
Differential Regulation of Cellular FAM111B by Human Adenovirus C Type 5 E1 Oncogenes

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Culture Conditions
2.2. Plasmids and Transient Transfections
2.3. Luciferase Reporter Assays
2.4. Viruses
2.5. Antibodies and Protein Analysis
2.6. In Vitro Translation and Pulldown Assays
2.7. Indirect Immunofluorescence
2.8. Isolation and Quantification of Nucleic Acids
3. Results and Discussion
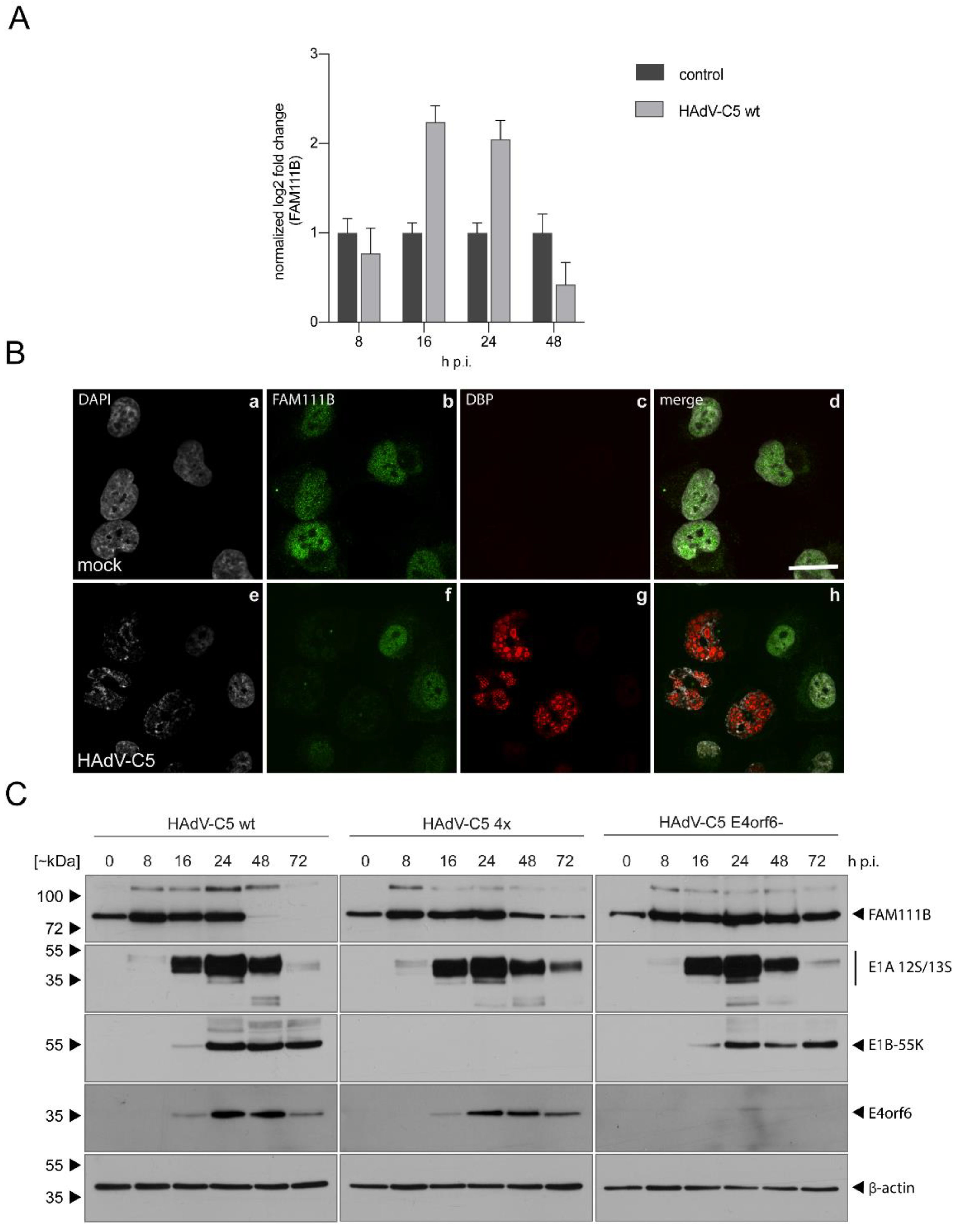
3.1. FAM111B RNA and Protein Levels Are Upregulated in HAdV-C5-Infections
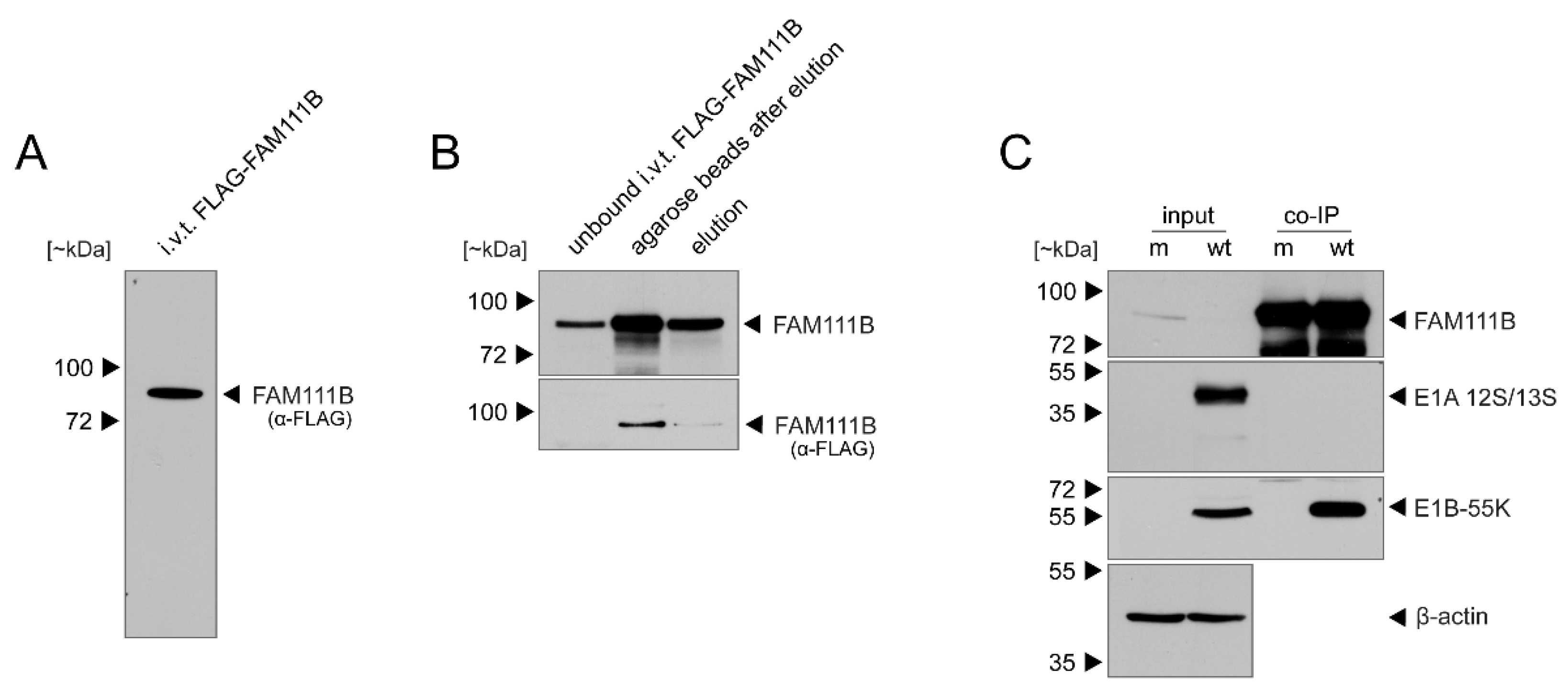
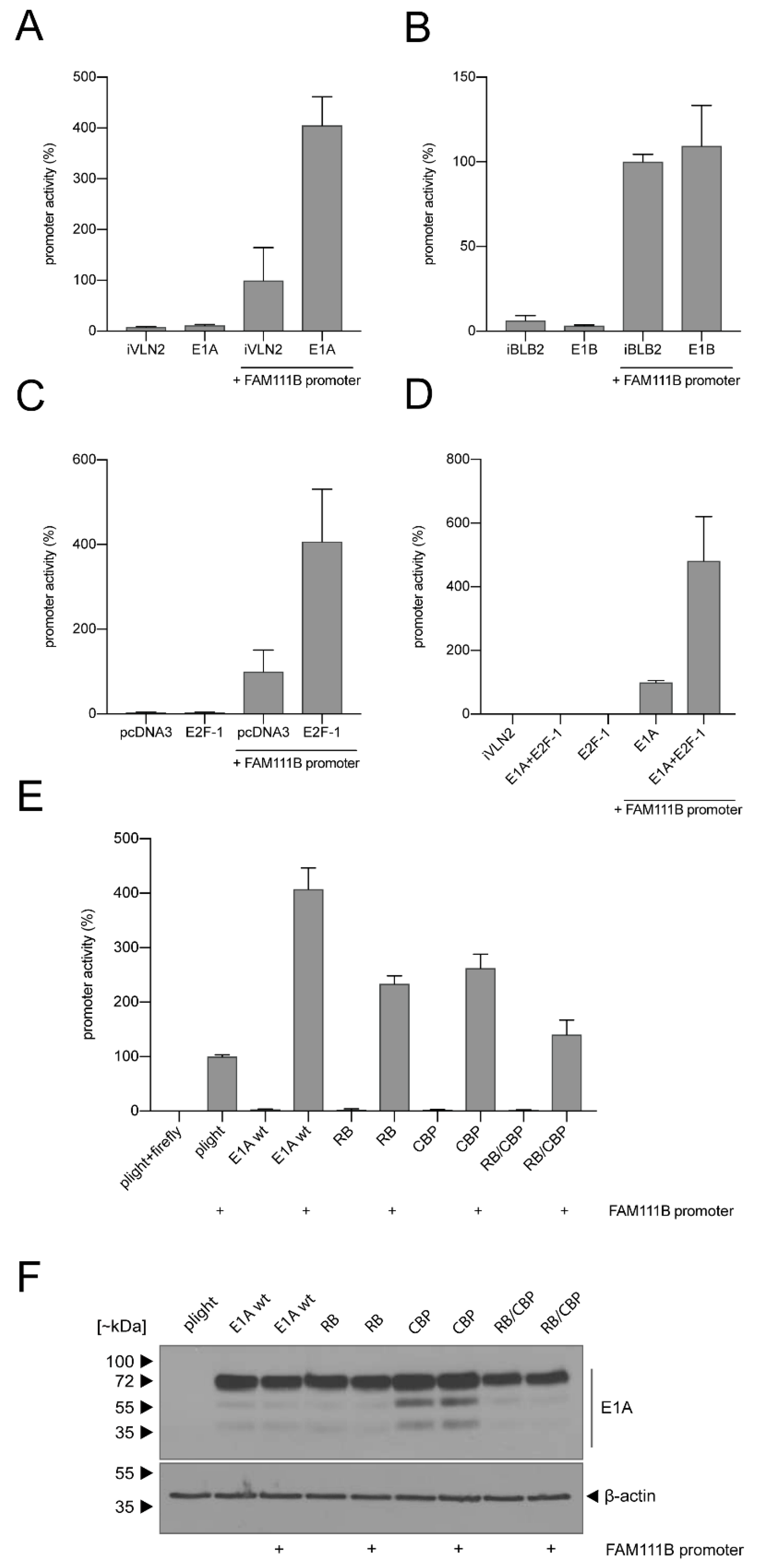
3.2. FAM111B Expression Levels Are Differentially Regulated by E1A, E1B-55K and E4orf6
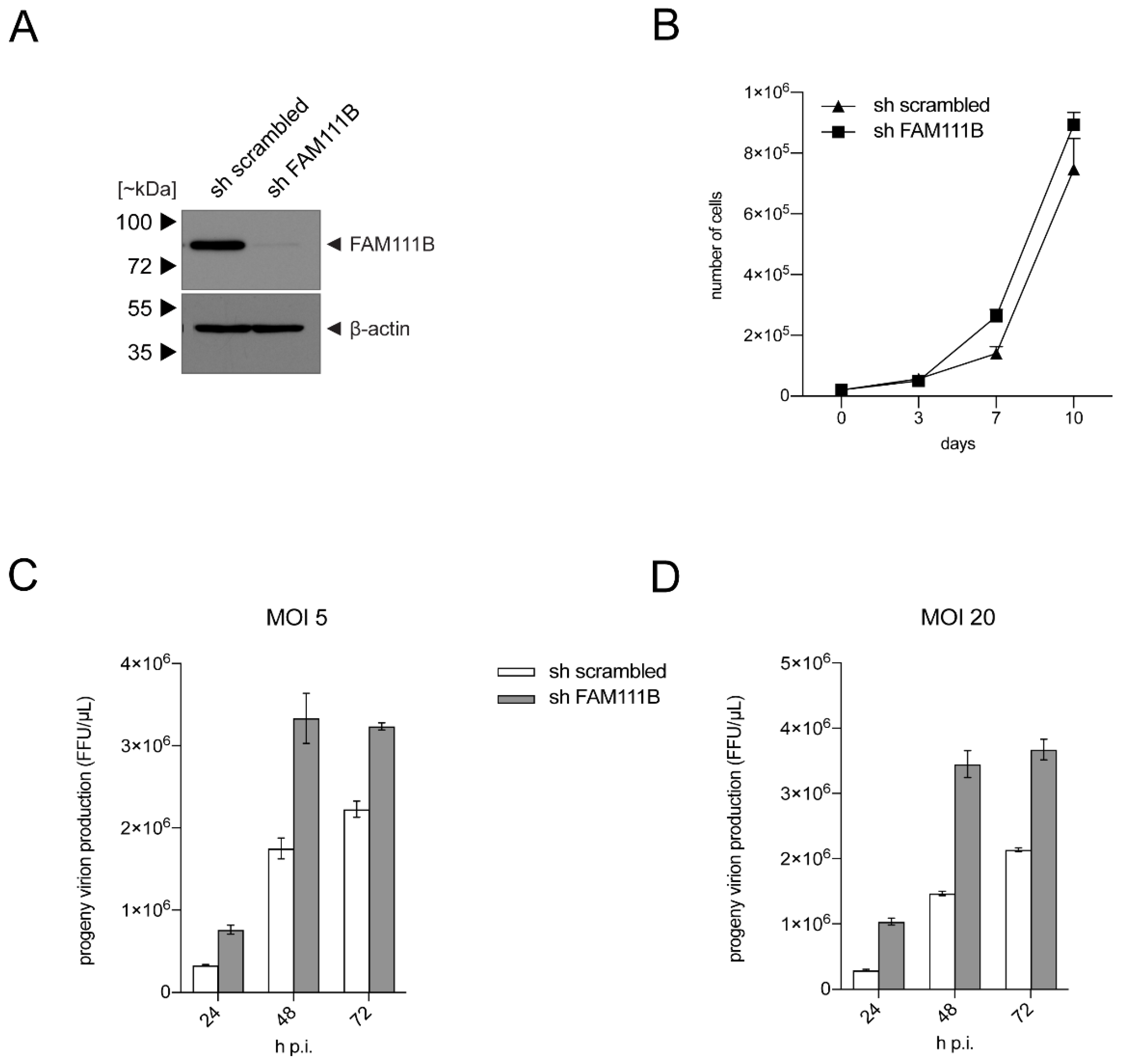
3.3. FAM111B Knockdown Increases HAdV-C5 Gene Expression
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lion, T. Adenovirus infections in immunocompetent and immunocompromised patients. Clin. Microbiol. Rev. 2014, 27, 441–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, G.; Flint, S.J. Normal human cell proteins that interact with the adenovirus type 5 E1B 55kDa protein. Virology 2017, 504, 12–24. [Google Scholar] [CrossRef]
- Mercier, S.; Kury, S.; Shaboodien, G.; Houniet, D.T.; Khumalo, N.P.; Bou-Hanna, C.; Bodak, N.; Cormier-Daire, V.; David, A.; Faivre, L.; et al. Mutations in FAM111B cause hereditary fibrosing poikiloderma with tendon contracture, myopathy, and pulmonary fibrosis. Am. J. Hum. Genet. 2013, 93, 1100–1107. [Google Scholar] [CrossRef]
- Abraham, M.B.; Li, D.; Tang, D.; O’Connell, S.M.; McKenzie, F.; Lim, E.M.; Hakonarson, H.; Levine, M.A.; Choong, C.S. Short stature and hypoparathyroidism in a child with Kenny-Caffey syndrome type 2 due to a novel mutation in FAM111A gene. Int. J. Pediatr. Endocrinol. 2017, 2017, 1. [Google Scholar] [CrossRef] [Green Version]
- Nikkel, S.M.; Ahmed, A.; Smith, A.; Marcadier, J.; Bulman, D.E.; Boycott, K.M. Mother-to-daughter transmission of Kenny-Caffey syndrome associated with the recurrent, dominant FAM111A mutation p.Arg569His. Clin. Genet. 2014, 86, 394–395. [Google Scholar] [CrossRef] [PubMed]
- Akamatsu, S.; Takata, R.; Haiman, C.A.; Takahashi, A.; Inoue, T.; Kubo, M.; Furihata, M.; Kamatani, N.; Inazawa, J.; Chen, G.K.; et al. Common variants at 11q12, 10q26 and 3p11.2 are associated with prostate cancer susceptibility in Japanese. Nat. Genet. 2012, 44, 426–429. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, K.; Nojima, S.; Hijiki, S.; Tahara, S.; Ohshima, K.; Matsui, T.; Hori, Y.; Kurashige, M.; Umeda, D.; Kiyokawa, H.; et al. FAM111B enhances proliferation of KRAS-driven lung adenocarcinoma by degrading p16. Cancer Sci. 2020, 111, 2635–2646. [Google Scholar] [CrossRef]
- Sun, H.; Liu, K.; Huang, J.; Sun, Q.; Shao, C.; Luo, J.; Xu, L.; Shen, Y.; Ren, B. FAM111B, a direct target of p53, promotes the malignant process of lung adenocarcinoma. Onco Targets Ther. 2019, 12, 2829–2842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, W.; Wu, X.; Starnes, S.; Johnson, S.K.; Haessler, J.; Wang, S.; Chen, L.; Barlogie, B.; Shaughnessy, J.D., Jr.; Zhan, F. An analysis of the clinical and biologic significance of TP53 loss and the identification of potential novel transcriptional targets of TP53 in multiple myeloma. Blood 2008, 112, 4235–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fine, D.A.; Rozenblatt-Rosen, O.; Padi, M.; Korkhin, A.; James, R.L.; Adelmant, G.; Yoon, R.; Guo, L.; Berrios, C.; Zhang, Y.; et al. Identification of FAM111A as an SV40 host range restriction and adenovirus helper factor. PLoS Pathog. 2012, 8, e1002949. [Google Scholar] [CrossRef] [PubMed]
- Nie, M.; Oravcová, M.; Jami-Alahmadi, Y.; Wohlschlegel, J.A.; Lazzerini-Denchi, E.; Boddy, M.N. Activation of FAM111A Protease Induces Defects in Nuclear Function that Likely Underlie its Roles in Disease and Viral Restriction. bioRxiv 2020. [Google Scholar] [CrossRef]
- Panda, D.; Fernandez, D.J.; Lal, M.; Buehler, E.; Moss, B. Triad of human cellular proteins, IRF2, FAM111A, and RFC3, restrict replication of orthopoxvirus SPI-1 host-range mutants. Proc. Natl. Acad. Sci. USA 2017, 114, 3720–3725. [Google Scholar] [CrossRef] [Green Version]
- Tarnita, R.M.; Wilkie, A.R.; DeCaprio, J.A. Contribution of DNA Replication to the FAM111A-Mediated Simian Virus 40 Host Range Phenotype. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Schreiner, S.; Wimmer, P.; Sirma, H.; Everett, R.D.; Blanchette, P.; Groitl, P.; Dobner, T. Proteasome-dependent degradation of Daxx by the viral E1B-55K protein in human adenovirus-infected cells. J. Virol. 2010, 84, 7029–7038. [Google Scholar] [CrossRef] [Green Version]
- Speiseder, T.; Hofmann-Sieber, H.; Rodriguez, E.; Schellenberg, A.; Akyuz, N.; Dierlamm, J.; Spruss, T.; Lange, C.; Dobner, T. Efficient Transformation of Primary Human Mesenchymal Stromal Cells by Adenovirus Early Region 1 Oncogenes. J. Virol. 2017, 91, e01782-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groitl, P.; Dobner, T. Construction of adenovirus type 5 early region 1 and 4 virus mutants. Methods Mol. Med. 2007, 130, 29–39. [Google Scholar] [CrossRef]
- Kindsmüller, K.; Groitl, P.; Härtl, B.; Blanchette, P.; Hauber, J.; Dobner, T. Intranuclear targeting and nuclear export of the adenovirus E1B-55K protein are regulated by SUMO1 conjugation. Proc. Natl. Acad. Sci. USA 2007, 104, 6684–6689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchette, P.; Kindsmüller, K.; Groitl, P.; Dallaire, F.; Speiseder, T.; Branton, P.E.; Dobner, T. Control of mRNA export by adenovirus E4orf6 and E1B55K proteins during productive infection requires E4orf6 ubiquitin ligase activity. J. Virol. 2008, 82, 2642–2651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kindsmüller, K.; Schreiner, S.; Leinenkugel, F.; Groitl, P.; Kremmer, E.; Dobner, T. A 49-kilodalton isoform of the adenovirus type 5 early region 1B 55-kilodalton protein is sufficient to support virus replication. J. Virol. 2009, 83, 9045–9056. [Google Scholar] [CrossRef] [Green Version]
- Bürck, C.; Mund, A.; Berscheminski, J.; Kieweg, L.; Müncheberg, S.; Dobner, T.; Schreiner, S. KAP1 Is a Host Restriction Factor That Promotes Human Adenovirus E1B-55K SUMO Modification. J. Virol. 2016, 90, 930–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freudenberger, N.; Meyer, T.; Groitl, P.; Dobner, T.; Schreiner, S. Human Adenovirus Core Protein V Is Targeted by the Host SUMOylation Machinery To Limit Essential Viral Functions. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Puvion-Dutilleul, F.; Pedron, J.; Cajean-Feroldi, C. Identification of intranuclear structures containing the 72K DNA-binding protein of human adenovirus type 5. Eur. J. Cell Biol. 1984, 34, 313–322. [Google Scholar]
- Cheng, C.Y.; Blanchette, P.; Branton, P.E. The adenovirus E4orf6 E3 ubiquitin ligase complex assembles in a novel fashion. Virology 2007, 364, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Schreiner, S.; Wimmer, P.; Dobner, T. Adenovirus degradation of cellular proteins. Future Microbiol. 2012, 7, 211–225. [Google Scholar] [CrossRef]
- Blanchette, P.; Cheng, C.Y.; Yan, Q.; Ketner, G.; Ornelles, D.A.; Dobner, T.; Conaway, R.C.; Conaway, J.W.; Branton, P.E. Both BC-box motifs of adenovirus protein E4orf6 are required to efficiently assemble an E3 ligase complex that degrades p53. Mol. Cell. Biol. 2004, 24, 9619–9629. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.Y.; Gilson, T.; Wimmer, P.; Schreiner, S.; Ketner, G.; Dobner, T.; Branton, P.E.; Blanchette, P. Role of E1B55K in E4orf6/E1B55K E3 ligase complexes formed by different human adenovirus serotypes. J. Virol. 2013, 87, 6232–6245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Querido, E.; Blanchette, P.; Yan, Q.; Kamura, T.; Morrison, M.; Boivin, D.; Kaelin, W.G.; Conaway, R.C.; Conaway, J.W.; Branton, P.E. Degradation of p53 by adenovirus E4orf6 and E1B55K proteins occurs via a novel mechanism involving a Cullin-containing complex. Genes Dev. 2001, 15, 3104–3117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelka, P.; Miller, M.S.; Cecchini, M.; Yousef, A.F.; Bowdish, D.M.; Dick, F.; Whyte, P.; Mymryk, J.S. Adenovirus E1A directly targets the E2F/DP-1 complex. J. Virol. 2011, 85, 8841–8851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelka, P.; Ablack, J.N.; Torchia, J.; Turnell, A.S.; Grand, R.J.; Mymryk, J.S. Transcriptional control by adenovirus E1A conserved region 3 via p300/CBP. Nucleic Acids Res. 2009, 37, 1095–1106. [Google Scholar] [CrossRef] [Green Version]
- Rouillard, A.D.; Gundersen, G.W.; Fernandez, N.F.; Wang, Z.; Monteiro, C.D.; McDermott, M.G.; Ma’ayan, A. The harmonizome: A collection of processed datasets gathered to serve and mine knowledge about genes and proteins. Database 2016, 2016. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ip, W.-H.; Wilkens, B.; Solomatina, A.; Martin, J.; Melling, M.; Hidalgo, P.; Bertzbach, L.D.; Speiseder, T.; Dobner, T. Differential Regulation of Cellular FAM111B by Human Adenovirus C Type 5 E1 Oncogenes. Viruses 2021, 13, 1015. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061015
Ip W-H, Wilkens B, Solomatina A, Martin J, Melling M, Hidalgo P, Bertzbach LD, Speiseder T, Dobner T. Differential Regulation of Cellular FAM111B by Human Adenovirus C Type 5 E1 Oncogenes. Viruses. 2021; 13(6):1015. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061015
Chicago/Turabian StyleIp, Wing-Hang, Britta Wilkens, Anastasia Solomatina, Judith Martin, Michael Melling, Paloma Hidalgo, Luca D. Bertzbach, Thomas Speiseder, and Thomas Dobner. 2021. "Differential Regulation of Cellular FAM111B by Human Adenovirus C Type 5 E1 Oncogenes" Viruses 13, no. 6: 1015. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061015