
COVID-19 Severity Potentially Modulated by Cardiovascular-Disease-Associated Immune Dysregulation


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Downloading Data
2.2. Differential Expression
2.3. GSEA
2.4. CIBERSORTx
3. Results
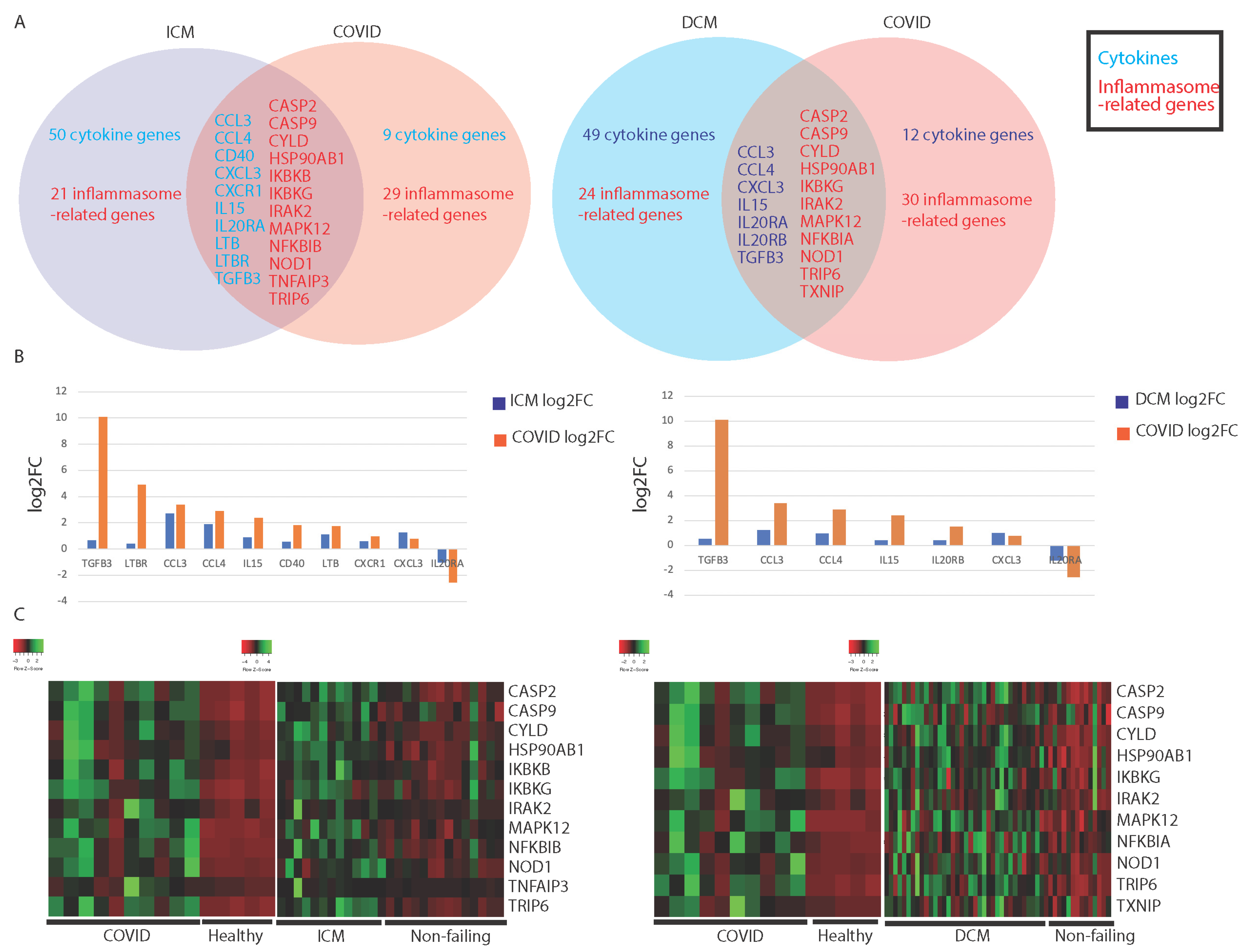
3.1. Comparing Immune Profiles of COVID-19 and Cardiomyopathy Patients
3.1.1. Similarities in Immune-Associated Gene Dysregulation in COVID-19 and Cardiomyopathy
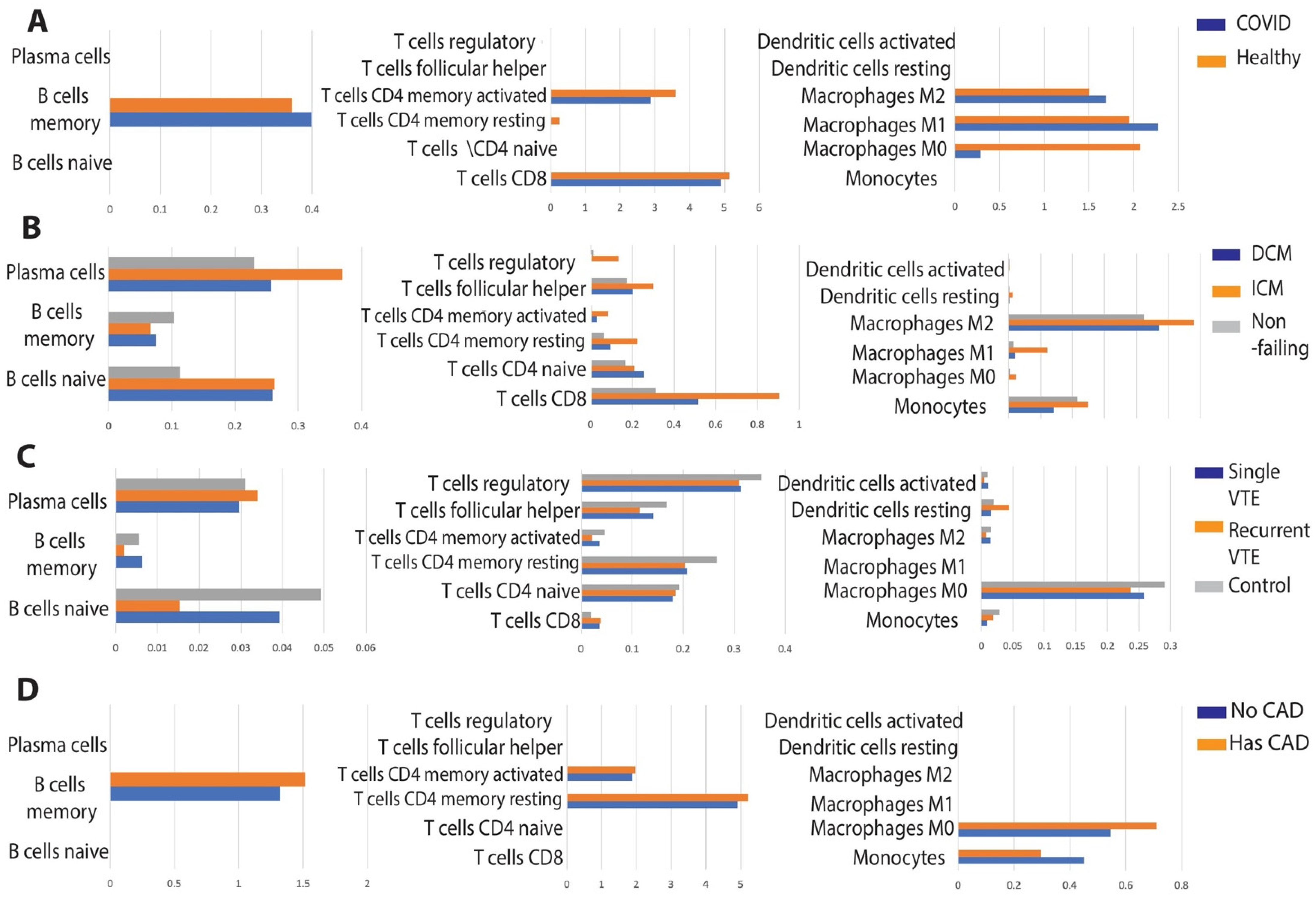
3.1.2. Comparison of Immune Cell Population Abundance in COVID-19 vs. Cardiomyopathy
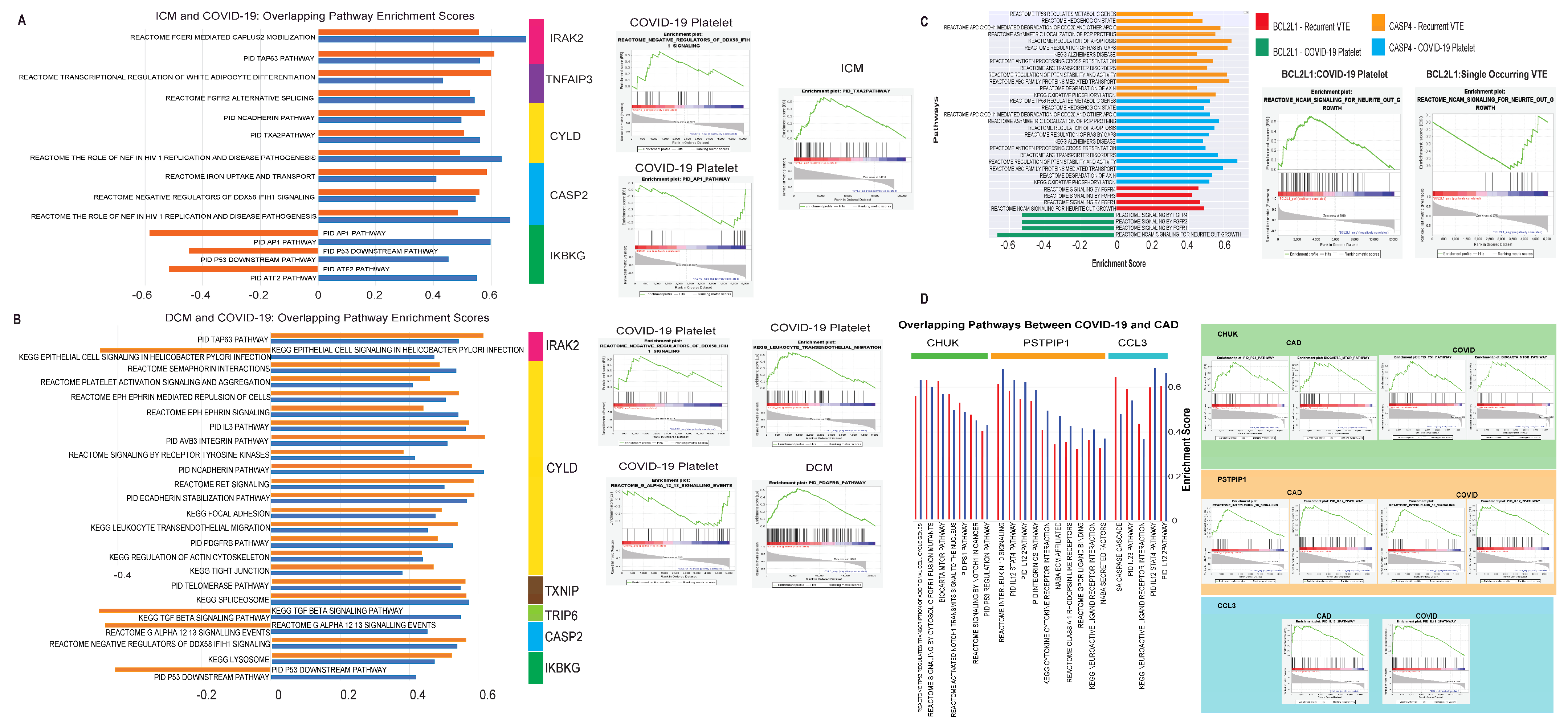
3.1.3. Evaluation of Canonical Pathways Correlated with Genes Dysregulated in Both COVID-19 and Cardiomyopathy
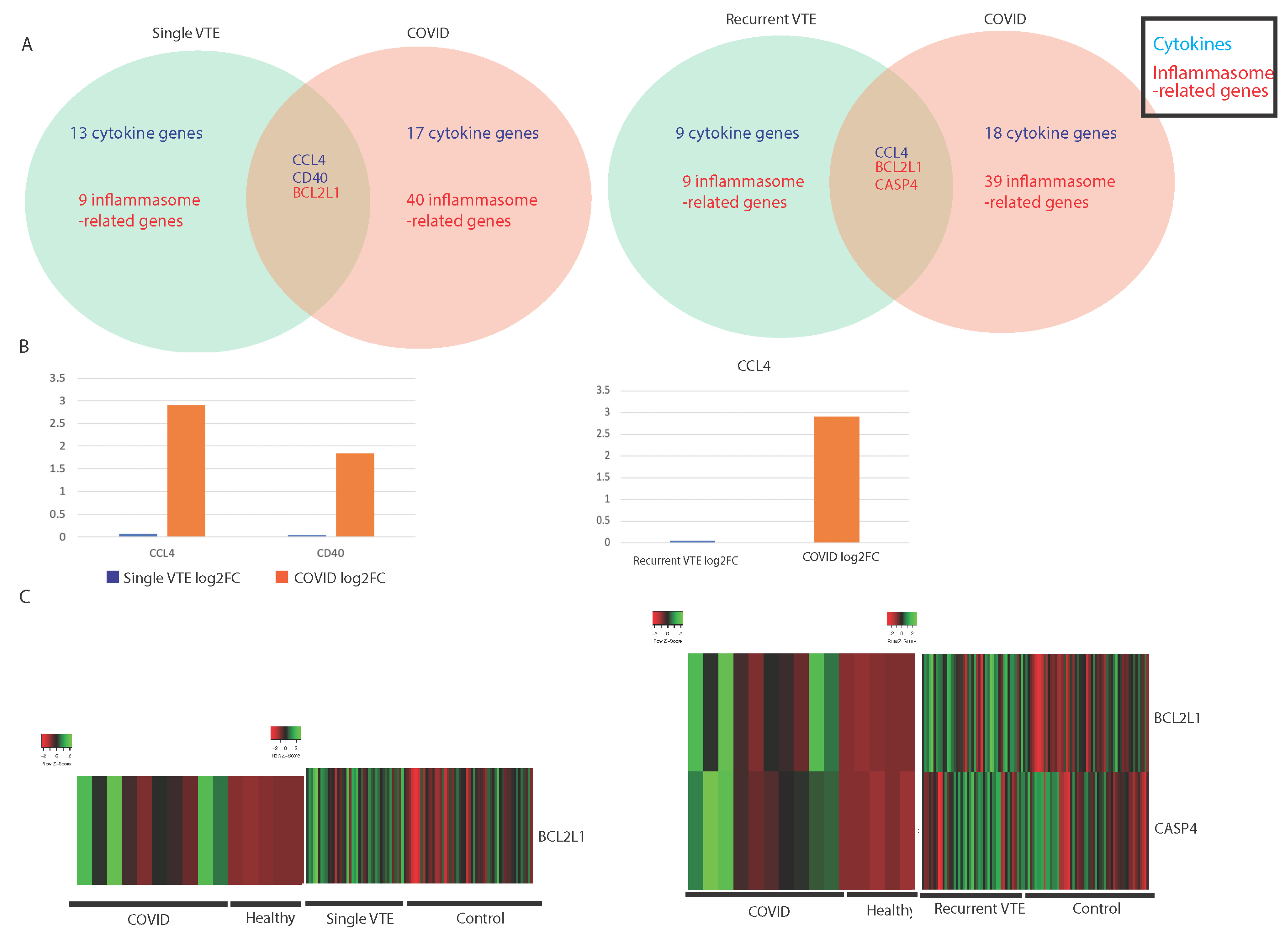
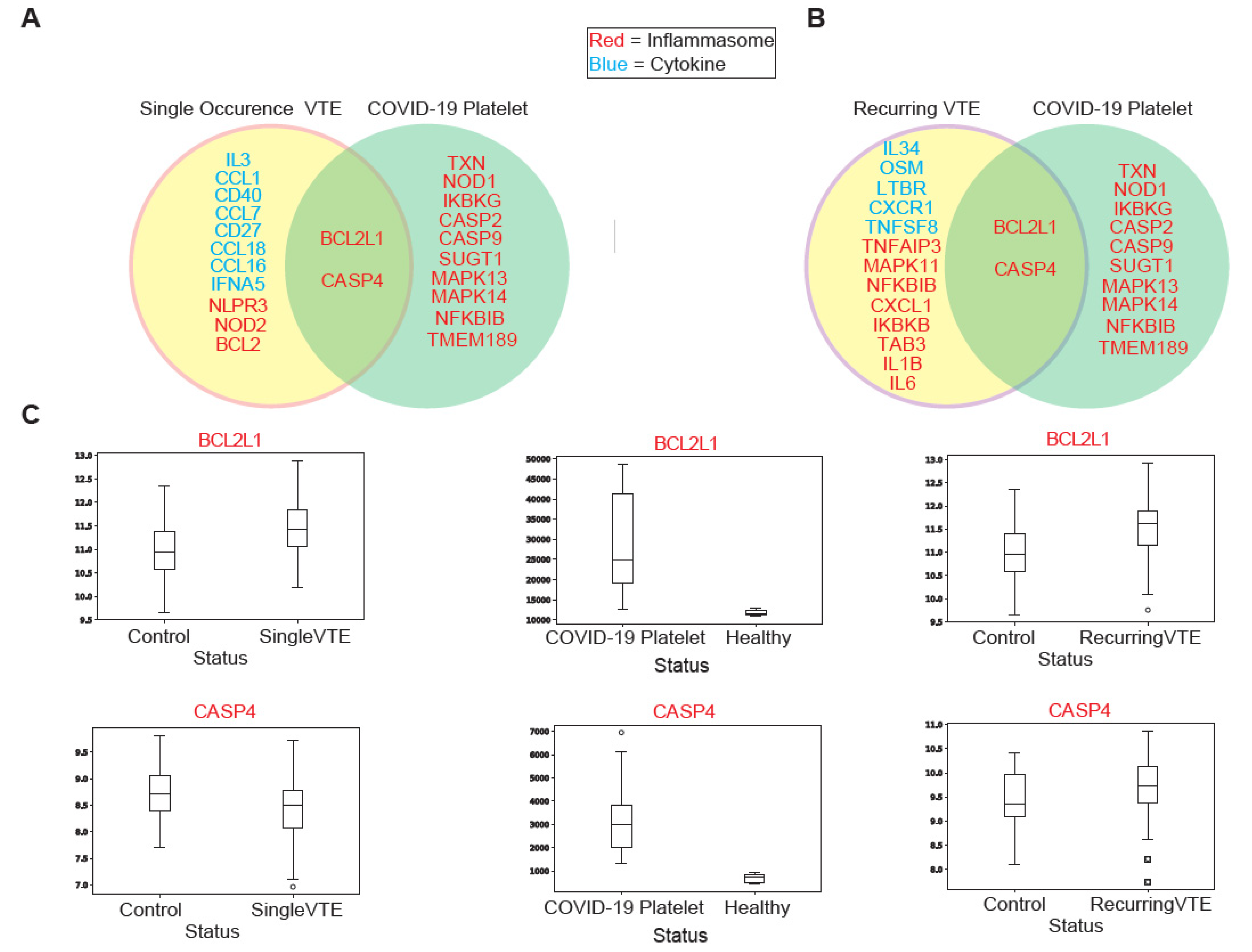
3.2. Comparing Immune Profiles of COVID-19 and VTE Patients
3.2.1. Similarities in Immune-Associated Gene Dysregulation in COVID-19 and VTE
3.2.2. Comparison of Immune Cell Population Abundance in COVID-19 vs. VTE
3.2.3. Evaluation of Canonical Pathways Correlated with Genes Dysregulated in COVID-19 and VTE
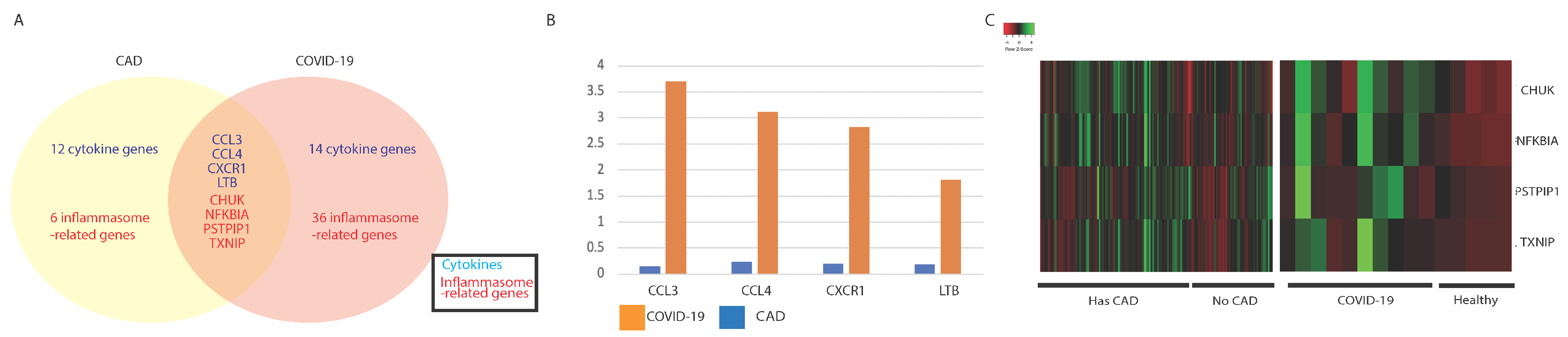
3.3. Comparing Immune Profiles of COVID-19 and CAD Patients
3.3.1. Similarities in Immune-Associated Gene Dysregulation in COVID-19 and CAD
3.3.2. Comparison of Immune Cell Population Abundance in COVID-19 vs. CAD
3.3.3. Evaluation of Pathways Correlated with Genes Dysregulated in COVID-19 and CAD
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ICM Only | ICM and COVID-19 | COVID-19 Only | DCM Only | DCM and COVID-19 | COVID-19 Only | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CCL17 | CXCR4 | TNF | CCL3 | CCL2 | MAPK3 | CCL11 | CXCR4 | TNF | CCL3 | CD40 | MAPK8 |
| CCL21 | IFNA14 | TNFSF12 | CCL4 | CKLF | MAPK8 | CCL17 | EPO | TNFSF11 | CCL4 | CXCL5 | NAIP |
| CCL22 | IL11 | TNFSF13 | CD40 | CXCL5 | MAPK9 | CCL2 | IFNA14 | TNFSF12 | CXCL3 | CXCR1 | NFKBIB |
| CCL24 | IL11RA | TNFSF13B | CXCL3 | EPOR | NAIP | CCL22 | IL10 | TNFSF13B | IL15 | EPOR | POLR2J4 |
| CCL5 | IL12A | TNFSF8 | CXCR1 | IL15RA | NFKBIA | CCL24 | IL11 | TNFSF14 | IL20RA | IL1RN | RELA |
| CCL8 | IL16 | TNFSF9 | IL15 | IL1RN | POLR2J4 | CCL5 | IL11RA | BIRC2 | IL20RB | LTB | SUGT1 |
| CCR10 | IL17B | APP | IL20RA | IL20RB | PSTPIP1 | CCL8 | IL12A | BIRC3 | TGFB3 | LTBR | TAB1 |
| CCR3 | IL17C | BIRC2 | LTB | TGFB1 | RELA | CCR1 | IL13 | CARD8 | CASP2 | TGFB1 | TAB3 |
| CCR4 | IL17D | CARD8 | LTBR | TGFBR2 | SUGT1 | CCR10 | IL15RA | CCL11 | CASP9 | CCL2 | TNFAIP3 |
| CCR7 | IL1A | CCL5 | TGFB3 | BCL2L1 | TAB1 | CCR3 | IL16 | CCL2 | CYLD | CKLF | TRAF6 |
| CD27 | IL1B | CCL8 | CASP2 | BIRC3 | TAB3 | CCR4 | IL17B | CCL5 | HSP90AB1 | IL15RA | UBE2N |
| CD4 | IL2 | CXCL1 | CASP9 | CARD6 | TMEM189 | CCR7 | IL17D | CCL8 | IKBKG | TGFBR2 | BIRC3 |
| CKLF | IL23A | CXCL2 | CYLD | CASP4 | TRAF6 | CD27 | IL18 | CXCL2 | IRAK2 | BCL2L1 | HSP90AA1 |
| CX3CL1 | IL25 | HSP90AA1 | HSP90AB1 | CASP8 | TXN | CD4 | IL20 | HSP90AA1 | MAPK12 | CARD6 | IRAK1 |
| CXCL1 | IL27 | IL1B | IKBKB | CHUK | TXNIP | CKLF | IL23A | IL18 | NFKBIA | CASP4 | MAPK1 |
| CXCL10 | IL33 | IL6 | IKBKG | HSP90AA1 | UBE2N | CX3CL1 | IL25 | IRAK1 | NOD1 | CASP8 | MAPK10 |
| CXCL14 | IL34 | MAPK1 | IRAK2 | HSP90B1 | CXCL14 | IL33 | MAP3K7 | TRIP6 | CHUK | MAPK9 | |
| CXCL16 | IL6 | MAPK11 | MAPK12 | IRAK1 | CXCL16 | OSMR | MAPK1 | TXNIP | HSP90B1 | PSTPIP1 | |
| CXCL2 | IL9R | MAPK9 | NFKBIB | MAPK1 | CXCL2 | TGFB2 | MAPK10 | IKBKB | TMEM189 | ||
| CXCL9 | OSM | NFKB1 | NOD1 | MAPK10 | CXCL9 | TGFBR1 | MAPK9 | MAPK13 | TXN | ||
| CXCR2 | TGFB2 | NLRC4 | TNFAIP3 | MAPK13 | CXCR2 | TGFBR2 | NLRC4 | MAPK14 | |||
| CXCR3 | TGFBR1 | NLRP3 | TRIP6 | MAPK14 | CXCR3 | TGFBR3 | NLRP1 | MAPK3 | |||
| TAB2 | NLRP3 | ||||||||||
| TAB3 | PSTPIP1 | ||||||||||
| TNF | PYCARD | ||||||||||
| TRAF6 | TAB2 | ||||||||||
| XIAP | TMEM189 | ||||||||||
| - | - | - | - | - | - | - | - | TNF | - | - | - |
| TXN | |||||||||||
| Single VTE Only | Single VTE and COVID-19 | COVID-19 Only | Recurrent VTE Only | Recurrent VTE and COVID-19 | COVID-19 Only | ||||
|---|---|---|---|---|---|---|---|---|---|
| CCL1 | CCL4 | CCL2 | MAPK8 | IKBKB | CD27 | CCL4 | CCL2 | MAPK3 | IKBKB |
| CCL16 | CD40 | CCL3 | NAIP | IKBKG | CXCL10 | BCL2L1 | CCL3 | MAPK8 | IKBKG |
| CCL18 | BCL2L1 | CKLF | NFKBIA | IRAK1 | CXCL8 | CASP4 | CD40 | NAIP | IRAK1 |
| CCL7 | CXCL3 | NFKBIB | IRAK2 | IFNG | CKLF | NFKBIA | IRAK2 | ||
| CD27 | CXCL5 | NOD1 | TAB1 | IL13 | CXCL3 | NFKBIB | TAB1 | ||
| CXCL10 | CXCR1 | POLR2J4 | TAB3 | IL16 | CXCL5 | NOD1 | TAB3 | ||
| CXCL8 | EPOR | PSTPIP1 | TMEM189 | IL1A | CXCR1 | POLR2J4 | TMEM189 | ||
| IL13 | IL15 | SUGT1 | TNFAIP3 | IL3 | EPOR | PSTPIP1 | TNFAIP3 | ||
| IL16 | IL15RA | CASP4 | TRAF6 | TNFSF10 | IL15 | SUGT1 | TRAF6 | ||
| IL1A | IL1RN | MAPK1 | TRIP6 | MAPK1 | IL15RA | MAPK1 | TRIP6 | ||
| IL3 | IL20RA | MAPK9 | TXN | MAPK9 | IL1RN | MAPK9 | TXN | ||
| TNFSF10 | IL20RB | RELA | TXNIP | RELA | IL20RA | RELA | TXNIP | ||
| IFNA5 | LTB | BIRC3 | UBE2N | BIRC2 | IL20RB | BIRC3 | UBE2N | ||
| BCL2 | LTBR | CARD6 | CARD8 | LTB | CARD6 | ||||
| CASP5 | TGFB1 | CASP2 | CASP5 | LTBR | CASP2 | ||||
| IL18 | TGFB3 | CASP8 | IL18 | TGFB1 | CASP8 | ||||
| NLRP3 | TGFBR2 | CASP9 | IL1B | TGFB3 | CASP9 | ||||
| NOD2 | MAPK10 | CHUK | NLRP3 | TGFBR2 | CHUK | ||||
| CASP4 | MAPK12 | CYLD | MAPK10 | CYLD | |||||
| MAPK1 | MAPK13 | HSP90AA1 | MAPK12 | HSP90AA1 | |||||
| MAPK9 | MAPK14 | HSP90AB1 | MAPK13 | HSP90AB1 | |||||
| RELA | MAPK3 | HSP90B1 | MAPK14 | HSP90B1 | |||||
| CAD Only | CAD and COVID-19 | COVID-19 Only | ||
|---|---|---|---|---|
| CCR4 | CCL3 | CCL2 | CASP4 | MAPK9NAIP |
| CD27 | CCL4 | CKLF | CASP8 | NFKBIB |
| CD40 | CXCR1 | CXCL3 | CASP9 | NOD1 |
| CXCR3 | LTB | CXCL5 | CYLD | POLR2J4 |
| CXCR4 | CHUK | EPOR | HSP90AA1 | RELA |
| IL17B | NFKBIA | IL15 | HSP90AB1 | SUGT1 |
| IL21 | PSTPIP1 | IL15RA | HSP90B1 | TAB1 |
| IL6 | TXNIP | IL1RN | IKBKB | TAB3 |
| OSM | IL20RA | IKBKG | TMEM189 | |
| TNFSF10 | IL20RB | IRAK1 | TNFAIP3 | |
| TNFSF13B | LTBR | IRAK2 | TRAF6 | |
| TNFSF14 | TGFB1 | MAPK1 | TRIP6 | |
| CASP1 | TGFB3 | MAPK10 | TXN | |
| CASP5 | TGFBR2 | MAPK12 | UBE2N | |
| MAPK11 | BCL2L1 | MAPK13 | ||
| NLRP3 | BIRC3 | MAPK14 | ||
| PYCARD | CARD6 | MAPK3 | ||
| TAB2 | CASP2 | MAPK8 | ||
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Tsitsiou, E.; Lindsay, M.A. microRNAs and the immune response. Curr. Opin. Pharmacol. 2009, 9, 514–520. [Google Scholar] [CrossRef]
- Yang, J.; Zheng, Y.; Gou, X.; Pu, K.; Chen, Z.; Guo, Q.; Ji, R.; Wang, H.; Wang, Y.; Zhou, Y. Prevalence of comorbidities and its effects in patients infected with SARS-CoV-2: A systematic review and meta-analysis. Int. J. Infect. Dis. 2020, 94, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Pranata, R.; Huang, I.; Lim, M.A.; Wahjoepramono, E.J.; July, J. Impact of cerebrovascular and cardiovascular diseases on mortality and severity of COVID-19–systematic review, meta-analysis, and meta-regression. J. Stroke Cerebrovasc. Dis. 2020, 29, 104949. [Google Scholar] [CrossRef]
- Tian, W.; Jiang, W.; Yao, J.; Nicholson, C.J.; Li, R.H.; Sigurslid, H.H.; Wooster, L.; Rotter, J.I.; Guo, X.; Malhotra, R. Predictors of mortality in hospitalized COVID-19 patients: A systematic review and meta-analysis. J. Med. Virol. 2020, 92, 1875–1883. [Google Scholar] [CrossRef]
- Shi, S.; Qin, M.; Shen, B.; Cai, Y.; Liu, T.; Yang, F.; Gong, W.; Liu, X.; Liang, J.; Zhao, Q.; et al. Association of Cardiac Injury with Mortality in Hospitalized Patients with COVID-19 in Wuhan, China. JAMA Cardiol. 2020, 5, 802–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.; Fan, Y.; Chen, M.; Wu, X.; Zhang, L.; He, T.; Wang, H.; Wan, J.; Wang, X.; Lu, Z. Cardiovascular Implications of Fatal Outcomes of Patients With Coronavirus Disease 2019 (COVID-19). JAMA Cardiol. 2020, 5, 811–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.; Qin, M.; Cai, Y.; Liu, T.; Shen, B.; Yang, F.; Cao, S.; Liu, X.; Xiang, Y.; Zhao, Q.; et al. Characteristics and clinical significance of myocardial injury in patients with severe coronavirus disease 2019. Eur. Heart J. 2020, 41, 2070–2079. [Google Scholar] [CrossRef]
- Tay, M.Z.; Poh, C.M.; Rénia, L.; Macary, P.A.; Ng, L.F.P. The trinity of COVID-19: Immunity, inflammation and intervention. Nat. Rev. Immunol. 2020, 20, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Vaninov, N. In the eye of the COVID-19 cytokine storm. Nat. Rev. Immunol. 2020, 20, 277. [Google Scholar] [CrossRef] [PubMed]
- Connors, J.M.; Levy, J.H. Thromboinflammation and the hypercoagulability of COVID-19. J. Thromb. Haemost. 2020, 18, 1559–1561. [Google Scholar] [CrossRef]
- Wichmann, D.; Sperhake, J.P.; Lutgehetmann, M.; Steurer, S.; Edler, C.; Heinemann, A.; Heinrich, F.; Mushumba, H.; Kniep, I.; Schroder, A.S.; et al. Autopsy Findings and Venous Thromboembolism in Patients With COVID-19: A Prospective Cohort Study. Ann. Intern. Med. 2020, 173, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Schurink, B.; Roos, E.; Radonic, T.; Barbe, E.; Bouman, C.S.C.; de Boer, H.H.; de Bree, G.J.; Bulle, E.B.; Aronica, E.M.; Florquin, S.; et al. Viral presence and immunopathology in patients with lethal COVID-19: A prospective autopsy cohort study. Lancet Microbe 2020, 1, e290–e299. [Google Scholar] [CrossRef]
- Braunwald, E. Cardiomyopathies. Circ. Res. 2017, 121, 711–721. [Google Scholar] [CrossRef] [PubMed]
- CDC–NCHS–National Center for Health Statistics. Cdc. Gov. 2021. Available online: https://www.cdc.gov/nchs/index.htm (accessed on 26 May 2021).
- Rose, N.R. Critical Cytokine Pathways to Cardiac Inflammation. J. Interf. Cytokine Res. 2011, 31, 705–710. [Google Scholar] [CrossRef]
- An, N.; Gao, Y.; Si, Z.; Zhang, H.; Wang, L.; Tian, C.; Yuan, M.; Yang, X.; Li, X.; Shang, H.; et al. Regulatory Mechanisms of the NLRP3 Inflammasome, a Novel Immune-Inflammatory Marker in Cardiovascular Diseases. Front. Immunol. 2019, 10, 1592. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Duan, F.; Hu, J.; Luo, B.; Huang, B.; Lou, X.; Sun, X.; Li, H.; Zhang, X.; Yin, S.; et al. NLRP3 inflammasome-mediated pyroptosis contributes to the pathogenesis of non-ischemic di-lated cardiomyopathy. Redox Biol. 2020, 34, 101523. [Google Scholar] [CrossRef]
- Poredos, P.; Jezovnik, M.K. Endothelial Dysfunction and Venous Thrombosis. Angiology 2018, 69, 564–567. [Google Scholar] [CrossRef] [Green Version]
- Cecchini, R.; Cecchini, A.L. SARS-CoV-2 infection pathogenesis is related to oxidative stress as a response to aggression. Med. Hypotheses 2020, 143, 110102. [Google Scholar] [CrossRef] [PubMed]
- Gavriilaki, E.; Anyfanti, P.; Gavriilaki, M.; Lazaridis, A.; Douma, S.; Gkaliagkousi, E. Endothelial Dysfunction in COVID-19: Lessons Learned from Coronaviruses. Curr. Hypertens. Rep. 2020, 22, 63. [Google Scholar] [CrossRef]
- Zhang, L.; Feng, X.; Zhang, D.; Jiang, C.; Mei, H.; Wang, J.; Zhang, C.; Li, H.; Xia, X.; Kong, S.; et al. Deep Vein Thrombosis in Hospitalized Patients With COVID-19 in Wuhan, China. Circulation 2020, 142, 114–128. [Google Scholar] [CrossRef]
- Bilaloglu, S.; Aphinyanaphongs, Y.; Jones, S.; Iturrate, E.; Hochman, J.; Berger, J.S. Thrombosis in Hospitalized Patients With COVID-19 in a New York City Health System. JAMA 2020, 324, 799–801. [Google Scholar] [CrossRef]
- Spyropoulos, A.C. The management of venous thromboembolism in hospitalized patients with COVID-19. Blood Adv. 2020, 4, 4028. [Google Scholar] [CrossRef] [PubMed]
- Von Bruhl, M.L.; Stark, K.; Steinhart, A.; Chandraratne, S.; Konrad, I.; Lorenz, M.; Khandoga, A.; Tirniceriu, A.; Coletti, R.; Kollnberger, M.; et al. Monocytes, neutrophils, and platelets cooperate to initiate and propagate venous thrombosis in mice in vivo. J. Exp. Med. 2012, 209, 819–835. [Google Scholar] [CrossRef] [PubMed]
- Brill, A.; Fuchs, T.A.; Savchenko, A.S.; Thomas, G.; Martinod, K.; De Meyer, S.F.; Bhandari, A.A.; Wagner, D.D. Neutrophil extracellular traps promote deep vein thrombosis in mice. J. Thromb. Haemost. 2012, 10, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, T.A.; Brill, A.; Wagner, D.D. Neutrophil Extracellular Trap (NET) Impact on Deep Vein Thrombosis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1777–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Fu, B.; Zheng, X.; Wang, D.; Zhao, C.; Qi, Y.; Sun, R.; Tian, Z.; Xu, X.; Wei, H. Pathogenic T cells and inflammatory monocytes incite inflammatory storm in severe COVID-19 patients. Natl. Sci. Rev. 2020, 7, 998–1002. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Guo, R.; Lei, L.; Liu, H.; Wang, Y.; Wang, Y.; Dai, T.; Zhang, T.; Lai, Y.; Wang, J.; et al. COVID-19 infection induces readily detectable morphological and inflammation-related phenotypic changes in peripheral blood monocytes, the severity of which correlate with patient outcome. medRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus–Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef]
- Skaggs, B.J.; Hahn, B.H.; McMahon, M. Accelerated atherosclerosis in patients with SLE—mechanisms and management. Nat. Rev. Rheumatol. 2012, 8, 214–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasceri, V.; Willerson, J.T.; Yeh, E.T.H. Direct Proinflammatory Effect of C-Reactive Protein on Human Endothelial Cells. Circulation 2000, 102, 2165–2168. [Google Scholar] [CrossRef]
- Leibovitz, E.; Hertz, Y.; Liberman, E.; Sclarovsky, S.; Berliner, S. Increased adhesiveness of white blood cells in patients with unstable angina: Additional evidence for an involvement of the immune-inflammatory system. Clin. Cardiol. 1997, 20, 1017–1020. [Google Scholar] [CrossRef]
- Min, X.; Lu, M.; Tu, S.; Wang, X.; Zhou, C.; Wang, S.; Pang, S.; Qian, J.; Ge, Y.; Guo, Y.; et al. Serum Cytokine Profile in Relation to the Severity of Coronary Artery Disease. BioMed Res. Int. 2017, 2017, 4013685. [Google Scholar] [CrossRef]
- Ali, N. Elevated level of C-reactive protein may be an early marker to predict risk for severity of COVID-19. J. Med Virol. 2020, 92, 2409–2411. [Google Scholar] [CrossRef]
- Zhao, K.; Li, R.; Wu, X.; Zhao, Y.; Wang, T.; Zheng, Z.; Zeng, S.; Ding, X.; Nie, H. Clinical features in 52 patients with COVID-19 who have increased leukocyte count: A retrospective analysis. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 2279–2287. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine Storm in COVID-19: The Current Evidence and Treatment Strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef] [PubMed]
- COVID-19 and vascular disease. EBioMedicine 2020, 58, 102966. [CrossRef] [PubMed]
- Nishiga, M.; Wang, D.W.; Han, Y.; Lewis, D.B.; Wu, J.C. COVID-19 and cardiovascular disease: From basic mechanisms to clinical perspectives. Nat. Rev. Cardiol. 2020, 17, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.J.; Liang, W.H.; Zhao, Y.; Liang, H.-R.; Chen, Z.-S.; Li, Y.-M.; Liu, X.-Q.; Chen, R.-C.; Tang, C.-L.; Wang, T.; et al. Comorbidity and its impact on 1590 patients with COVID-19 in China: A nationwide analysis. Eur. Respir. J. 2020, 55, 2000547. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D.A.; Stashenko, G.J.; Akay, O.M.; Price, L.I.; Owzar, K.; Ginsburg, G.S.; Chi, J.T.; Ortel, T.L. Whole blood gene expression analyses in patients with single versus recurrent venous thromboembolism. Thromb. Res. 2011, 128, 536–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravi, S.; Schuck, R.N.; Hilliard, E.; Lee, C.R.; Dai, X.; Lenhart, K.; Willis, S.; Jensen, B.C.; Stouffer, G.A.; Patterson, C.; et al. Clinical Evidence Supports a Protective Role for CXCL5 in Coronary Artery Disease. Am. J. Pathol. 2017, 187, 2895–2911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RNA-seq of Platelets from SARS-CoV-2 Covid-19; University of Utah: Salt Lake City, UT, USA, 2020.
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdóttir, H.; Tamayo, P.; Mesirov, J.P. Molecular signatures database (MSigDB) 3.0. Bioinformatics 2011, 27, 1739–1740. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Khodadoust, M.S.; Liu, C.L.; Newman, A.M.; Alizadeh, A.A. Profiling Tumor Infiltrating Immune Cells with CIBERSORT. Methods Mol. Biol. 2018, 1711, 243–259. [Google Scholar] [PubMed]
- Shin, J.S.; Greer, A.M. The role of FcepsilonRI expressed in dendritic cells and monocytes. Cell Mol. Life Sci. 2015, 72, 2349–2360. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.Y.; Morten, B.C.; Antony, J.; Henderson, L.; Lasham, A.; Campbell, H.; Cunliffe, H.; Horsfield, J.A.; Reddel, R.R.; Avery-Kiejda, K.A.; et al. Regulation of the interferon-gamma (IFN-gamma) pathway by p63 and Delta133p53 isoform in different breast cancer subtypes. Oncotarget 2018, 9, 29146–29161. [Google Scholar] [CrossRef] [Green Version]
- Keating, S.E.; Maloney, G.M.; Moran, E.M.; Bowie, A.G. IRAK-2 participates in multiple toll-like receptor signaling pathways to NFkappaB via activation of TRAF6 ubiquitination. J. Biol. Chem. 2007, 282, 33435–33443. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.; Gawanbacht, A.; Habjan, M.; Rang, A.; Borner, C.; Schmidt, A.M.; Veitinger, S.; Jacob, R.; Devignot, S.; Kochs, G.; et al. Incoming RNA Virus Nucleocapsids Containing a 5′-Triphosphorylated Genome Activate RIG-I and Antiviral Signaling. Cell Host Microbe 2013, 13, 336–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Liu, Z.; Ke, Y.; Wang, F. Intrinsic FGFR2 and Ectopic FGFR1 Signaling in the Prostate and Prostate Cancer. Front. Genet. 2019, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Hamanaka, N. Eicosanoids in Mammals. Compr. Nat. Prod. Chem. 1999, 1, 159–206. [Google Scholar]
- Mukherjee, S.; Kumar, R.; Tsakem Lenou, E.; Basrur, V.; Kontoyiannis, D.L.; Ioakeimidis, F.; Mosialos, G.; Theiss, A.L.; Flavell, R.A.; Venuprasad, K. Deubiquitination of NLRP6 inflammasome by Cyld critically regulates intestinal inflammation. Nat. Immunol. 2020, 21, 626–635. [Google Scholar] [CrossRef]
- Castellino, F.; Huang, A.Y.; Altan-Bonnet, G.; Stoll, S.; Scheinecker, C.; Germain, R.N. Chemokines enhance immunity by guiding naive CD8+ T cells to sites of CD4+ T cell–dendritic cell interaction. Nature 2006, 440, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Szauter, K.M.; Jansen, M.K.; Okimoto, G.; Loomis, M.; Kimura, J.H.; Heller, M.; Ku, T.; Tiirikainen, M.; Boyd, C.D.; Csiszar, K.; et al. Persistent Inflammatory Pathways Associated with Early Onset Myocardial Infarction in a Medicated Multiethnic Hawaiian Cohort. Biochem. Insights 2011, 4, BCI.S6976. [Google Scholar] [CrossRef] [Green Version]
- Casson, C.; Yu, J.; Reyes, V.M.; Taschuk, F.O.; Yadav, A.; Copenhaver, A.M.; Nguyen, H.T.; Collman, R.G.; Shin, S. Human caspase-4 mediates noncanonical inflammasome activation against gram-negative bacterial pathogens. Proc. Natl. Acad. Sci. USA 2015, 112, 6688–6693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Liu, Y.; Cao, L.; Wang, D.; Guo, M.; Jiang, A.; Guo, D.; Hu, W.; Yang, J.; Tang, Z.; et al. Transcriptomic characteristics of bronchoalveolar lavage fluid and peripheral blood mononucle-ar cells in COVID-19 patients. Emerg. Microbes Infect. 2020, 9, 761–770. [Google Scholar] [CrossRef]
- Kandikattu, H.K.; Venkateshaiah, S.U.; Mishra, A. Synergy of Interleukin (IL)-5 and IL-18 in eosinophil mediated pathogenesis of allergic diseases. Cytokine Growth Factor Rev. 2019, 47, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Verbist, K.C.; Klonowski, K.D. Functions of IL-15 in anti-viral immunity: Multiplicity and variety. Cytokine 2012, 59, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Abers, M.S.; Delmonte, O.M.; Ricotta, E.E.; Fintzi, J.; Fink, D.L.; de Jesus, A.A.A.; Zarember, K.A.; Alehashemi, S.; Oikonomou, V.; Desai, J.V.; et al. An immune-based biomarker signature is associated with mortality in COVID-19 patients. JCI Insight 2021, 6. [Google Scholar] [CrossRef]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Bansal, S.S.; Ismahil, M.A.; Goel, M.; Zhou, G.; Rokosh, G.; Hamid, T.; Prabhu, S.D. Dysfunctional and Proinflammatory Regulatory T-Lymphocytes Are Essential for Ad-verse Cardiac Remodeling in Ischemic Cardiomyopathy. Circulation 2019, 139, 206–221. [Google Scholar] [CrossRef]
- Bansal, S.S.; Ismahil, M.A.; Goel, M.; Patel, B.; Hamid, T.; Rokosh, G.; Prabhu, S.D. Activated T Lymphocytes are Essential Drivers of Pathological Remodeling in Ischemic Heart Failure. Circ. Heart Fail. 2017, 10, e003688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanton, R.M.; Carrillo-Salinas, F.J.; Alcaide, P. T-cell recruitment to the heart: Friendly guests or unwelcome visitors? Am. J. Physiol. Heart Circ. Physiol. 2019, 317, H124–H140. [Google Scholar] [CrossRef]
- Merad, M.; Martin, J.C. Pathological inflammation in patients with COVID-19: A key role for monocytes and macrophages. Nat. Rev. Immunol. 2020, 20, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.T.; Chen, J.W. Emerging role of chemokine CC motif ligand 4 related mechanisms in diabetes mellitus and cardiovas-cular disease: Friends or foes? Cardiovasc. Diabetol. 2016, 15, 117. [Google Scholar] [CrossRef] [Green Version]
- Papoff, G.; Presutti, D.; Lalli, C.; Bolasco, G.; Santini, S.; Manelfi, C.; Fustaino, V.; Alemà, S.; Ruberti, G. CASP4 gene silencing in epithelial cancer cells leads to impairment of cell migration, cell-matrix adhesion and tissue invasion. Sci. Rep. 2018, 8, 17705. [Google Scholar] [CrossRef]
- Malas, M.B.; Naazie, I.N.; Elsayed, N.; Mathlouthi, A.; Marmor, R.; Clary, B. Thromboembolism risk of COVID-19 is high and associated with a higher risk of mortality: A systematic review and meta-analysis. EClinicalMedicine 2020, 29–30, 100639. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.K.; Mainbourg, S.; Friggeri, A.; Bertoletti, L.; Douplat, M.; Dargaud, Y.; Grange, C.; Lobbes, H.; Provencher, S.; Lega, J.-C. Arterial and venous thromboembolism in COVID-19: A study-level meta-analysis. Thorax 2021. [Google Scholar] [CrossRef]
- Clerkin, K.J.; Fried, J.A.; Raikhelkar, J.; Sayer, G.; Griffin, J.M.; Masoumi, A.; Jain, S.S.; Burkhoff, D.; Kumaraiah, D.; Rabbani, L.; et al. COVID-19 and Cardiovascular Disease. Circulation 2020, 141, 1648–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, E.; van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Collins, R.G.; Velji, R.; Guevara, N.V.; Hicks, M.J.; Chan, L.; Beaudet, A.L. P-Selectin or intercellular adhesion molecule (ICAM)-1 deficiency substantially protects against atherosclerosis in apolipoprotein E-deficient mice. J. Exp. Med. 2000, 191, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Brasier, A.R. The nuclear factor-kappaB-interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 2010, 86, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gareus, R.; Kotsaki, E.; Xanthoulea, S.; van der Made, I.; Gijbels, M.J.; Kardakaris, R.; Polykratis, A.; Kollias, G.; de Winther, M.P.; Pasparakis, M. Endothelial cell-specific NF-kappaB inhibition protects mice from atherosclerosis. Cell Metab. 2008, 8, 372–383. [Google Scholar] [CrossRef] [Green Version]
- Merhi-Soussi, F.; Kwak, B.; Magne, D.; Chadjichristos, C.; Berti, M.; Pelli, G.; James, R.W.; Mach, F.; Gabay, C. Interleukin-1 plays a major role in vascular inflammation and atherosclerosis in male apolipoprotein E-knockout mice. Cardiovasc. Res. 2005, 66, 583–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, Y.; Raines, E.W.; Plump, A.S.; Breslow, J.L.; Ross, R. Upregulation of VCAM-1 and ICAM-1 at atherosclerosis-prone sites on the endothelium in the ApoE-deficient mouse. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 842–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tikellis, C.; Jandeleit-Dahm, K.; Sheehy, K.; Murphy, A.; Chin-Dusting, J.; Kling, D.; Sebokova, E.; Cooper, M.; Mizrahi, J.; Woollard, K. Reduced plaque formation induced by rosiglitazone in an STZ-diabetes mouse model of atherosclerosis is associated with downregulation of adhesion molecules. Atherosclerosis 2008, 199, 55–64. [Google Scholar] [CrossRef]
- Pamukcu, B.; Lip, G.Y.H.; Devitt, A.; Griffiths, H.R.; Shantsila, E. The role of monocytes in atherosclerotic coronary artery disease. Ann. Med. 2010, 42, 394–403. [Google Scholar] [CrossRef]
- Wang, W.; Ye, L.; Ye, L.; Li, B.; Gao, B.; Zeng, Y.; Kong, L.; Fang, X.; Zeng, H.; Wu, Z.; et al. Up-regulation of IL-6 and TNF-alpha induced by SARS-coronavirus spike protein in murine macrophages via NF-kappaB pathway. Virus Res. 2007, 128, 1–8. [Google Scholar] [CrossRef]
- McGonagle, D.; Sharif, K.; O’Regan, A.; Bridgewood, C. The Role of Cytokines including Interleukin-6 in COVID-19 induced Pneumonia and Macrophage Activation Syndrome-Like Disease. Autoimmun. Rev. 2020, 19, 102537. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Articles Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Hirano, T.; Murakami, M. COVID-19: A New Virus, but a Familiar Receptor and Cytokine Release Syndrome. Immun. 2020, 52, 731–733. [Google Scholar] [CrossRef]
- Ye, Q.; Wang, B.; Mao, J. The pathogenesis and treatment of the ‘Cytokine Storm’ in COVID-19. J. Infect. 2020, 80, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Madjid, M.; Safavi-Naeini, P.; Solomon, S.D.; Vardeny, O. Potential Effects of Coronaviruses on the Cardiovascular System: A Review. JAMA Cardiol 2020, 5, 831–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweet, M.E.; Cocciolo, A.; Slavov, D.; Jones, K.L.; Sweet, J.R.; Graw, S.L.; Reece, T.B.; Ambardekar, A.V.; Bristow, M.R.; Mestroni, L.; et al. Transcriptome analysis of human heart failure reveals dysregulated cell adhesion in dilated cardiomyopathy and activated immune pathways in ischemic heart failure. BMC Genom. 2018, 19, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, A.C.; Castaneda, G.; Li, W.T.; Chen, C.; Shende, N.; Chakladar, J.; Taub, P.R.; Chang, E.Y.; Ongkeko, W.M. COVID-19 Severity Potentially Modulated by Cardiovascular-Disease-Associated Immune Dysregulation. Viruses 2021, 13, 1018. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061018
Lee AC, Castaneda G, Li WT, Chen C, Shende N, Chakladar J, Taub PR, Chang EY, Ongkeko WM. COVID-19 Severity Potentially Modulated by Cardiovascular-Disease-Associated Immune Dysregulation. Viruses. 2021; 13(6):1018. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061018
Chicago/Turabian StyleLee, Abby C., Grant Castaneda, Wei Tse Li, Chengyu Chen, Neil Shende, Jaideep Chakladar, Pam R. Taub, Eric Y. Chang, and Weg M. Ongkeko. 2021. "COVID-19 Severity Potentially Modulated by Cardiovascular-Disease-Associated Immune Dysregulation" Viruses 13, no. 6: 1018. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061018