core genome of the opportunistic pathogen updated. PLoS ONE 2015, 10, e0126468. [Google Scholar] [CrossRef]
- Bert, F.; Maubec, E.; Bruneau, B.; Berry, P.; Lambert-Zechovsky, N. Multi-resistant Pseudomonas aeruginosa outbreak associated with contaminated tap water in a neurosurgery intensive care unit. J. Hosp. Infect. 1998, 39, 53–62. [Google Scholar] [CrossRef]
- Eckmanns, T.; Oppert, M.; Martin, M.; Amorosa, R.; Zuschneid, I.; Frei, U.; Rüden, H.; Weist, K. An outbreak of hospital-acquired Pseudomonas aeruginosa infection caused by contaminated bottled water in intensive care units. Clin. Microbiol. Infect. 2008, 14, 454–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naze, F.; Jouen, E.; Randriamahazo, R.T.; Simac, C.; Laurent, P.; Blériot, A.; Chiroleu, F.; Gagnevin, L.; Pruvost, O.; Michault, A. Pseudomonas aeruginosa outbreak linked to mineral water bottles in a neonatal intensive care unit: Fast typing by use of high-resolution melting analysis of a variable-number tandem-repeat locus. J. Clin. Microbiol. 2010, 48, 3146–3152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bicking Kinsey, C.; Koirala, S.; Solomon, B.; Rosenberg, J.; Robinson, B.F.; Neri, A.; Laufer Halpin, A.; Arduino, M.J.; Moulton-Meissner, H.; Noble-Wang, J.; et al. Pseudomonas aeruginosa Outbreak in a Neonatal Intensive Care Unit Attributed to Hospital Tap Water. Infect. Control Hosp. Epidemiol. 2017, 38, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.K.; Brooks, R.B.; Glowicz, J.; Keckler, M.S.; Christensen, B.E.; Tsai, V.; Mitchell, C.S.; Wilson, L.E.; Laxton, R.; Moulton-Meissner, H.; et al. Outbreak investigation of Pseudomonas aeruginosa infections in a neonatal intensive care unit. Am. J. Infect. Control 2019, 47, 1148–1150. [Google Scholar] [CrossRef] [PubMed]
- Brousseau, N.; Lévesque, B.; Guillemet, T.A.; Cantin, P.; Gauvin, D.; Giroux, J.P.; Gingras, S.; Proulx, F.; Côté, P.A.; Dewailly, E. Contamination of public whirlpool spas: Factors associated with the presence of Legionella spp., Pseudomonas aeruginosa and Escherichia coli. Int. J. Environ. Health Res. 2013, 23, 1–15. [Google Scholar] [CrossRef]
- Guida, M.; Di Onofrio, V.; Gallè, F.; Gesuele, R.; Valeriani, F.; Liguori, R.; Romano Spica, V.; Liguori, G. Pseudomonas aeruginosa in Swimming Pool Water: Evidences and Perspectives for a New Control Strategy. Int. J. Environ. Res. Public Health 2016, 13, 919. [Google Scholar] [CrossRef] [Green Version]
- Germinario, C.; Tafuri, S.; Napoli, C.; Martucci, M.; Termite, S.; Pedote, P.; Montagna, M.T.; Quarto, M. An outbreak of pneumonia in a thermal water spa contaminated with Pseudomonas aeruginosa: An epidemiological and environmental concern. Afr. J. Microbiol. Res. 2012, 6, 1978–1984. [Google Scholar] [CrossRef]
- Rice, S.A.; van den Akker, B.; Pomati, F.; Roser, D. A risk assessment of Pseudomonas aeruginosa in swimming pools: A review. J. Water Health 2012, 10, 181–196. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Beattie, T.K.; Knapp, C.W. Relationship between antibiotic- and disinfectant-resistance profiles in bacteria harvested from tap water. Chemosphere 2016, 152, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Hou, A.M.; Yang, D.; Miao, J.; Shi, D.Y.; Yin, J.; Yang, Z.W.; Shen, Z.Q.; Wang, H.R.; Qiu, Z.G.; Liu, W.L.; et al. Chlorine injury enhances antibiotic resistance in Pseudomonas aeruginosa through over expression of drug efflux pumps. Water Res. 2019, 156, 366–371. [Google Scholar] [CrossRef]
- Adams, M.H. Bacteriophages; Interscience Publishers Inc.: New York, NY, USA, 1959. [Google Scholar]
- Azeredo, J.; Sillankorva, S.; Pires, D.P. Pseudomonas Bacteriophage Isolation and Production. In Pseudomonas Methods and Protocols. Methods in Molecular Biology; Filloux, A., Ramos, J.L., Eds.; Humana Press: New York, NY, USA, 2014; Volume 1149, pp. 23–32. [Google Scholar] [CrossRef]
- Ellis, E.L.; Delbruck, M. The growth of bacteriophage. J. Gen. Physiol. 1939, 22, 365–384. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Version 0.7.2; The Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC—A Novel Tool to Calculate the Intergenomic Similarities of Prokaryote-Infecting Viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. VICTOR: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besemer, J.; Borodovsky, M. Heuristic approach to deriving models for gene finding. Nucleic Acids Res. 1999, 27, 3911–3920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Ceyssens, P.-J.; Miroshnikov, K.; Mattheus, W.; Krylov, V.; Robben, J.; Noben, J.-P.; Vanderschraeghe, S.; Sykilinda, N.; Kropinski, A.M.; Volckaert, G.; et al. Comparative analysis of the widespread and conserved PB1-like viruses infecting Pseudomonas aeruginosa. Environ. Microbiol. 2009, 11, 2874–2883. [Google Scholar] [CrossRef]
- Danis-Wlodarczyk, K.; Olszak, T.; Arabski, M.; Wasik, S.; Majkowska-Skrobek, G.; Augustyniak, D.; Gula, G.; Briers, Y.; Jang, H.B.; Vandenheuvel, D.; et al. Characterization of the Newly Isolated Lytic Bacteriophages KTN6 and KT28 and Their Efficacy against Pseudomonas aeruginosa Biofilm. PLoS ONE 2015, 10, e0127603. [Google Scholar] [CrossRef]
- Adriaenssens, E.; Brister, J.R. How to Name and Classify Your Phage: An Informal Guide. Viruses 2017, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Alemayehu, D.; Casey, P.G.; McAuliffe, O.; Guinane, C.M.; Martin, J.G.; Shanahan, F.; Coffey, A.; Ross, R.P.; Hill, C. Bacteriophages φMR299-2 and φNH-4 can eliminate Pseudomonas aeruginosa in the murine lung and on cystic fibrosis lung airway cells. mBio 2012, 3, e00029-12. [Google Scholar] [CrossRef] [Green Version]
- Selezska, K.; Kazmierczak, M.; Müsken, M.; Garbe, J.; Schobert, M.; Häussler, S.; Wiehlmann, L.; Rohde, C.; Sikorski, J. Pseudomonas aeruginosa population structure revisited under environmental focus: Impact of water quality and phage pressure. Environ. Microbiol. 2012, 14, 1952–1967. [Google Scholar] [CrossRef]
- Gill, J.J.; Hyman, P. Phage choice, isolation, and preparation for phage therapy. Curr. Pharm. Biotechnol. 2010, 11, 2–14. [Google Scholar] [CrossRef]
- Schmerer, M.; Molineux, I.J.; Bull, J.J. Synergy as a rationale for phage therapy using phage cocktails. PeerJ 2014, 2, e590. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, R.S.; Svenningsen, S.L.; Sneppen, K.; Mitarai, N. A growing microcolony can survive and support persistent propagation of virulent phages. Proc. Natl. Acad. Sci. USA 2018, 115, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Testa, S.; Berger, S.; Piccardi, P.; Oechslin, F.; Resch, G.; Mitri, S. Spatial structure affects phage efficacy in infecting dual-strain biofilms of Pseudomonas aeruginosa. Commun. Biol. 2019, 2, 405. [Google Scholar] [CrossRef] [Green Version]
- Magin, V.; Garrec, N.; Andrés, Y. Selection of Bacteriophages to Control In Vitro 24 h Old Biofilm of Pseudomonas Aeruginosa Isolated from Drinking and Thermal Water. Viruses 2019, 11, 749. [Google Scholar] [CrossRef] [Green Version]
- Chegini, Z.; Khoshbayan, A.; Taati Moghadam, M.; Farahani, I.; Jazireian, P.; Shariati, A. Bacteriophage therapy against Pseudomonas aeruginosa biofilms: A review. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 45. [Google Scholar] [CrossRef]
- Pires, D.; Sillankorva, S.; Faustino, A.; Azeredo, J. Use of newly isolated phages for control of Pseudomonas aeruginosa PAO1 and ATCC 10145 biofilms. Res. Microbiol. 2011, 162, 798–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forti, F.; Roach, D.R.; Cafora, M.; Pasini, M.E.; Horner, D.S.; Fiscarelli, E.V.; Rossitto, M.; Cariani, L.; Briani, F.; Debarbieux, L.; et al. Design of a Broad-Range Bacteriophage Cocktail That Reduces Pseudomonas aeruginosa Biofilms and Treats Acute Infections in Two Animal Models. Antimicrob. Agents Chemother. 2018, 62, e02573-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef]
- Oechslin, F. Resistance Development to Bacteriophages Occurring during Bacteriophage Therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.; Hawkins, C.H.; Anggård, E.E.; Harper, D.R. A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa; a preliminary report of efficacy. Clin. Otolaryngol. 2009, 34, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Oechslin, F.; Piccardi, P.; Mancini, S.; Gabard, J.; Moreillon, P.; Entenza, J.M.; Resch, G.; Que, Y.-A. Synergistic Interaction Between Phage Therapy and Antibiotics Clears Pseudomonas aeruginosa Infection in Endocarditis and Reduces Virulence. J. Infect. Dis. 2017, 215, 703–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akturk, E.; Oliveira, H.; Santos, S.B.; Costa, S.; Kuyumcu, S.; Melo, L.D.R.; Azeredo, J. Synergistic Action of Phage and Antibiotics: Parameters to Enhance the Killing Efficacy Against Mono and Dual-Species Biofilms. Antibiotics 2019, 8, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agún, S.; Fernández, L.; González-Menéndez, E.; Martínez, B.; Rodríguez, A.; García, P. Study of the Interactions Between Bacteriophage phiIPLA-RODI and Four Chemical Disinfectants for the Elimination of Staphylococcus aureus Contamination. Viruses 2018, 10, 103. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Transmission electron microscopic images of phages (A) V523 and (B) V524. The scale bars are 100 nm.
Figure 1.
Transmission electron microscopic images of phages (A) V523 and (B) V524. The scale bars are 100 nm.

Figure 2.
One-step growth curves of the phage (A) V523 and (B) V524. The error bars represent the standard deviations for technical triplicates. The latent periods and the burst sizes are indicated.
Figure 2.
One-step growth curves of the phage (A) V523 and (B) V524. The error bars represent the standard deviations for technical triplicates. The latent periods and the burst sizes are indicated.

Figure 3.
A synteny plot displaying the genome structure of phages V523 and V524, as well as their identity to other closely related Pbunavirus phages. The figure was created using Easyfig [42] and the genomes are displayed linearly for visualization. Detailed information regarding the gene annotation can be found in Supplementary Table S1.
Figure 3.
A synteny plot displaying the genome structure of phages V523 and V524, as well as their identity to other closely related Pbunavirus phages. The figure was created using Easyfig [42] and the genomes are displayed linearly for visualization. Detailed information regarding the gene annotation can be found in Supplementary Table S1.

Figure 4.
A heat map displaying the similarity between V523 and V524 to reference genomes within the Pbunavirus genus and closely related published Pbunavirus phage, created using VIRIDIC [39]. Accession numbers for previous phage isolates can be found in Supplementary Table S1.
Figure 4.
A heat map displaying the similarity between V523 and V524 to reference genomes within the Pbunavirus genus and closely related published Pbunavirus phage, created using VIRIDIC [39]. Accession numbers for previous phage isolates can be found in Supplementary Table S1.

Figure 5.
A VICTOR tree produced with FastME based on nucleotide identity calculated with the D0-formula [40], with the newly isolated phages (V523 and V524) and reference genomes within the Pbunavirus genus and closely related published Pbunavirus phage strains. Accession numbers for previous phage isolates can be found in Supplementary Table S1.
Figure 5.
A VICTOR tree produced with FastME based on nucleotide identity calculated with the D0-formula [40], with the newly isolated phages (V523 and V524) and reference genomes within the Pbunavirus genus and closely related published Pbunavirus phage strains. Accession numbers for previous phage isolates can be found in Supplementary Table S1.

Figure 6.
The effect of phages on (A) the P. aeruginosa PAO1 strain and (B) the environmental strain 17V1507. Three separate phages and the phage cocktail were tested. The phages were added after 24 h. Host (solid lines) and phage (dashed lines) numbers were determined from triplicate experiments, and the error bars indicate standard deviations.
Figure 6.
The effect of phages on (A) the P. aeruginosa PAO1 strain and (B) the environmental strain 17V1507. Three separate phages and the phage cocktail were tested. The phages were added after 24 h. Host (solid lines) and phage (dashed lines) numbers were determined from triplicate experiments, and the error bars indicate standard deviations.

Figure 7.
Average log10 reductions of (A) the P. aeruginosa PAO1 strain and (B) the environmental strain 17V1507 using bacteriophages in water. Three separate phages and a phage cocktail were tested. Host reductions were determined from triplicate experiments, and the error bars indicate standard deviations.
Figure 7.
Average log10 reductions of (A) the P. aeruginosa PAO1 strain and (B) the environmental strain 17V1507 using bacteriophages in water. Three separate phages and a phage cocktail were tested. Host reductions were determined from triplicate experiments, and the error bars indicate standard deviations.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Averages and standard deviations of physical-chemical properties of the test water (n = 4).
Table 1.
Averages and standard deviations of physical-chemical properties of the test water (n = 4).
| Parameter | Drinking Water |
|---|---|
| pH | 7.3 ± 0.3 |
| Conductivity, μS cm−1 | 446 ± 8 |
| AOC 1, μg AOC-C L−1 | 113 ± 70 |
| AOC, μg acetate-C L−1 | 74 ± 47 |
| MAP 2, μg MAP-P L−1 | 1.5 ± 0.6 |
1 AOC, assimilable organic carbon; 2 MAP, microbially available phosphorus.
Table 2.
Host range of individual phages V523, V524 and JG003.
| Host Strain | Source | V523 | V524 | JG003 |
|---|---|---|---|---|
| PAO1 (DSM 19880) | Clinical | ++ | ++ | ++ |
| 17V1507 | Swimming pool | − | + | + |
| 17V1508 | Swimming pool | − | + | + |
| 17V1509 | Swimming pool | − | ++ | ++ |
| 17V1510 | Swimming pool | − | + | ++ |
| 17V1511 | Swimming pool | − | + | + |
| 17V1512 | Swimming pool | − | + | ++ |
| 18V0528/1b | Swimming pool | + | + | + |
| 19V2041 | Swimming pool | + | + | ++ |
| 17V427/2a | Wastewater | + | ++ | + |
| 17V427/7a | Wastewater | + | ++ | + |
| 17V427/9a | Wastewater | + | ++ | + |
| 19V2030 | Water tank | + | ++ | + |
| 19V2031 | Water tank | + | ++ | + |
| 19V2032 | Water tank | + | ++ | + |
| 19V2033 | Water tank | + | + | + |
| 19V2034 | Water tank | + | + | + |
| 19V2035 | Water tank | − | ++ | ++ |
| 19V2036 | Water tank | + | ++ | + |
| 19V2037 | Water tank | + | ++ | + |
| 19V2038 | Water tank | + | ++ | + |
| 19V2039 | Water tank | ++ | + | + |
| 19V2040 | Water tank | + | + | ++ |
| 19V2042 | Water tank | + | ++ | + |
| 19V1974 | Clinical | − | ++ | ++ |
| 19V1976 | Clinical | + | − | + |
| 19V1977 | Clinical | + | − | ++ |
| 19V1978 | Clinical | − | − | ++ |
| 19V1979 | Clinical | ++ | + | − |
| 19V1980 | Clinical | ++ | − | ++ |
| 19V1981 | Clinical | ++ | − | + |
| 19V1982 | Clinical | + | − | ++ |
| 19V1983 | Clinical | + | ++ | + |
| 19V1984 | Clinical | − | + | + |
| ATCC 27853 | Clinical | − | + | + |
| DSM 50071 | Not known | − | + | + |
| # of sensitive strains | 24/36 (67%) | 30/36 (83%) | 35/36 (97%) | |
| # of sensitive strains (clear) | 5/36 (14%) | 15/36 (42%) | 12/36 (33%) |
++, clear plaque (complete lysis); +, turbid plaque (partial lysis); −, no plaque (no lysis); #, number.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kauppinen, A.; Siponen, S.; Pitkänen, T.; Holmfeldt, K.; Pursiainen, A.; Torvinen, E.; Miettinen, I.T. Phage Biocontrol of Pseudomonas aeruginosa in Water. Viruses 2021, 13, 928. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050928
AMA Style
Kauppinen A, Siponen S, Pitkänen T, Holmfeldt K, Pursiainen A, Torvinen E, Miettinen IT. Phage Biocontrol of Pseudomonas aeruginosa in Water. Viruses. 2021; 13(5):928. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050928
Chicago/Turabian StyleKauppinen, Ari, Sallamaari Siponen, Tarja Pitkänen, Karin Holmfeldt, Anna Pursiainen, Eila Torvinen, and Ilkka T. Miettinen. 2021. "Phage Biocontrol of Pseudomonas aeruginosa in Water" Viruses 13, no. 5: 928. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050928
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.