
Dynamics and Extent of Non-Structural Protein 1-Antibody Responses in Tick-Borne Encephalitis Vaccination Breakthroughs and Unvaccinated Patients


Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Serum Samples
2.2. Generation of Recombinant TBE NS1
2.3. Virus Growth, Purification and Inactivation
2.4. NS1 IgG ELISA
2.5. Virion IgG ELISA
2.6. Statistical Analyses
3. Results
3.1. NS1 IgG Antibodies in TBE Patients and Vaccinees
3.2. Comparison of Anti-NS1 IgG Responses in TBE Patients with and without Prior Vaccination
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Vaccines against tick-borne encephalitis: WHO position paper. Wkly. Epidemiol. Rec. 2011, 86, 241–256. [Google Scholar]
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Hombach, J.; Barrett, A.D.T.; Kollaritsch, H. Tickborne encephalitis vaccines. In Plotkin’s Vaccines, 7th ed.; Orenstein, W.A., Offit, P.A., Edwards, K.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1080–1094.e5. [Google Scholar]
- Heinz, F.X.; Stiasny, K.; Holzmann, H.; Grgic-Vitek, M.; Kriz, B.; Essl, A.; Kundi, M. Vaccination and tick-borne encephalitis, central Europe. Emerg. Infect. Dis. 2013, 19, 69–76. [Google Scholar] [CrossRef]
- Stiasny, K.; Holzmann, H.; Heinz, F.X. Characteristics of antibody responses in tick-borne encephalitis vaccination breakthroughs. Vaccine 2009, 27, 7021–7026. [Google Scholar] [CrossRef] [PubMed]
- Andersson, C.R.; Vene, S.; Insulander, M.; Lindquist, L.; Lundkvist, A.; Gunther, G. Vaccine failures after active immunisation against tick-borne encephalitis. Vaccine 2010, 28, 2827–2831. [Google Scholar] [CrossRef]
- Lotrič-Furlan, S.; Bogovič, P.; Avšič-Županc, T.; Jelovšek, M.; Lusa, L.; Strle, F. Tick-borne encephalitis in patients vaccinated against this disease. J. Intern. Med. 2017, 282, 142–155. [Google Scholar] [CrossRef] [Green Version]
- Dobler, G.; Kaier, K.; Hehn, P.; Böhmer, M.M.; Kreusch, T.M.; Borde, J.P. Tick-borne encephalitis virus vaccination breakthrough infections in Germany: A retrospective analysis from 2001 to 2018. Clin. Microbiol. Infect. 2020, 26, e7–e13. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.N.; Sonnberger, M.; Troescher, A.; Krehan, I.; Hauser, A.; Panholzer, J.; von Oertzen, T.J. Patients with breakthrough tick-borne encephalitis suffer a more severe clinical course and display extensive magnetic resonance imaging changes. Eur. J. Neurol. 2020, 27, 1201–1209. [Google Scholar] [CrossRef]
- Watterson, D.; Modhiran, N.; Young, P.R. The many faces of the flavivirus NS1 protein offer a multitude of options for inhibitor design. Antivir. Res. 2016, 130, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Muller, D.A.; Young, P.R. The flavivirus NS1 protein: Molecular and structural biology, immunology, role in pathogenesis and application as a diagnostic biomarker. Antivir. Res. 2013, 98, 192–208. [Google Scholar] [CrossRef] [Green Version]
- Glasner, D.R.; Puerta-Guardo, H.; Beatty, P.R.; Harris, E. The Good, the Bad, and the Shocking: The Multiple Roles of Dengue Virus Nonstructural Protein 1 in Protection and Pathogenesis. Annu. Rev. Virol. 2018, 5, 227–253. [Google Scholar] [CrossRef]
- Albinsson, B.; Rönnberg, B.; Vene, S.; Lundkvist, Å. Antibody responses to tick-borne encephalitis virus non-structural protein 1 and whole virus antigen-a new tool in the assessment of suspected vaccine failure patients. Infect. Ecol. Epidemiol. 2019, 9, 1696132. [Google Scholar] [CrossRef] [PubMed]
- Girl, P.; Bestehorn-Willmann, M.; Zange, S.; Borde, J.P.; Dobler, G.; von Buttlar, H. Tick-Borne Encephalitis Virus Nonstructural Protein 1 IgG Enzyme-Linked Immunosorbent Assay for Differentiating Infection versus Vaccination Antibody Responses. J. Clin. Microbiol. 2020, 58, e01783-19. [Google Scholar] [CrossRef] [PubMed]
- Mora-Cárdenas, E.; Aloise, C.; Faoro, V.; Knap Gašper, N.; Korva, M.; Caracciolo, I.; D’Agaro, P.; Avšič-Županc, T.; Marcello, A. Comparative specificity and sensitivity of NS1-based serological assays for the detection of flavivirus immune response. PLoS Negl. Trop. Dis. 2020, 14, e0008039. [Google Scholar] [CrossRef] [PubMed]
- Salat, J.; Mikulasek, K.; Larralde, O.; Pokorna Formanova, P.; Chrdle, A.; Haviernik, J.; Elsterova, J.; Teislerova, D.; Palus, M.; Eyer, L.; et al. Tick-Borne Encephalitis Virus Vaccines Contain Non-Structural Protein 1 Antigen and may Elicit NS1-Specific Antibody Responses in Vaccinated Individuals. Vaccines 2020, 8, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albinsson, B.; Vene, S.; Rombo, L.; Blomberg, J.; Lundkvist, Å.; Rönnberg, B. Distinction between serological responses following tick-borne encephalitis virus (TBEV) infection vs. vaccination, Sweden 2017. Eurosurveillance 2018, 23, 17-00838. [Google Scholar] [CrossRef] [PubMed]
- Stiasny, K.; Aberle, J.H.; Chmelik, V.; Karrer, U.; Holzmann, H.; Heinz, F.X. Quantitative determination of IgM antibodies reduces the pitfalls in the serodiagnosis of tick-borne encephalitis. J. Clin. Virol. 2012, 54, 115–120. [Google Scholar] [CrossRef]
- Malafa, S.; Medits, I.; Aberle, J.H.; Aberle, S.W.; Haslwanter, D.; Tsouchnikas, G.; Wölfel, S.; Huber, K.L.; Percivalle, E.; Cherpillod, P.; et al. Impact of flavivirus vaccine-induced immunity on primary Zika virus antibody response in humans. PLoS Negl. Trop. Dis. 2020, 14, e0008034. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K.; Johnson, R.A. Maturation of the head of bacteriophage T4. II. Head-related, aberrant tau-particles. J. Mol. Biol. 1973, 80, 601–611. [Google Scholar] [CrossRef]
- Iacono-Connors, L.C.; Smith, J.F.; Ksiazek, T.G.; Kelley, C.L.; Schmaljohn, C.S. Characterization of Langat virus antigenic determinants defined by monoclonal antibodies to E, NS1 and preM and identification of a protective, non-neutralizing preM-specific monoclonal antibody. Virus Res. 1996, 43, 125–136. [Google Scholar] [CrossRef]
- Heinz, F.X.; Kunz, C. Homogeneity of the structural glycoprotein from European isolates of tick-borne encephalitis virus: Comparison with other flaviviruses. J. Gen. Virol. 1981, 57 Pt 2, 263–274. [Google Scholar] [CrossRef]
- Heinz, F.X.; Allison, S.L.; Stiasny, K.; Schalich, J.; Holzmann, H.; Mandl, C.W.; Kunz, C. Recombinant and virion-derived soluble and particulate immunogens for vaccination against tick-borne encephalitis. Vaccine 1995, 13, 1636–1642. [Google Scholar] [CrossRef]
- Hofmann, H.; Heinz, F.X.; Dippe, H. ELISA for IgM and IgG antibodies against tick-borne encephalitis virus: Quantification and standardization of results. Zent. Bakteriol. Mikrobiol. Hyg. A 1983, 255, 448–455. [Google Scholar] [CrossRef]
- Wilder-Smith, A.; Ooi, E.-E.; Horstick, O.; Wills, B. Dengue. Lancet 2019, 393, 350–363. [Google Scholar] [CrossRef]
- Bogovič, P.; Lotrič-Furlan, S.; Avšič-Županc, T.; Lusa, L.; Strle, F. Factors associated with severity of tick-borne encephalitis: A prospective observational study. Travel Med. Infect. Dis. 2018, 26, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Lenhard, T.; Ott, D.; Jakob, N.J.; Martinez-Torres, F.; Grond-Ginsbach, C.; Meyding-Lamadé, U. Clinical outcome and cerebrospinal fluid profiles in patients with tick-borne encephalitis and prior vaccination history. Ticks Tick-Borne Dis. 2018, 9, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Hansson, K.E.; Rosdahl, A.; Insulander, M.; Vene, S.; Lindquist, L.; Gredmark-Russ, S.; Askling, H.H. Tick-borne Encephalitis Vaccine Failures: A 10-year Retrospective Study Supporting the Rationale for Adding an Extra Priming Dose in Individuals Starting at Age 50 Years. Clin. Infect. Dis. 2019, 70, 245–251. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| TBE Patients | No. of Cases | Age (Years) | Sex (f/m) | |
|---|---|---|---|---|
| Median | Range | |||
| Unvaccinated | 18 | 57 | 6–74 | 8/10 |
| Vaccination breakthroughs | 18 (4 a, 14 b) | 52 | 5–75 | 6/12 |
| TBE Patients | Days between Serum Samples | ||
|---|---|---|---|
| n | Median (Days) | Range (Days) | |
| 1st to 2nd samples (d5–19) | |||
| Unvaccinated | 12 | 14 | 6–17 |
| Vaccination breakthroughs | 15 | 12 | 5–18 |
| 1st to 3rd samples (d20–45) | |||
| Unvaccinated | 8 | 25 | 20–45 |
| Vaccination breakthroughs | 6 | 28 | 21–40 |
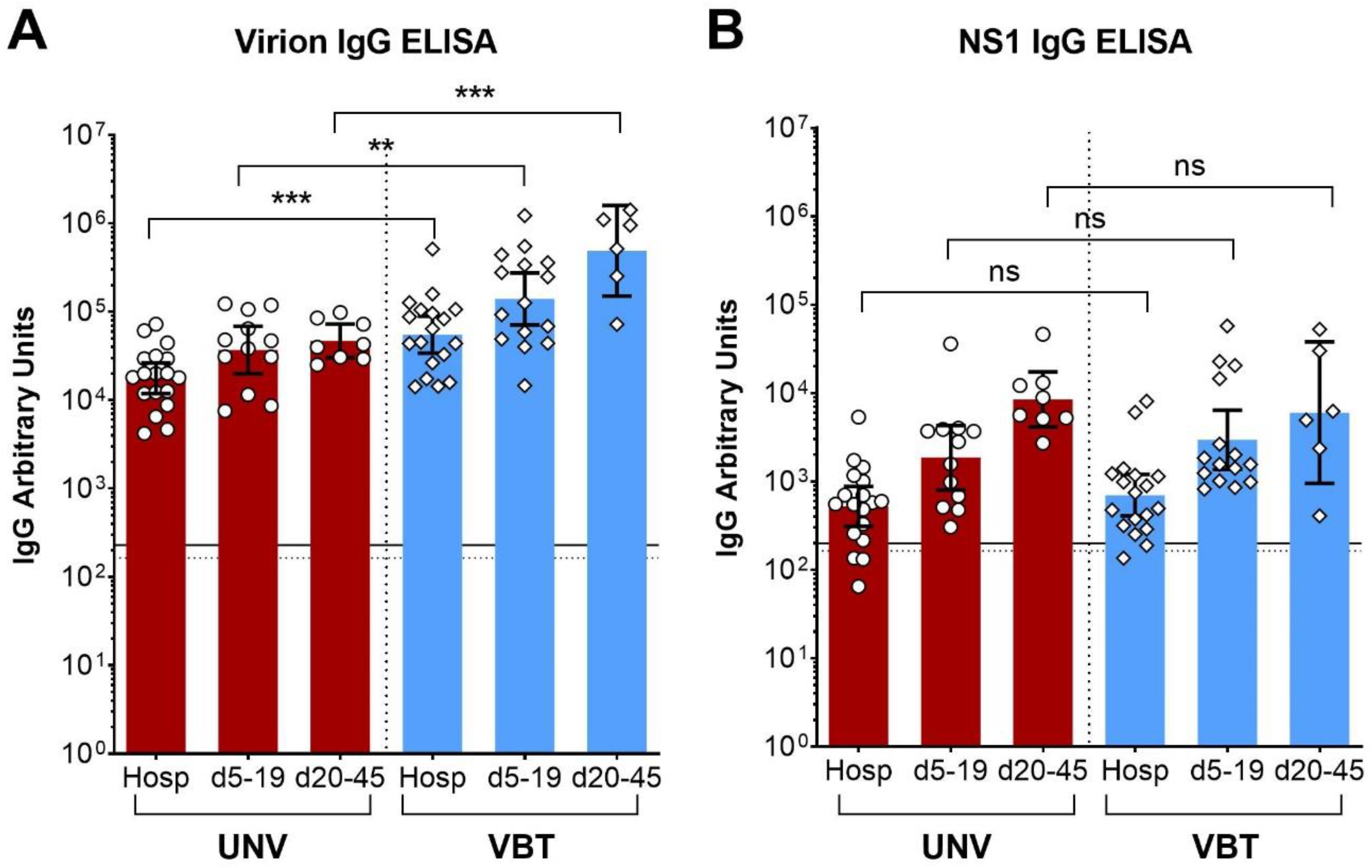
| TBE Patients | Virion IgG ELISA | ||||
| Arbitrary Units (Geometric Mean) | p Values ANOVA (Dunnett’s MCT a) | ||||
| Hosp b | d5–19 | d20–45 | Hosp vs. d5–19 | Hosp vs. d20–45 | |
| Unvaccinated | 17,705 | 36,844 | 46,866 | 0.041 | 0.016 |
| Vaccination breakthroughs | 55,105 | 139,477 | 490,156 | 0.039 | 0.0003 |
| TBE Patients | NS1 IgG ELISA | ||||
| Arbitrary units (geometric mean) | p values ANOVA (Dunnett’s MCT a) | ||||
| Hosp | d5–19 | d20–45 | Hosp vs. d5–19 | Hosp vs. d20–45 | |
| Unvaccinated | 523 | 1860 | 8485 | 0.008 | <0.0001 |
| Vaccination breakthroughs | 700 | 2954 | 6008 | 0.007 | 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stiasny, K.; Leitner, A.; Holzmann, H.; Heinz, F.X. Dynamics and Extent of Non-Structural Protein 1-Antibody Responses in Tick-Borne Encephalitis Vaccination Breakthroughs and Unvaccinated Patients. Viruses 2021, 13, 1007. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061007
Stiasny K, Leitner A, Holzmann H, Heinz FX. Dynamics and Extent of Non-Structural Protein 1-Antibody Responses in Tick-Borne Encephalitis Vaccination Breakthroughs and Unvaccinated Patients. Viruses. 2021; 13(6):1007. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061007
Chicago/Turabian StyleStiasny, Karin, Agnes Leitner, Heidemarie Holzmann, and Franz X. Heinz. 2021. "Dynamics and Extent of Non-Structural Protein 1-Antibody Responses in Tick-Borne Encephalitis Vaccination Breakthroughs and Unvaccinated Patients" Viruses 13, no. 6: 1007. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061007