
From A to m6A: The Emerging Viral Epitranscriptome


Virology Unit, Department of Experimental and Health Sciences, Universitat Pompeu Fabra, 08003 Barcelona, Spain
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Viruses 2021, 13(6), 1049; https://0-doi-org.brum.beds.ac.uk/10.3390/v13061049
Submission received: 30 April 2021
/
Revised: 21 May 2021
/
Accepted: 23 May 2021
/
Published: 1 June 2021
(This article belongs to the Special Issue RNA-Binding Proteins as Cellular Targets to Mediate the Virus Lifecycle)
Abstract
:There are over 100 different chemical RNA modifications, collectively known as the epitranscriptome. N6-methyladenosine (m6A) is the most commonly found internal RNA modification in cellular mRNAs where it plays important roles in the regulation of the mRNA structure, stability, translation and nuclear export. This modification is also found in viral RNA genomes and in viral mRNAs derived from both RNA and DNA viruses. A growing body of evidence indicates that m6A modifications play important roles in regulating viral replication by interacting with the cellular m6A machinery. In this review, we will exhaustively detail the current knowledge on m6A modification, with an emphasis on its function in virus biology.
1. Introduction
Because of their limited coding capacity, viruses completely depend on the host machinery to multiply. This intimate dependency occurs at multiple layers whose components and features we are still far from understanding. A novel layer recently uncovered is the interaction of viruses with the host RNA modification machinery. Since the discovery of the first chemical RNA modification, pseudouridine (Ψ), in 1957 [1], more than 100 different RNA modifications have been described, collectively known as the epitranscriptome. These modifications occur post-transcriptionally mainly in non-coding RNAs (ncRNA), such as transfer RNA (tRNA) and ribosomal RNA (rRNA). Nevertheless, ten different modifications also occur in mRNAs [2], of which some of the most studied include N6-methyladenosine (m6A), N5-methylcytosine (m5C), and 2′-O-methylation (or Nm, where N stands for any nucleotide).
The m6A modification, discovered in 1974 [3,4], entails of the addition of a methyl group to an adenosine base at the nitrogen-6 position. The m5C modification, discovered in 1958 [5], entails the addition of a methyl group to a cytosine base at the carbon-5 position. Finally, the Nm modification, discovered in 1967 [6], involves the addition of a methyl group to the 2′-hydroxyl group of the ribose moiety.
The m6A modification, is a dynamic process that can be reversed in specific contexts [7,8]. For other RNA modifications, the proteins that catalyse the addition of the chemical modification are well established [9], yet, the precise molecular mechanisms and which proteins interact with all these diverse modifications remain unknown for the large majority. A major limitation has been the lack of reliable and technically accessible methods that allow for precise and quantitative detection of RNA modifications. However, recent methodological advances are rapidly changing this situation. Initial analyses of RNA modifications involved the development of enzymatic digestions of RNA coupled to liquid chromatography-tandem mass spectrometry (LC-MS/MS). While this approach allows for accurate quantification of RNA modifications [10], it does not provide any information on their location. Moreover, the purity of the RNA sample is essential for correct quantifications. These two limitations were overcome for some RNA modifications with the rise of second-generation sequencing that allow mapping them at single-nucleotide resolution in a transcriptome-wide manner [11]. In addition, the recent development of third-generation nanopore RNA sequencing now allows for real-time detection of some RNA modifications at single-nucleotide resolution in native full-length RNA sequences [12,13].
All these recent methodological advances have accelerated our knowledge on the functions of RNA modifications in the cell. These include modulation of RNA structure, facilitating nuclear export, translational activation and regulation of RNA stability [14]. In agreement with these key molecular roles, a myriad of fundamental physiological processes is fine-tuned by RNA modifications including cell differentiation [15], brain development [16], embryogenesis [17], fertility [8], the immune system [18] and the circadian clock [19]. As a consequence, it is not a surprise that alterations in the cellular epitranscriptomic landscape are linked to a wide-range of diseases such as cancer, auto-immunity, neurological and genetic disorders [20,21,22,23]. Excitingly, RNA modifications also play key roles in viral infections. Viral RNA genomes and viral mRNAs are chemically modified during infection and these modifications seem to play fundamental roles in viral life cycles [24,25,26,27]. Moreover, viral infections alter the host epitranscriptomic landscape and these global changes have been shown to affect viral replication [28]. Because of methodological limitations, these viral studies have been mainly focused on m6A modifications. In this review, we will summarise the current knowledge of the interplay between m6A modifications and viral infection. We will also highlight current limitations in the field and propose key future directions.
2. N6-Methyladenosine (m6A)
The m6A modification is the most abundant internal modification in cellular mRNA transcripts. Around 25% of all cellular mRNA transcripts are thought to be m6A-modified, mainly around the translation stop codon and at the 3′ untranslated region (3′UTR) [29,30]. Because of this abundance and of the recent ability to accurately map its location in a transcriptome-wide manner, m6A is the most studied RNA modification. In addition to mRNAs, m6A modifications are found in long non-coding RNAs (lncRNAs) [31], circular RNAs [32], the 18S rRNA and 28S rRNA [9], primary microRNAs (pri-miRNAs) [33] and small nuclear RNAs (snRNAs) [34].
The regulation of the m6A modification landscape in host mRNAs is a dynamic process that involves m6A-methyltransferases to add the modification (writers), m6A-binding proteins to recognise and bind to m6A modifications (readers) and m6A-demethylases to remove the methyl mark (erasers) [2] (Figure 1). The addition of m6A methylations occurs co-transcriptionally in the nucleus and is primarily catalyzed by the METTL3–METTL14 methyltransferase complex [35]. In this complex, METTL3 serves as the catalytic subunit and METTL14 as the allosteric adaptor that maintains the complex integrity and binds the complex to the target RNA [36]. Consistently, METTL3 knockout in mouse embryonic stem cells led to a near-complete depletion of m6A on mRNAs [37]. The METTL3–METTL14 complex also includes different adaptor proteins, of which the most important is Wilms tumor associated protein (WTAP). This protein is required for the localization of the METTL3–METTL14 complex into nuclear speckles, where pre-mRNA splicing occurs, and for the catalytic activity of the METTL3–METTL14 complex [38]. In the absence of WTAP, RNA-binding capacity of the METTL3–METTL14 complex is strongly reduced [38]. The METTL3–METTL14 complex adds the m6A modification in mRNAs at the consensus sequence DRACH (D = A, G or U; R = G or A; H = A, C or U) [2]. However, not all DRACH motifs are m6A-modified and around 20% of m6A modifications occur outside the DRACH motif [39]. To allow reversible regulation, m6A modifications can be erased by the m6A demethylases fat mass and obesity-associated protein (FTO) [7] and AlkB homolog 5 (ALKBH5) [8]. However, the widespread reversibility of the process has been challenged [40] and the main substrate of FTO has been proposed to be N6,2′-O-dimethyladenosine (m6Am) [41].
The m6A marks in mRNA molecules are primarily recognised by the five YT521-B homology (YTH) domain-containing proteins YTHDF1, YTHDF2, YTHDF3, YTHDC1 and YTHDC2. Structural crystallographic studies revealed that m6A is directly recognised and bound by a conserved aromatic cage structure present in the YTH domain of YTH readers [42]. This domain recognises RNA in an m6A-dependent manner without sequence selectivity and regardless of the RNA length [43,44]. A recent RNA affinity assay coupled to mass spectrometry analysis also identified YTHDF1–3 and YTHDC1 as m1A readers, however, their affinity was lower for m1A RNA baits than for m6A baits [45]. Although initially it was proposed that each YTH reader plays different roles, recent evidence suggests a model in which YTH readers behave redundantly [46,47,48]. A major effort from multiple laboratories has been done to identify the role of YTH readers. The main findings are, first, YTHDF1 promotes CAP-dependent translation by interacting with the 5′ UTR-associated eIF3 protein [49]. Interestingly, m6A modification in 5′UTRs can also promote CAP-independent translation by directly recruiting eIF3 [50]. Second, YTHDF2 is the main YTH reader that regulates mRNA stability. It recruits the CCR4–NOT deadenylase complex, ultimately promoting degradation of m6A-modified RNAs [46,51]. Third, YTHDF3 supports the function of both YTHDF1 and YTHDF2 by binding them in an RNA-independent manner [47,52]. It has been suggested that YTHDF3 would be the first reader to interact with m6A and that this binding would facilitate the access to YTHDF1 or YTHDF2 [47]. Finally, YTHDC1 affects splicing of m6A-containing pre-mRNAs [53] and promotes nuclear export of m6A-decorated mRNAs [54] and the helicase YTHDC2, which is primarily but not exclusively expressed in testes, positively regulating translation of structured mRNAs containing m6A modifications located in their coding sequence [55]. Moreover, YTHDC2 plays important roles in spermatogenesis [56,57].
Additional m6A readers have been recently identified. These include IGF2BP proteins (IGF2BP1, IGF2BP2 and IGF2BP3) [58], eIF3 [50], FMRP, FXR1, FXR2 [59,60,61], SND1 [59] and three heterogeneous nuclear ribonucleoproteins, hnRNPA2B1 [33], hnRNPC [62] and hnRNPG [63]. However, their mode of binding to m6A-modified mRNAs and their role remains to be completely understood. Interestingly, the top two motifs bound by FMRP identified by PAR-CLIP-m6A-seq [64] are almost identical to the two top motifs bound by SND1 in m6A-modified exons [59], the shared motifs being UGGAC and CU(A/U)CG. Intriguingly, FXR1, FXR2, FMRP and SND1 belong to the Tudor domain ‘Royal family’ and contain an aromatic cage which is structurally similar to the one found in YTH readers [65]. Therefore, these proteins might potentially bind m6A directly using this aromatic cage. This does not seem to be the case for the three heterogeneous nuclear ribonucleoproteins. No obvious aromatic cage was identified in the tandem RNA recognition motif (RRM) domains of hnRNPA2B1 [66]. Moreover, hnRNPA2B1 displayed slightly higher affinity to unmethylated RNA than m6A-modified RNA in isothermal titration calorimetry experiments [66]. This indicates that hnRNPA2B1 is an indirect m6A reader, which may require further protein interactors to bind m6A sites. Finally, the hnRNPC and hnRNPG proteins also bind indirectly to m6A modifications [62,63]. Importantly, their binding occurs in an RNA secondary structure-dependent manner and it has been suggested that hnRNPA2B1 might use this mode of binding as well [66]. The limited literature available on all these unconventional readers suggests that they bind fewer m6A sites compared with the promiscuous YTH readers [59,66]. Future crystallographic or nuclear magnetic resonance (NMR) studies will be necessary to further clarify their mode of binding to m6A-decorated transcripts.
The location of the cellular m6A machinery has also been thoroughly investigated. Although both writers and erasers are mainly nuclear, they are also found in the cytoplasm [67,68,69,70]. A fraction of FTO has been described to shuttle between the nucleus and cytoplasm [71]; moreover, its subcellular localization was cell type-dependent [69]. Regarding the readers, YTHDC1 is exclusively nuclear while YTHDC2 locates both in the nucleus and in the cytoplasm [55]. The rest of YTHDF readers are widely believed to be exclusively cytoplasmic, however, recent studies also detected YTHDF1, YTHDF2 and YTHDF3 in the nucleus of different cell lines [55,70]. The current availability of high quality endogenous antibodies does now allow, at least for some proteins, detailed characterization of their cellular location under physiological and perturbed conditions.
The m6A modification plays key roles in mRNA fates. It is described to modulate mRNA structure [62], splicing [53], nuclear export [72], translation [49,73], stability [47,51] and mRNA localization in stress granules [74]. Under physiological conditions, m6A affects a variety of processes such as fertility [8], stem cell renewal capability [75], embryogenesis [17], neuronal development [16], adipogenesis [76] and immune regulation [18]. Importantly, dysregulation of the m6A RNA methylation pathway has been connected to the development of a range of human diseases [23] including cancer [77], autoimmune diseases [78] and neurological disorders [79]. Moreover, m6A modifications have been extensively reported in viral RNAs and shown to regulate viral life cycles [27,80].
Currently, most methods to detect m6A modifications transcriptome-wide combine immunoprecipitations with an anti-m6A specific antibody coupled to high-throughput sequencing. The most technically accessible technique, N6-methyladenosine-sequencing (m6A-seq) (Figure 2A), developed in 2012 [29,30], consists of an initial chemical fragmentation step in which mRNAs are reduced to 100–200 nucleotides in length. An input sample is then saved as control and the rest of the fragmented mRNAs are subjected to m6A-immunoprecipitation. Following reverse transcription and high-throughput sequencing of input and m6A-immunoprecipitated samples, bioinformatic analyses similar to those in ChIP-seq are applied to identify regions enriched for m6A decorations, which are commonly referred as m6A peaks. Major drawbacks of this method include the lack of single-nucleotide resolution, as it solely provides an insight into the location of m6A modifications within a window of 100–200 nucleotides, lack of isoform-specificity and m6A stoichiometry (m6A abundance at a given site), and a low reproducibility. Even when m6A-seq was performed in the same cell lines, the m6A peaks in mRNAs overlapped only 30% to 60% between different studies [81]. In recent years, several novel techniques with improved m6A resolution are driving our knowledge in the mechanisms and functions underlying m6A modification. These include antibody-dependent and independent methods. Two antibody-dependent methods use UV-crosslinking of anti-m6A specific antibodies to the m6A-modified RNA, the photo-crosslinking-assisted m6A-sequencing (PA-m6A-seq) [82] (Figure 2B) and the m6A individual-nucleotide-resolution crosslinking and immunoprecipitation (miCLIP) [39]. PA-m6A-seq pinpoints m6A within a window of ≈ 23 nucleotides while miCLIP provides single-nucleotide resolution, however, in contrast to m6A-seq, miCLIP requires the use of radioactivity during cDNA library preparation.
In the last five years, multiple new methods have been developed to improve the detection of m6A modifications. The m6A-level and isoform-characterization sequencing (m6A-LAIC-seq) [83] allows, after m6A-immunoprecipitation of full-length RNAs, quantifying m6A stoichiometry in a transcriptome-wide manner. Another recent antibody-independent method, named meCLICK-seq, utilises click chemistry to attach small molecules to any type of m6A-methylated RNA followed by specific cleavage and degradation of these RNAs [84]. By comparing cells treated with and without a click-degrader that catalyses the cleavage of RNA, depletion of modified RNA species is identified across the transcriptome. However, none of these two methods offer an insight into the location of m6A sites in the transcripts. To overcome this, four new antibody-independent methods named DART-seq [85], MAZTER-seq [86], SEAL-seq [87] and m6A-label-seq [88] allow transcriptome-wide m6A mapping. DART-seq (Deamination Adjacent to RNA modification Targets) enables global m6A profiling using as little as 10 ng of total RNA to estimate m6A abundance in individual RNAs. This elegant technique is based on the fusion of the cytidine deaminase APOBEC1 to the m6A-binding YTH domain of YTHDF2. Transfection of the APOBEC1–YTH fusion protein in cells induces C-to-U deamination at sites adjacent to m6A modifications and after isolation of total RNA these editing events are identified via standard RNA-seq. MAZTER-seq is a new enzymatic method that takes advantage of the bacterial RNase MazF, which cleaves the RNA immediately upstream of an unmethylated ACA sequence but not an m6A-CA sequence. MAZTER-seq offers quantification of m6A stoichiometry at single-nucleotide resolution but only at 16–25% of all m6A methylation sites. SEAL-seq is an FTO-assisted chemical labeling method that shows high false-negative and high false-positive rates compared with other m6A sequencing methods. In this technique, first, the RNA demethylase FTO is expressed in bacteria and purified. Then, the RNA of interest is fragmented and treated with FTO, which oxidises m6A to the unstable N6-hydroxymethyladenosine (hm6A). Second, dithiothreitol (DTT) is added to convert hm6A to the more stable N6-dithiolsitolmethyladenosine (dm6A). The free sulfhydryl group present in dm6A then allows for biotin labelling and purification of the dm6A-containing RNAs with streptavidin beads. Recovered RNAs are finally subjected to library construction and deep-sequencing. Finally, m6A-label-seq is a metabolic labeling method that detects m6A at base resolution and is superior in characterising clustered m6A sites. In this method, cells are fed with a methionine analog which substitutes the methyl group on the METTL3 cofactor S-adenosyl methionine (SAM) with the allyl group. Cellular RNAs are therefore metabolically modified with N6-allyladenosine (a6A) instead of m6A modification. Following iodination-induced cyclization, a6A is converted to Cyc-A which during reverse transcription induces misincorporation at the opposite site in cDNA, thus following deep-sequencing A-to-C/T/G mutations can be identified. Another recent exciting approach to obtain antibody-independent localization of m6A modifications is based on nanopore sequencing [89,90]. In this technology, protein nanopores are embedded into a synthetic membrane. When an ionic current is applied and RNA molecules move through these nanopores, the corresponding disruption of the current intensity provides information to identify both the sequence and the modified RNA nucleotide. This approach offers single-nucleotide specificity, isoform-specificity and very long reads without the need of a reverse transcription step. To date, three studies have already used this technology to decipher the epitranscriptome of viral RNA genomes [91,92,93].
3. m6A Modifications and Viral Replication
Early studies carried out more than 40 years ago identified m6A modifications in simian virus 40 (SV40) [94], adenovirus [95], influenza A virus (IAV) [96,97], herpes simplex virus [98], avian sarcoma virus [99] and Rous sarcoma virus (RSV) [100,101]. Since then, m6A modifications have been found in many other viral mRNAs, transcribed from DNA and RNA viruses, and retroviruses and in viral RNA genomes. To date, members of the Flaviviridae, Orthomyxoviridae, Paramyxoviridae, Retroviridae, Togaviridae, Picornaviridae, Polyomaviridae, Hepadnaviridae, Adenoviridae, Rhabdoviridae, Herpesviridae and Coronaviridae viral families (Table 1) have been reported to contain m6A modifications. Most intracellular steps of the viral life cycle have already been described to be affected by m6A modifications (Figure 3). These effects were pro- or anti- viral depending on the virus (Table 2). To address the role of m6A modification in viral RNAs, the general approach has been to manipulate the key components of the m6A cellular machinery via overexpression or depletion, or to mutate the DRACH motifs within the viral RNAs to abolish specific m6A modifications. It is important to note that in the overexpression and depletion studies, the observed effects on virus replication might be indirect as the cellular RNA metabolism will inevitably be affected. In turn, altering the DRACH motif at a specific location in viral RNAs might cause effects beyond those caused by abrogating m6A RNA modification as genetic information in viral genomes is highly compacted. Thus, to ascertain a role of specific m6A modifications in the viral RNAs, at least a combination of these two approaches would be needed. However, to date few studies incorporate mutational testing within the DRACH motifs in viral mRNAs or viral RNA genomes. Of note, the effect of altering the m6A machinery on viral infection might change depending on the cell type used, indicating the high complexity of m6A epitranscriptomic regulation during viral infections [102]. The next sections will summarise in detail the current findings on the role of m6A modifications in both RNA and DNA viruses.
4. Epitranscriptomic Regulation of RNA Viruses
Most studies on the role of m6A modifications in RNA viruses focused on single-stranded RNA viruses. These include the positive-sense RNA dengue (DENV), zika (ZIKV), West-Nile (WNV) and the hepatitis C (HCV) viruses, all in the Flaviviridae family, the negative-sense RNA influenza virus (IAV) and respiratory syncytial (RSV) viruses and the retrovirus human immunodeficiency virus serotype 1 (HIV-1). All these viruses except IAV and HIV-1, carry out their viral cycles exclusively in the cytoplasm. This poses an intriguing question as both the writing and erasing m6A RNA modification machineries are believed to function mainly in the nucleus and re-localization of the involved enzymes into the cytoplasm was not observed in flavivirus or RSV infections [67,68,70]. However, recent studies in enterovirus 71 (EV71) and in human metapneumovirus (HMPV)-infected cells demonstrated that writers and erasers relocate from the nucleus to the cytoplasm [112,113]. Moreover, in HMPV infection, METTL14 strongly co-localised with the viral N protein in cytoplasmic inclusion bodies [113], the sites of HMPV replication. Further confocal microscopy analyses are thus required to determine whether the key RNA-modifying enzymes concentrate in other RNA viral replication factories in the cytoplasm.
In line with a complex RNA virus-m6A machinery interplay, currently there is no global pro- or anti-viral role of m6A modification that can be generalised. Current data support a pro-viral role for some viruses such as HIV-1 and IAV, and an anti-viral role for others, such as HCV, ZIKV and porcine epidemic diarrhea virus (PEDV) (Table 2).
4.1. m6A and HIV-1
The HIV-1 genome consists of one single-stranded positive-sense RNA that is found as a dimer and is retrotranscribed in the cytosol of the infected cell into one molecule of DNA that ultimately is integrated into the genomic DNA. Once integrated, the viral DNA exploits the nuclear machinery to transcribe the viral mRNAs and the genomic RNA. These will be exported to the cytosol where they will be translated and, in the case of the genomic RNA, encapsidated to produce the new viral progeny.
HIV-1 RNA contains multiple m6A modifications [103,104,106]. In addition, HIV-1 infection causes a global increase of m6A modifications in host mRNAs [103]. Interestingly, this increase does not require HIV-1 replication per se. Binding of the HIV-1 gp120 envelope protein to the CD4 receptor in T lymphocytes is sufficient to induce the global increase in m6A modifications in cellular mRNAs [125]. Whether and how this affects HIV-1 replication and/or the antiviral immunity remains to be clarified.
The function of m6A modifications within the HIV-1 RNA also remains unclear. Both the untranslated and coding regions have been described to carry m6A modifications [103,104,106]. To address their function, most of the studies focus on analysing the effect on HIV-1 infection of overexpressing or silencing different enzymes of the m6A modification pathway, including writers, readers and erasers. While two studies assigned a positive role to the m6A modifications [103,104], others assigned a negative one [105,106,107]. Although the reason of these contradictory results remains unclear, they might be explained by differences in methodology, cell types used or in the specific m6A modification pathway enzymes studied. Importantly, one of the studies addressed the role of specific m6A modifications. They showed that m6A modification of a highly conserved adenosine in the stem loop II region of HIV-1 Rev Response Element (RREIIB) promotes nuclear export and viral replication [103]. These results supported, at least for one specific location, a positive role of m6A modification in the HIV-1 life cycle. However, a recent study challenged the functional role of this modified adenosine. Using NMR analysis and a fluorescence polarization binding assay, it was shown that this m6A site has little effect on the structure, or on the binding affinity of RREIIB for the Rev arginine-rich-motif (ARM) in vitro [126]. Moreover, none of the other studies mapping m6A modifications in HIV-1 have observed this m6A site. Future sequencing studies at single-base resolution or independent of m6A antibodies may help in clarifying this discrepancy. Remarkably, recent work showed that YTHDF3 proteins are incorporated into HIV-1 particles in a nucleocapsid-dependent manner [107]. Once incorporated, YTHDF3 proteins limit infection in the new target cell, specifically, at the reverse transcription step. In turn, HIV-1 proteases within the virion cleave YTHDF3 proteins to ensure optimal infectivity of the mature virion.
In conclusion, current data support both negative and positive roles for m6A modifications and the m6A modification machinery in the HIV-1 life cycle. Further work will be required to complete our understanding of this novel HIV-1-host interaction.
4.2. m6A and the Flaviviridae Family
All positive-RNA viruses, including the members of the Flaviviridae family, replicate exclusively in the cytosol. Moreover, in contrast to the genomes of the other viral groups, the single-stranded positive-sense RNA genomes are highly structured and have a triple function. They act as mRNAs to translate the viral proteins, as templates for replication and as genomes for encapsidation. The regulation of these three functions occurs via specific RNA structures and binding of proteins that we do not completely understand yet.
Multiple m6A modifications have been detected by m6A-seq in the genome of DENV, ZIKV, WNV, YFV, and HCV. The highest density of m6A modifications is located in the coding regions of the RNA-dependent RNA-polymerase (NS5) and the protease (NS3) [67]. In HCV, 19 m6A peaks and 42 YTHDF-binding sites were identified by m6A-seq and PAR-CLIP, respectively. However, only two high-confidence YTHDF-binding sites overlapped with the m6A peaks identified by all m6A-seq replicates, suggesting that YTHDF readers might also interact with HCV RNA at non methylated sites. Silencing of the METTL3–METTL14 writers or YTHDF1-3 readers increased HCV virus production while silencing of the FTO eraser showed the opposite effect [67]. Further work demonstrated that this effect occurs at the step of virion production or release. Interestingly, analysis of the subcellular localization of YTHDF1-3 proteins by confocal microscopy revealed that these proteins colocalize with the HCV core protein around lipid-droplets, the site of HCV particle assembly [67]. This was the first observation of host m6A readers being recruited to specific viral compartments. Whether this is a common feature of all flaviviruses remains to be further explored. Finally, experiments abrogating four identified putative m6A modifications in the E1 gene of HCV RNA resulted in enhanced binding of the viral RNA to the HCV core protein by 2-fold compared to the wild type viral RNA. In addition, the mutated viral RNA produced 3-fold more infectious virions in the supernatant than the wild type virus. Taken together, the authors propose that m6A modifications in E1 are bound by YTHDF proteins to negatively regulate HCV packaging while the HCV core protein binds to unmodified viral RNA within the E1 region and facilitates packaging and virion exit.
As found for HCV, m6A modifications negatively regulate ZIKV infection [68]. m6A-seq identified twelve m6A peaks in the ZIKV RNA genome. Silencing of METTL3, METTL14 or YTHDF proteins increased ZIKV production while silencing ALKBH5 decreased it [68]. Overall, the studies in HCV and ZIKV uncovered an anti-viral role for the m6A modification in flavivirus RNA. Further investigations will be required to establish whether YTHDF proteins act to negatively regulate packaging in all members of the Flaviviridae, as observed for HCV. This raises the question of why m6A modifications are potentially conserved in flaviviruses RNA genomes. Plausible explanations include that m6A modification might regulate specific stages of flavivirus replication or the host antiviral response. Importantly, recent studies indicate that m6A modifications affect the innate immune sensing [113,127].
Infection by flaviviruses alters the m6A methylome of host mRNAs [28,68]. Some of these changes alter the expression of proteins that affect flavivirus infection such as RIOK3 or CIRBP [28]. Moreover, cellular pathways activated during viral infections, such as innate immunity and the ER stress response, also contributed to the changes in m6A modifications of host mRNAs [28]. Due to the small number of common cellular m6A peaks differentially methylated after infection in multiple members of the Flaviviridae family (DENV, ZIKV, WNV, and HCV), it will be of interest to address whether other RNA viruses from different families also share this manipulation of the m6A pathway.
4.3. m6A and Chikungunya Virus
The genome of the single-stranded positive-sense RNA virus Chikungunya (CHIKV) from the Togaviridae family also contains m6A modifications [111]. The authors developed a novel method named viral cross-linking and solid-phase purification (VIR–CLASP) to capture interactions between the pre-replicated viral genome and cellular proteins. In this technique, 4-thiouridine (4SU)-labeled virus is used to infect unlabelled cells followed by photocrosslinking, cell lysis and purification of the 4SU-labeled viral genomes and interacting crosslinked host proteins with Solid-Phase Reversible Immo-bilization (SPRI) beads. After nuclease digestion of the RNA, the identity of the crosslinked host proteins can be retrieved by mass spectrometry analysis. VIR–CLASP revealed that YTHDF2 and YTHDF3 interact with the CHIKV RNA. Moreover, m6A modifications were detected within the first 2000 nucleotides of the 5′-end of the CHIKV genome. Overexpression and depletion studies showed a pro-viral role for YTHDF2 and an anti-viral role for YTHDF1 and YTHDF3, a pattern similarly observed for HIV-1, however the molecular mechanisms behind these phenotypes requires further investigation. It is worth highlighting that CHIKV expresses the viral proteins from genomic and subgenomic viral RNAs. The 11.7 kB-long genomic RNA is partially translated to produce four non-structural proteins. In addition, CHIKV synthesises a 4.1 kB subgenomic RNA, identical to the 3’region of the genomic RNA, that gives rise to the structural proteins. Future studies mapping RNA modifications in the genomic and subgenomic RNAs will be necessary to establish whether they are differently modified to carry out distinct functions during the complex life cycle of these viruses.
4.4. m6A and Coronaviruses
The genome of coronaviruses consists of a single-stranded positive-sense RNA molecule. As CHIKV, coronaviruses also produce subgenomic RNAs. The first identification of m6A modifications in a coronavirus was carried out in the porcine epidemic diarrhea virus (PEDV) [108], shortly followed by SARS-CoV-2 [91]. PEDV does not infect humans but causes high mortality associated with severe diarrhea and vomiting in piglets younger than one week old. m6A-seq revealed that the PEDV genome isolated from purified viral particles contains seven m6A peaks, most of them located in ORF1b, which encodes nonstructural (NSP) proteins. Depletion of m6A writers, YTHDF1 or YTHDF2 increased PEDV replication while FTO depletion decreased it. Initial nanopore sequencing analyses suggested that m6A might not be one of the major modifications present in SARS-CoV-2 [91]; however, in a recent study which combined m6A-seq and miCLIP analyses, 8 m6A modifications were identified at single-base resolution in Vero cells [110]. A drawback of this study is that these experiments were carried out using fragmented total RNA, thus they do not allow to locate m6A sites to the subgenomic viral RNAs or the region of the genomic RNA that overlaps with these subgenomic RNAs. Depletion of METTL3 or METTL14 led to increased viral replication, while ALKBH5 knockdown had the opposite effect. Of the three YTHDF readers, only depletion of YTHDF2 affected viral replication, which was increased compared with control cells. Therefore, and as found for HCV, ZIKV and PEDV, m6A modifications negatively regulate SARS-CoV-2 infection.
4.5. m6A and Enterovirus 71
The positive-sense single-stranded enterovirus 71 (EV71) RNA genome contains clear m6A peaks detected by m6A-seq in the genes of the structural VP3 and VP1 and the helicase 2C [112]. Intriguingly, in contrast to that observed for other viral infections, expression and location of m6A writers, erasers and readers were altered upon EV71 infection, highlighting a powerful host epitranscriptomic manipulation by EV71. In Vero cells, depletion of METTL3, YTHDF2 and YTHDF3 decreased EV71 replication while FTO knockdown had the opposite effect. Moreover, single point mutation of m6A sites in VP1 or 2C resulted in decreased viral titers, indicating a clear positive role for m6A in EV71 infection [112]. However, in RD cells, depletion of YTHDF1-3 readers increased EV71 replication [112]. The reason of these divergent m6A-mediated responses to EV71 infection in different cells lines remains unknown. Interestingly, mass spectrometry analysis revealed an interaction between METTL3 and the viral RNA-dependent RNA polymerase (RdRp) 3D. Moreover, METTL3 enhanced sumoylation and ubiquitination of the RdRP 3D to facilitate viral replication. The physical and functional interaction of the writer METTL3 with the viral polymerase opens new perspectives in the interplay between the m6A machinery and the viral life cycle.
4.6. m6A and Influenza A Virus
IAV is known to contain m6A methylations since 1976 [96], however the location of these and their implications for viral replication remained elusive for decades. IAV contains a segmented genome of eight single-stranded RNA molecules of negative polarity (vRNAs). The IAV genome replicates in the nucleus and expresses the viral proteins in the cytoplasm via expression of RNAs of positive polarity that function as mRNAs. In 2017, a study using PA-m6A-seq mapped m6A modifications on both the plus (mRNAs) and minus (vRNAs) strands on multiple locations of HA, NP, M, NA and at lower levels on PB1, PB2 and PA viral segments [109]. METTL3 knockout caused an eight-fold reduction in expression of IAV genes which was accompanied by a significant reduction in the production of IAV virions. Additionally, overexpression of YTHDF2, but not YTHDF1 or YTHDF3, increased IAV gene expression and viral replication. PAR-CLIP sequencing showed a correlation between YTHDF1-3 binding sites and the viral m6A peaks in the positive and negative strands. Moreover, mutation of multiple m6A motifs in the HA gene reduced HA mRNA and protein expression without affecting the expression of other viral mRNA and proteins. Importantly, these viral mutants show reduced IAV pathogenicity in mice. Although the mutations did not affect the amino acid composition, the effect of mutating so many nucleotides at once in the viral RNA structure was not evaluated. This is especially important as m6A is known to regulate secondary RNA structure and RNA-binding protein interactions [62]. An additional remaining question is how YTHDF readers can access the m6A sites in the vRNA segments, which are tightly bound by nucleocapsid (NP) protein.
4.7. m6A and Respiratory Syncytial Virus
A recent study revealed with m6A-seq that the single-stranded negative sense RSV genomic RNA (gRNA), complementary RNA (cRNA) and the multiple viral RNA transcripts are internally m6A-modified in HeLa and A549 cells, with the majority of the m6A peaks conserved between the two cell lines [70]. The G viral transcript encoding the attachment glycoprotein present in the surface of the RSV virion was the most extensively modified of all 10 viral mRNA transcripts. m6A writers positively regulated RSV replication while m6A erasers showed the opposite effect. Overexpression of YTHDF1-3 proteins significantly increased RSV protein expression, gRNA, mRNA synthesis and virion production. Synonymous mutations of conserved m6A motifs under three m6A peaks in the G transcript showed reduced replication in A549 cells, primary human airway epithelial cultures and respiratory tracts of cotton rats. Together, these results consistently argue for a positive role of m6A in RSV infection. Moreover, RSV infection significantly altered the host epitranscriptom; however, in this case the effect was heavily dependent on the cell type analysed, with 2356 m6A peaks differentially methylated in HeLa cells and only 44 peaks in A549 cells. The reason of these differences between cell lines remains unknown. It might be plausible that differences in the host cell shut-off dynamics might affect the m6A peak calling.
4.8. m6A and Vesicular Stomatitis Virus
The vesicular stomatitis virus (VSV) is a negative-sense RNA virus from the Rhabdoviridae family. miCLIP experiments revealed that the VSV positive-sense RNAs contain 18 m6A sites [114]. During VSV infection, METTL3, but not METTL14, relocated from the nuclei to the cytoplasm where it modified VSV (+) RNAs and decreased viral dsRNA formation. Moreover, METTL3 knockdown increased the dsRNA levels on m6A sites of VSV RNA. Consequently, the ability of innate RNA sensors, such as RIG-I and MDA5, to bind viral dsRNA was suppressed and the activation of the innate antiviral response prevented. Interestingly, depletion of METTL3 enhanced type I IFN expression and protected mice against VSV infection. These findings highlight METTL3 as a potential target for antiviral intervention.
5. Epitranscriptomic Regulation of DNA Viruses
Most DNA viruses replicate in the nucleus and thus have access to the writing and erasing m6A machinery. Consistently, the mRNAs expressed from DNA viruses contain m6A modifications. As observed for RNA viruses, no global pro- or anti-viral role for viral m6A modifications can be generally ascribed to all DNA viruses studied to date (Table 2).
5.1. m6A and Simian Virus 40
In 1975 it was already reported that the mRNA from the simian virus 40 (SV40), a small DNA tumor virus from the Polyomaviridae family, was m6A-modified [94]; however, the functional importance of this observation was not elucidated until recently. Using PA-m6A-seq, two m6A clusters were mapped in the SV40 early region and 11 m6A clusters in the SV40 late region [115]. Additionally, PAR-CLIP revealed that most m6A clusters on SV40 transcripts are bound by YTHDF2 and YTHDF3. Overexpression of YTHDF2 increased SV40 replication, while knockout of YTHDF2 or METTL3 decreased SV40 replication, indicating a positive role for the m6A modification in SV40 replication. Notably, synonymous mutations of the 20 potential DRACH consensus sequences within the 11 m6A clusters present in the SV40 late region suggested a non-canonical mechanism of m6A modification. Only three of the 11 m6A clusters were entirely lost in the mutant virus (VPm), while six m6A clusters showed reduced m6A levels and two m6A clusters were unaffected. This argues that some m6A sites in viral RNA may be added independently of METTL3 and/or that m6A modifications may occur outside of the DRACH motif more frequently in viral RNA than in host mRNAs. Importantly, the VPm mutant virus replicated more slowly and produced smaller plaques than wild-type (WT) virus. When the VP1 ORF (which contains most of the SV40 m6A clusters) was expressed from a plasmid either in WT version or with the mutated m6A sites, a 10-fold lower protein expression was seen in the mutant VP1 compared with the WT, without a change in mRNA abundance or the encoded protein sequence, pointing to a deficient mRNA nuclear transport and/or translation.
5.2. m6A and Herpesviruses
A general feature of viruses within the Herpesviridae family is that they cause both lytic (acute) and latent infections. This adds another layer of complexity in the Herpesviridae family as two different types of viral transcripts are produced in latently or lytically infected cells. At large, m6A modifications regulate latent and lytic viral transcripts of members of the Herpesviridae family, as well as viral oncogenesis. Although initial reports carried out decades ago showed an internal m6A modification in herpes simplex virus type 1 (HSV-1) RNA [98], within the Herpesviridae family, the role of m6A in viral RNAs from herpesvirus-infected cells has only been extensively studied in the human oncogenic viruses Kaposi’s sarcoma-associated herpesvirus (KSHV) and Epstein-Barr virus (EBV).
In the latent stage, KSHV is dormantly present as an episome in the host nucleus and expresses only a few genes to sustain latency. Reactivation of KSHV induces the lytic infection which is triggered by the production of the viral master regulator ORF50 mRNA transcript that encodes the replication and transcription activator (RTA) protein. To date, three different groups have mapped m6A modifications in KSHV transcripts with the use of m6A-seq [59,102,117]. A comparative analysis of m6A peaks detected in both viral and cellular transcripts between all three studies showed an excellent overlap between m6A peaks, especially in KSHV mRNAs when the same cell type was compared [59].
The role of METTL3 and FTO has been evaluated by two independent groups in BCBL-1 cells, a primary effusion lymphoma B-cell line [59,116]. In both studies, FTO depletion increased m6A levels in KSHV mRNAs and enhanced lytic mRNA and protein expression, while METTL3 depletion had the opposite effect, indicating a pro-viral role for m6A during lytic KSHV infection in this cell line. In addition, YTHDF1, YTHDF2 or YTHDF3 depletion also decreased KSHV lytic replication in the same cell line [59]. In contrast, in TREx BCBL-1 cells, which are BCBL-1 cells that contain a Myc-tagged version of spliced viral RTA under the control of a doxycycline-inducible promoter, METTL3 and YTHDF2 knockdown resulted in an increase in RTA protein expression, however, despite the increase in this key viral lytic protein, virion production was not affected [102].
Regarding the role of YTHDF proteins in KSHV replication, contrasting results are also found even when studying the same cell type. Depletion of YTHDF2 in iSLK.BAC16 cells, a renal carcinoma cell line infected by recombinant KSHV BAC16, led to enhanced KSHV replication and increased half-lives of viral transcripts, suggesting an anti-viral role for YTHDF2, while the other YTHDF readers did not consistently affect KSHV replication [117]. However, depletion of METTL3 and YTHDF2 in iSLK.219 cells (iSLK cells infected with the KSHV strain rKSHV.219) decreased RTA protein levels and significantly reduced virion production [102], while the other YTHDF readers did not affect KSHV replication. The observation of both pro-viral and anti-viral roles for YTHDF2 in almost identical cell lines is thus again an issue to take into account.
Mapping of m6A sites in KSHV and host transcripts from different cell lines was carried out in an impressive m6A-seq study with five different KSHV latently infected cell lines and two cell lines undergoing lytic replication [117]. Moreover, the changes induced by KSHV in the cellular epitranscriptome were also studied in detail by comparing four pairs of uninfected and latently infected cell lines. Intriguingly, it was revealed that KSHV infection triggered 5′UTR hypomethylation and 3′-UTR hypermethylation of host mRNAs, especially in those transcripts involved in transformation pathways, suggesting that KSHV may alter the host epitranscriptomic landscape to induce cellular transformation. This study opened up the opportunity to identify the role of specific m6A modifications in cellular mRNAs in the bi-phasic KSHV life cycle, including assessing their role in oncogenic transformation pathways.
More recently, the use of a novel m6A peak-calling algorithm [128] facilitated the mapping of m6A modifications in KSHV mRNAs by m6A-seq in TREx-BCBL-1 cells [59]. To assess which proteins bind to an m6A-modified hairpin identified in open reading frame 50 (ORF50) mRNA, RNA affinity coupled to mass spectrometry analysis was performed with a methylated ORF50 RNA hairpin and an unmethylated control. Interestingly, in addition to YTH readers, the m6A-modified hairpin was specifically bound by several Tudor-containing proteins, including staphylococcal nuclease domain-containing protein 1 (SND1), all of which contain an aromatic cage structurally similar to the one found in the YTH domain [65,129]. RIP-seq confirmed a strong binding affinity of SND1 for the ORF50 mRNA and revealed a specific overlap between the RNA-binding sites of endogenous SND1 with cellular m6A peaks, uncovering SND1 as a novel m6A reader. Importantly, SND1 binding to ORF50 mRNA was dependent on the methylated status of this RNA and SND1 depletion led to a marked inhibition of KSHV early expression, including the ORF50 RNA. Moreover, another recent study also associated SND1 with m6A modification and showed that SND1 could alter m6A levels in colorectal cancer cell lines [130].
In summary, all KSHV m6A epitranscriptomic studies to date have reported m6A modifications in ORF50 RNA, highlighting the importance of this highly regulated RNA for KSHV lytic replication. Future experiments using long-read sequencing should help distinguish where the multiple m6A sites are located in both the spliced and unspliced ORF50 RNA and the exact role of m6A in this key transcript during lytic reactivation. Moreover, single-nucleotide mapping of m6A sites in both viral and cellular mRNAs during latent and lytic infection coupled to mutagenesis of specific m6A sites will allow further gain and loss of function studies to elucidate how m6A influences KSHV replication and how KSHV triggers the oncogenic pathway.
Another member of the Herpesviridae family was recently reported to harbour m6A modifications is Epstein–Barr virus (EBV) [118,119]). Using m6A-seq, two different groups mapped m6A modifications in the latent and lytic EBV mRNAs [118,119]. METTL14 depletion led to reduced stability of latent transcripts, whereas it increased stability of lytic transcripts [118]. Interestingly, METTL14 transcript and protein levels were dramatically increased in EBV latently infected cells. EBV latent antigen EBNA3C positively regulated METTL14 mRNA expression by activating the METTL14 promoter and also stabilising METTL14 protein. Moreover, METTL14 was upregulated in EBV-positive tumors and co-localised with EBNA3C. Notably, knockdown of METTL14 or EBNA3C in NOD-SCID mice led to a dramatic slowing of tumor growth compared to the control group. Overall, these results point at METTL14 as an essential factor in EBV replication and oncogenesis representing an innovative therapeutic target to treat EBV-positive tumors. Another team observed that out of all the m6A-related proteins, YTHDF1 depletion promoted EBV infection the most [119]. Mechanistically, YTHDF1 binds the viral lytic transcripts BZLF1 and BRLF1, and YTHDF1 knockdown increased their half-lives [119].
5.3. m6A and Adenovirus
The nuclear-replicating adenoviral RNAs were reported to bear m6A modifications in the 1970s [95]; however, it was only recently found that by using direct RNA long-read sequencing these modifications were located at the nucleotide- and at the isoform-level [93]. As many adenoviral RNAs have overlapping regions, m6A-seq failed to accurately map viral m6A modifications. Instead, with the combination of wild type and METTL3 KO A549 cell lines and direct RNA sequencing, a total of 97 m6A sites were elucidated in the adenoviral serotype 5 (Ad5) RNAs. While depletion of YTHDF1-3 readers or m6A erasers did not affect viral replication, METTL3 or METTL14 knockdown reduced viral replication. Specifically, METTL3 knockdown negatively impacted the splicing and accumulation of late adenoviral RNAs [93].
5.4. m6A and Hepatitis B Virus
The hepatitis B virus (HBV) genome consists of a partially double-stranded molecule of circular DNA that localises into the nucleus. There, HBV DNA transcribes the viral mRNA and pregenomic RNA (pgRNA), which are exported to the cytosol to express the viral proteins. The pgRNA is also used in the cytosol as a template for reverse transcription to generate new molecules of HBV genomes. Recently, m6A modifications were detected in HBV mRNAs and pgRNA from HBV-expressing cells and from liver tissues of chronic HBV patients [120]. Depletion of writers YTHDF2 and YTHDF3 increased the expression of the HBV proteins HBc and HBs, whereas FTO or ALKBH5 depletion had the opposite effect, indicating a negative effect of the m6A machinery on HBV expression. Depletion of METTL3 and METTL14 writers or of the reader YTHDF2 significantly extended by ≈10 h the pgRNA half-life, indicating that m6A modification significantly lowers the stability of HBV RNA transcripts. Moreover, depletion of writers inhibited reverse transcription of HBV pgRNA. A conserved m6A consensus motif bearing an m6A modification was identified by m6A-seq in the epsilon stem loop structure at the 3′-end of all HBV transcripts and at both the 5′-end and 3′-end epsilon stem loops of pgRNA. Using different single point A to C mutant pgRNAs, either lacking m6A modifications at the 5′-end, 3′-end or both, revealed that the effect of m6A modification depends on the location of m6A in the pgRNA. Indeed, m6A methylation at the 5′ end of the pgRNA facilitated reverse transcription of pgRNA, while the m6A at the 3′ end made HBV RNAs less stable. How m6A modification modulates reverse transcription of HBV pgRNA is currently unknown but it is possible that YTHDF readers are involved in this modulation as seen for HIV-1.
6. Epitranscriptomic Regulation of the Immune Response to Viral Infection
The m6A RNA modification affects the innate and adaptive immune responses. Innate immunity to viral infections heavily relies on the type I IFN response in which foreign viral RNA is recognised by the pathogen recognition receptors (PRR). These include the toll-like receptors (TLR) 3, 7 and 9 in endosomes and the RIG-I like receptors (RLR) in the cytosol of which the most important include the retinoic acid-inducible gene-I (RIG-I) and the melanoma differentiation-associated protein 5 (MDA5). Triggering of endosomal or cytosolic RNA sensors induces a signaling cascade leading to the expression and secretion of IFNα and IFNβ that are recognised by the interferon receptor (IFNAR). This results in the activation of the JAK–STAT pathway and the subsequent transcription of hundreds of interferon stimulated genes (ISGs) that mediate an antiviral response [131].
Viruses have evolved to escape or inhibit the type I IFN response by multiple mechanisms. These include the m6A methylation of viral RNAs to prevent recognition by the innate RNA sensor RIG-I [113,127]. For example, m6A-deficient HMPV RNA facilitated the RIG-I conformational change necessary to enhance downstream IFN signaling [113]. Infection of cotton rats with mutant HMPV viruses lacking multiple m6A sites induced significantly higher type I IFN responses in cotton rats than the parental HMPV viruses and resulted in attenuated infection [113]. Moreover, following depletion of METTL3 or YTHDF2 proteins, the mRNA expression of IFNβ and various ISGs was enhanced in human and murine cytomegalovirus- (HCMV and MCMV), IAV-, adenovirus- or vesicular stomatitis virus (VSV)-infected cells [121]. Similarly, YTHDF2 and YTHDF3 silencing led to an increase in IFNβ mRNA levels in cells transfected with HCV RNA carrying a mutation that abrogates viral replication [127]. Moreover, the IFNβ levels were unaffected when cells were transfected with the same HCV RNA that contained a mutation that abrogated an m6A modification near a pathogen-associated molecular pattern (PAMP) present in the HCV RNA. Consistently, METTL14 depletion resulted in significantly elevated IFNβ mRNA levels in HCMV-infected cells and uninfected cells exposed to dsDNA, while ALKBH5 depletion reduced IFNβ mRNA levels [124]. Relevantly, the IFNβ mRNA has three m6A sites whose abolition stabilises IFNβ mRNA, which in turn sustains prolonged expression of the cytokine IFNβ, resulting in a stronger antiviral response [121]. YTHDF3 has also been reported to negatively regulate anti-viral immunity through specific suppression of ISGs expression and without affecting IFNβ mRNA levels [123]. Mechanistically, YTHDF3 promotes translation of the transcription repressor forkhead box protein O3 (FOXO3), which is known to negatively regulate ISG expression. Consequently, silencing of YTHDF3 in macrophages inhibited viral infection with several DNA and RNA viruses, which included VSV, encephalomyocarditis virus (EMCV) and herpes simplex virus type 1 (HSV-1). Importantly, YTHDF3-/- mice were protected against infection with VSV. Recently, it was also found that in response to VSV infection, host cells impair the enzymatic activity of the m6A eraser ALKBH5 by inducing R107 demethylation on the ALKBH5 protein [122]. Due to decreased ALKBH5 activity, m6A methylation is increased in the α-ketoglutarate dehydrogenase (OGDH) transcript. This leads to reduced OGDH mRNA and protein expression. As OGDH is an enzyme that produces the metabolite itaconate, required for viral replication, viral replication is inhibited without the involvement of the innate immune system.
Collectively, these data show an exciting interplay between viruses-m6A modifications and immune responses that might allow for novel therapeutic interventions.
7. Conclusions and Future Perspectives
Most of our current knowledge of m6A modifications in viral RNAs have emerged in the past five years, consequently we are only beginning to understand the exact abundance and role of this modification during viral replication. Currently, several high-throughput deep-sequencing m6A mapping techniques at single-nucleotide resolution exist; however, most viral studies to date have relied on m6A-seq to locate m6A in viral RNA, with the exception of the use of PA-m6A-seq for IAV, HIV-1, SV40 and EBV, miCLIP for VSV and SARS-CoV2, and the use of nanopore sequencing for adenovirus. Because the resolution of m6A-seq is low (100–200 nucleotides) and m6A can appear in clusters, which results in overlapping m6A peaks, peak-based prediction algorithms miss more than half of m6A sites occurring in these clusters [39]. Clearly, the use of single-nucleotide resolution mapping studies has to be the next step forward in understanding m6A modifications in viral RNA. Moreover, whether the identified m6A peaks in viral RNAs contain DRACH motifs or whether these modifications are METTL3-dependent, the major catalyser of m6A addition in host mRNAs remains to be clarified for the majority of viruses investigated to date. This is of particular importance because (i) when DRACH motifs are abrogated in viral RNAs, only a reduction of m6A levels is generally observed [70,115] and (ii) other host m6A-methyltransferases such as METTL16, METTL5 and ZCCHC4 have been recently identified. METTL16 targets specific host pre-mRNAs, mRNAs, snRNAs and lncRNAs [34]. METTL5 and ZCCHC4 exclusively methylate a single m6A site present in the 18S rRNA and in the 28S rRNA, respectively [132,133]. Therefore, it is feasible that viral RNAs might be methylated by enzymes other than METTL3, especially in viruses with non-canonical mRNAs, such as the very long and highly structured RNAs from flaviviruses and coronaviruses.
Abrogation of m6A in viral RNAs led to reduced viral replication in the vast majority of viruses investigated, indicating a general positive role for m6A in viral RNA. However, it is important to note that only one study in HBV [120] confirmed that the resulting effect on viral replication was a sole consequence of the loss of m6A and not a consequence of altering the viral RNA secondary structure after mutating A to C in the DRACH motif. This is particularly important when several mutations are introduced at once and it should be addressed to gather more solid evidence of the role of m6A modification. m6A mapping at single-nucleotide resolution will allow pinpointing specific sites to mutate instead of mutating all DRACH sites under a given m6A peak. Now that some m6A sites have been accurately located in viral RNAs, another step forward would be understanding the putative role of m6A in regulating viral RNA secondary structures and elucidating which m6A readers bind to these sites. Another critical question is whether m6A sites act redundantly in a single transcript, especially in those heavily m6A-modified viral RNAs, and whether abrogation of multiple sites is actually required to see a clear effect on viral infection.
The role of the YTHDF readers on viral replication has been extensively evaluated with depletion and overexpression studies in multiple viruses. Both YTHDF pro-viral and anti-viral functions have been reported depending on the virus and cell type used. While initial papers described different functions for each reader, with YTHDF1 enhancing translation of m6A-modified RNAs [49], YTHDF2 promoting m6A-modified mRNA degradation [51] and YTHDF3 enhancing both translation and mRNA degradation of methylated transcripts [47], these reader functions cannot always be directly translated to viral RNAs. For example, for some viruses YTHDF2 has been shown to mediate viral mRNA degradation [117,120], while in contrast, in other viruses overexpression of YTHDF2 increased viral RNA levels [111]. Moreover, a new model proposes that YTHDF1-3 readers act redundantly to mediate m6A-modified mRNA degradation [48]. Although not all viral studies have always evaluated the role of the three different YTHDF readers, it is now clear that YTHDF proteins do not always act redundantly and they clearly exhibit different phenotypes in viral infections. This has been observed in HIV-1, CHIKV, IAV and KSHV, suggesting that although the human C-terminus YTH domain is highly conserved between these proteins [134], the less conserved N-terminus differs and may add an extra layer of regulation that discriminate the role of these proteins on viral RNAs. Moreover, it is important to realise that depletion and overexpression of the m6A machinery has a broad effect on cellular homeostasis by affecting host RNA biology. Therefore, any effect on viral replication as a consequence of depleting m6A writers, erasers and readers could be an indirect effect from the depletion of crucial players in regulating RNA fates. For example, despite ALKBH5 depletion resulting in marked inhibition of VSV, m6A-seq analysis performed on WT and ALKBH5-deficient macrophages revealed that there was no up-regulation of m6A signal on VSV RNA. Thus, ALKBH5 promotes VSV replication not by targeting m6A modification of VSV RNA but by directly targeting m6A modification on the host OGDH mRNA to up-regulate the OGDH-itaconate metabolic pathway [122]. Finally, besides YTHDF proteins, other m6A readers could also play important roles in viral infections as recently shown for KSHV and the SND1 protein [59], hence, further RNA affinity experiments will be of great interest to elucidate the viral m6A-interactome.
To date, very few studies have shed light on how virus-induced changes in the host m6A landscape impact the viral replication cycle. Intriguingly, despite viral infections causing major perturbations in the cell at multiple levels, the virus-induced remodeling of the host epitranscriptome seems to occur at a limited amount of m6A sites [28,59]. Another important aspect to be explored is whether particular m6A sites in viral RNA can be subjected to demethylation during a viral infection cycle, and whether FTO or ALKBH5 may play a role in this process. Moreover, most studies have been carried out in transformed cell lines. Therefore, further epitranscriptomic studies using relevant primary cell lines and in vivo studies are essential. Most viruses infect a narrow spectrum of related hosts or cell types. Important exceptions are the emerging mosquito-borne viruses such as DENV, WNV, ZIKV and CHIKV that efficiently infect both humans and mosquitoes; two organisms separated by around one billion years of evolution. Whether the viral RNAs are differentially modified in both hosts and whether the arthropod m6A landscape is also altered under viral infection conditions is unexplored. However, under bacterial infection conditions, a recent study in Drosophila melanogaster flies showed that infection with the bacteria Wolbachia pipientis conveys protection against RNA virus infection by upregulation of the D.melanogaster methyltransferase MT2 [135]. MT2 is a m5C methyltransferase that catalyses a methylation to the fifth carbon on a cytosine base in tRNAAsp. Wolbachia-infected flies have a seven to eight-fold increase in MT2 transcript levels and repress Sindbis virus (SINV) infection. On the other hand, MT2 knockout flies are more susceptible to SINV infection than wild-type flies. Together, this study suggests an anti-viral function for m5C methylation in flies and argues for a possible role for m6A methylations in regulating virus infection in other insect vectors. It would thus be of great interest to study the interplay of m6A/mosquito-borne viruses in mosquitoes and to compare with that in humans. Putative differences might help understanding why and how these viruses cause an acute infection in human cells and a chronic one in mosquito cells.
The importance of the epitranscriptome in regulating viral replication and antiviral immunity opens the door for novel therapeutic interventions that target either the host or viral epitranscriptome to restrict viral replication. Our current knowledge supports that m6A modifications might act in a pro-viral or anti-viral manner depending on the virus, cell type analysed, and even on its location in the viral RNA molecule. Thus, an approach that specifically targets key m6A sites in the viral RNAs seems ideal, although quite challenging. A more general approach that enhances m6A methylations with specific compounds [136] might be of interest to treat viruses negatively regulated by them. However, these treatments will also affect the host RNAs. Besides drugging the viral epitranscriptome, generation of viral strains with altered m6A methylations might be useful for vaccine purposes because m6A modifications have been implicated in the antiviral immune response [18] and m6A-depleted viruses are known to induce potent type I IFN responses in vivo [113]. Although the functionality of such a vaccine has not been evaluated yet in vivo, it seems plausible that infection with live attenuated vaccines based on m6A-depleted viruses might induce potent adaptive immune responses providing the host with long lasting immunity to wild-type viruses. In line with this, RSV viruses expressing G transcripts depleted of m6A modification and m6A-deficient HMPV viruses were highly attenuated yet retained high immunogenicity in cotton rats [70,113], while IAV viruses depleted of m6A modification in HA resulted in reduced IAV pathogenicity in mice [109].
Recent advances point out the host m6A machinery as a key interactor in viral infections. Advancing in this novel virus-host interplay is of great interest not only in virology but also in cell biology as viruses have proven to be great tools to uncover novel aspects of multiple cellular processes. In conclusion, the journey of deciphering the viral m6A epitranscriptome, which started in the 1970s, has truly bloomed in the past five years and pointed to the host m6A machinery as being an important component in viral infections. However, to go beyond descriptive observations and derive a mechanistic understanding of viral RNA modifications and their benefits for virus survival requires further efforts. The recent m6A sequencing technologies will clearly be a major help for this endeavor.
Funding
This work was supported by the Spanish Ministry of Science and Innovation through grants PID2019-106959RB-I00/AEI/10.13039/501100011033, an institutional “Maria de Maeztu” Programme for Units of Excellence in R&D (CEX2018-000792-M), by the 2017 SGR 909 grant from the Secretaria d’Universitats i Recerca del Departament d’Economia i Coneixement of the Generalitat de Catalunya and a Beatriu Pinós postdoctoral fellowship awarded to Belinda Baquero-Perez which is funded by the Secretary of Universities and Research (Government of Catalonia) and by the Horizon 2020 programme of research and innovation of the European Union under the Marie Sklodowska-Curie grant agreement No 801370. All figures in this review were drawn using BioRender.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Davis, F.F.; Allen, F.W. Ribonucleic acids from yeast which contain a fifth nucleotide. J. Biol. Chem. 1957, 227, 907–915. [Google Scholar] [CrossRef]
- Zaccara, S.; Ries, R.J.; Jaffrey, S.R. Reading, writing and erasing mRNA methylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 608–624. [Google Scholar] [CrossRef]
- Desrosiers, R.; Friderici, K.; Rottman, F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc. Natl. Acad. Sci. USA 1974, 71, 3971–3975. [Google Scholar] [CrossRef] [Green Version]
- Perry, R.P.; Kelley, D.E. Existence of methylated messenger RNA in mouse L cells. Cell 1974, 1, 37–42. [Google Scholar] [CrossRef]
- Amos, H.; Korn, M. 5-Methyl cytosine in the RNA of Escherichia coli. Biochim. Biophys. Acta 1958, 29, 444–445. [Google Scholar] [CrossRef]
- Baskin, F.; Dekker, C.A. A Rapid and Specific Assay for Sugar Methylation in Ribonucleic Acid. J. Biol. Chem. 1967, 242, 5447–5449. [Google Scholar] [CrossRef]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.-G.; et al. N6-Methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef]
- Zheng, G.; Dahl John, A.; Niu, Y.; Fedorcsak, P.; Huang, C.-M.; Li Charles, J.; Vågbø Cathrine, B.; Shi, Y.; Wang, W.-L.; Song, S.-H.; et al. ALKBH5 Is a Mammalian RNA Demethylase that Impacts RNA Metabolism and Mouse Fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Motorin, Y.; Helm, M. RNA nucleotide methylation. Wires RNA 2011, 2, 611–631. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, C.; Limbach, P.A. Mass spectrometry of modified RNAs: Recent developments. Analyst 2016, 141, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xiong, X.; Yi, C. Epitranscriptome sequencing technologies: Decoding RNA modifications. Nat. Methods 2017, 14, 23–31. [Google Scholar] [CrossRef]
- Garalde, D.R.; Snell, E.A.; Jachimowicz, D.; Sipos, B.; Lloyd, J.H.; Bruce, M.; Pantic, N.; Admassu, T.; James, P.; Warland, A.; et al. Highly parallel direct RNA sequencing on an array of nanopores. Nat. Methods 2018, 15, 201–206. [Google Scholar] [CrossRef]
- Jonkhout, N.; Tran, J.; Smith, M.A.; Schonrock, N.; Mattick, J.S.; Novoa, E.M. The RNA modification landscape in human disease. RNA 2017, 23, 1754–1769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA Modifications in Gene Expression Regulation. Cell 2017, 169, 1187–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, J.V.; Cordero-Espinoza, L.; Oeztuerk-Winder, F.; Andersson-Rolf, A.; Selmi, T.; Blanco, S.; Tailor, J.; Dietmann, S.; Frye, M. Cytosine-5 RNA Methylation Regulates Neural Stem Cell Differentiation and Motility. Stem. Cell Rep. 2017, 8, 112–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livneh, I.; Moshitch-Moshkovitz, S.; Amariglio, N.; Rechavi, G.; Dominissini, D. The m6A epitranscriptome: Transcriptome plasticity in brain development and function. Nat. Rev. Neurosci. 2020, 21, 36–51. [Google Scholar] [CrossRef]
- Batista Pedro, J.; Molinie, B.; Wang, J.; Qu, K.; Zhang, J.; Li, L.; Bouley Donna, M.; Lujan, E.; Haddad, B.; Daneshvar, K.; et al. m6A RNA Modification Controls Cell Fate Transition in Mammalian Embryonic Stem Cells. Cell Stem. Cell 2014, 15, 707–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shulman, Z.; Stern-Ginossar, N. The RNA modification N6-methyladenosine as a novel regulator of the immune system. Nat. Immunol. 2020, 21, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Fustin, J.-M.; Doi, M.; Yamaguchi, Y.; Hida, H.; Nishimura, S.; Yoshida, M.; Isagawa, T.; Morioka Masaki, S.; Kakeya, H.; Manabe, I.; et al. RNA-Methylation-Dependent RNA Processing Controls the Speed of the Circadian Clock. Cell 2013, 155, 793–806. [Google Scholar] [CrossRef] [Green Version]
- Bohnsack, K.E.; Höbartner, C.; Bohnsack, M.T. Eukaryotic 5-methylcytosine (m⁵C) RNA Methyltransferases: Mechanisms, Cellular Functions, and Links to Disease. Genes 2019, 10, 102. [Google Scholar] [CrossRef] [Green Version]
- Dai, D.; Wang, H.; Zhu, L.; Jin, H.; Wang, X. N6-methyladenosine links RNA metabolism to cancer progression. Cell Death Dis. 2018, 9, 124. [Google Scholar] [CrossRef] [Green Version]
- Dimitrova, D.G.; Teysset, L.; Carré, C. RNA 2′-O-Methylation (Nm) Modification in Human Diseases. Genes 2019, 10, 117. [Google Scholar] [CrossRef] [Green Version]
- Kadumuri, R.V.; Janga, S.C. Epitranscriptomic Code and Its Alterations in Human Disease. Trends Mol. Med. 2018, 24, 886–903. [Google Scholar] [CrossRef] [PubMed]
- Gonzales-van Horn, S.R.; Sarnow, P. Making the Mark: The Role of Adenosine Modifications in the Life Cycle of RNA Viruses. Cell Host Microbe 2017, 21, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, E.M.; Courtney, D.G.; Tsai, K.; Cullen, B.R. Viral Epitranscriptomics. J. Virol. 2017, 91, e02263-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netzband, R.; Pager, C.T. Epitranscriptomic marks: Emerging modulators of RNA virus gene expression. Wiley Interdiscip. Rev. RNA 2020, 11, e1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, B.; Gao, S.-J. RNA epitranscriptomics: Regulation of infection of RNA and DNA viruses by N(6) -methyladenosine (m(6) A). Rev. Med. Virol. 2018, 28, e1983. [Google Scholar] [CrossRef] [Green Version]
- Gokhale, N.S.; McIntyre, A.B.R.; Mattocks, M.D.; Holley, C.L.; Lazear, H.M.; Mason, C.E.; Horner, S.M. Altered m6A Modification of Specific Cellular Transcripts Affects Flaviviridae Infection. Mol. Cell 2020, 77, 542–555.e8. [Google Scholar] [CrossRef] [PubMed]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive analysis of mRNA methylation reveals enrichment in 3’ UTRs and near stop codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [Green Version]
- Fazi, F.; Fatica, A. Interplay Between N6-Methyladenosine (m6A) and Non-coding RNAs in Cell Development and Cancer. Front. Cell Dev. Biol. 2019, 7, 116. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Zheng, G.; Ning, Q.; Zheng, J.; Dong, D. Translation and functional roles of circular RNAs in human cancer. Mol. Cancer 2020, 19, 30. [Google Scholar] [CrossRef]
- Alarcón, C.R.; Goodarzi, H.; Lee, H.; Liu, X.; Tavazoie, S.; Tavazoie, S.F. HNRNPA2B1 Is a Mediator of m6A-Dependent Nuclear RNA Processing Events. Cell 2015, 162, 1299–1308. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, K.E.; Chen, B.; Liu, K.; Hunter, O.V.; Xie, Y.; Tu, B.P.; Conrad, N.K. The U6 snRNA m(6)A Methyltransferase METTL16 Regulates SAM Synthetase Intron Retention. Cell 2017, 169, 824–835.e14. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3–METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Feng, J.; Xue, Y.; Guan, Z.; Zhang, D.; Liu, Z.; Gong, Z.; Wang, Q.; Huang, J.; Tang, C.; et al. Structural basis of N6-adenosine methylation by the METTL3–METTL14 complex. Nature 2016, 534, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Geula, S.; Moshitch-Moshkovitz, S.; Dominissini, D.; Mansour, A.A.; Kol, N.; Salmon-Divon, M.; Hershkovitz, V.; Peer, E.; Mor, N.; Manor, Y.S.; et al. m6A mRNA methylation facilitates resolution of naïve pluripotency toward differentiation. Science 2015, 347, 1002. [Google Scholar] [CrossRef] [PubMed]
- Ping, X.-L.; Sun, B.-F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.-J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.-S.; et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linder, B.; Grozhik, A.V.; Olarerin-George, A.O.; Meydan, C.; Mason, C.E.; Jaffrey, S.R. Single-nucleotide-resolution mapping of m6A and m6Am throughout the transcriptome. Nat. Methods 2015, 12, 767–772. [Google Scholar] [CrossRef]
- Mauer, J.; Jaffrey, S.R. FTO, m6Am, and the hypothesis of reversible epitranscriptomic mRNA modifications. FEBS Lett. 2018, 592, 2012–2022. [Google Scholar] [CrossRef] [Green Version]
- Mauer, J.; Luo, X.; Blanjoie, A.; Jiao, X.; Grozhik, A.V.; Patil, D.P.; Linder, B.; Pickering, B.F.; Vasseur, J.J.; Chen, Q.; et al. Reversible methylation of m6Am in the 5’ cap controls mRNA stability. Nature 2017, 541, 371–375. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.; Sun, H.; Xu, C. YTH Domain: A Family of N6-methyladenosine (m6A) Readers. Genom. Proteom. Bioinform. 2018, 16, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liu, K.; Ahmed, H.; Loppnau, P.; Schapira, M.; Min, J. Structural Basis for the Discriminative Recognition of N6-Methyladenosine RNA by the Human YT521-B Homology Domain Family of Proteins. J. Biol. Chem. 2015, 290, 24902–24913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Wang, X.; Liu, K.; Roundtree, I.A.; Tempel, W.; Li, Y.; Lu, Z.; He, C.; Min, J. Structural basis for selective binding of m6A RNA by the YTHDC1 YTH domain. Nat. Chem. Biol. 2014, 10, 927–929. [Google Scholar] [CrossRef]
- Dai, X.; Wang, T.; Gonzalez, G.; Wang, Y. Identification of YTH Domain-Containing Proteins as the Readers for N1-Methyladenosine in RNA. Anal. Chem. 2018, 90, 6380–6384. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Zhao, Y.; He, J.; Zhang, Y.; Xi, H.; Liu, M.; Ma, J.; Wu, L. YTHDF2 destabilizes m6A-containing RNA through direct recruitment of the CCR4–NOT deadenylase complex. Nat. Commun. 2016, 7, 12626. [Google Scholar] [CrossRef]
- Shi, H.; Wang, X.; Lu, Z.; Zhao, B.S.; Ma, H.; Hsu, P.J.; Liu, C.; He, C. YTHDF3 facilitates translation and decay of N6-methyladenosine-modified RNA. Cell Res. 2017, 27, 315–328. [Google Scholar] [CrossRef] [Green Version]
- Zaccara, S.; Jaffrey, S.R. A Unified Model for the Function of YTHDF Proteins in Regulating m6A-Modified mRNA. Cell 2020, 181, 1582–1595.e8. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, B.S.; Roundtree, I.A.; Lu, Z.; Han, D.; Ma, H.; Weng, X.; Chen, K.; Shi, H.; He, C. N6-methyladenosine Modulates Messenger RNA Translation Efficiency. Cell 2015, 161, 1388–1399. [Google Scholar] [CrossRef] [Green Version]
- Meyer Kate, D.; Patil Deepak, P.; Zhou, J.; Zinoviev, A.; Skabkin Maxim, A.; Elemento, O.; Pestova Tatyana, V.; Qian, S.-B.; Jaffrey Samie, R. 5′ UTR m6A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Chen, Y.-S.; Ping, X.-L.; Yang, X.; Xiao, W.; Yang, Y.; Sun, H.-Y.; Zhu, Q.; Baidya, P.; Wang, X.; et al. Cytoplasmic m6A reader YTHDF3 promotes mRNA translation. Cell Res. 2017, 27, 444–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, W.; Adhikari, S.; Dahal, U.; Chen, Y.-S.; Hao, Y.-J.; Sun, B.-F.; Sun, H.-Y.; Li, A.; Ping, X.-L.; Lai, W.-Y.; et al. Nuclear m6A Reader YTHDC1 Regulates mRNA Splicing. Mol. Cell 2016, 61, 507–519. [Google Scholar] [CrossRef] [Green Version]
- Roundtree, I.A.; Luo, G.Z.; Zhang, Z.; Wang, X.; Zhou, T.; Cui, Y.; Sha, J.; Huang, X.; Guerrero, L.; Xie, P.; et al. YTHDC1 mediates nuclear export of N(6)-methyladenosine methylated mRNAs. Elife 2017, 6, e31311. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Dong, L.; Liu, X.-M.; Guo, J.; Ma, H.; Shen, B.; Qian, S.-B. m6A in mRNA coding regions promotes translation via the RNA helicase-containing YTHDC2. Nat. Commun. 2019, 10, 5332. [Google Scholar] [CrossRef]
- Bailey, A.S.; Batista, P.J.; Gold, R.S.; Chen, Y.G.; de Rooij, D.G.; Chang, H.Y.; Fuller, M.T. The conserved RNA helicase YTHDC2 regulates the transition from proliferation to differentiation in the germline. Elife 2017, 6, e26116. [Google Scholar] [CrossRef]
- Hsu, P.J.; Zhu, Y.; Ma, H.; Guo, Y.; Shi, X.; Liu, Y.; Qi, M.; Lu, Z.; Shi, H.; Wang, J.; et al. Ythdc2 is an N6-methyladenosine binding protein that regulates mammalian spermatogenesis. Cell Res. 2017, 27, 1115–1127. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef]
- Baquero-Perez, B.; Antanaviciute, A.; Yonchev, I.D.; Carr, I.M.; Wilson, S.A.; Whitehouse, A. The Tudor SND1 protein is an m6A RNA reader essential for replication of Kaposi’s sarcoma-associated herpesvirus. Elife 2019, 8, e47261. [Google Scholar] [CrossRef]
- Edupuganti, R.R.; Geiger, S.; Lindeboom, R.G.H.; Shi, H.; Hsu, P.J.; Lu, Z.; Wang, S.-Y.; Baltissen, M.P.A.; Jansen, P.W.T.C.; Rossa, M.; et al. N6-methyladenosine (m6A) recruits and repels proteins to regulate mRNA homeostasis. Nat. Struct. Mol. Biol. 2017, 24, 870–878. [Google Scholar] [CrossRef]
- Sun, L.; Fazal, F.M.; Li, P.; Broughton, J.P.; Lee, B.; Tang, L.; Huang, W.; Kool, E.T.; Chang, H.Y.; Zhang, Q.C. RNA structure maps across mammalian cellular compartments. Nat. Struct. Mol. Biol. 2019, 26, 322–330. [Google Scholar] [CrossRef]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N6-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Zhou, K.I.; Parisien, M.; Dai, Q.; Diatchenko, L.; Pan, T. N6-methyladenosine alters RNA structure to regulate binding of a low-complexity protein. Nucleic Acids Res. 2017, 45, 6051–6063. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.J.; Shi, H.; Zhu, A.C.; Lu, Z.; Miller, N.; Edens, B.M.; Ma, Y.C.; He, C. The RNA-binding protein FMRP facilitates the nuclear export of N6-methyladenosine–containing mRNAs. J. Biol. Chem. 2019, 294, 19889–19895. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Tong, L. Molecular basis for the recognition of methylated adenines in RNA by the eukaryotic YTH domain. Proc. Natl. Acad. Sci. USA 2014, 111, 13834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Su, S.; Patil, D.P.; Liu, H.; Gan, J.; Jaffrey, S.R.; Ma, J. Molecular basis for the specific and multivariant recognitions of RNA substrates by human hnRNP A2/B1. Nat. Commun. 2018, 9, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gokhale, N.S.; McIntyre, A.B.R.; McFadden, M.J.; Roder, A.E.; Kennedy, E.M.; Gandara, J.A.; Hopcraft, S.E.; Quicke, K.M.; Vazquez, C.; Willer, J.; et al. N6-Methyladenosine in Flaviviridae Viral RNA Genomes Regulates Infection. Cell Host Microbe 2016, 20, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Lichinchi, G.; Zhao, B.S.; Wu, Y.; Lu, Z.; Qin, Y.; He, C.; Rana, T.M. Dynamics of Human and Viral RNA Methylation during Zika Virus Infection. Cell Host Microbe 2016, 20, 666–673. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Liu, F.; Lu, Z.; Fei, Q.; Ai, Y.; He, P.C.; Shi, H.; Cui, X.; Su, R.; Klungland, A.; et al. Differential m6A, m6Am, and m1A Demethylation Mediated by FTO in the Cell Nucleus and Cytoplasm. Mol. Cell 2018, 71, 973–985.e975. [Google Scholar] [CrossRef] [Green Version]
- Xue, M.; Zhao, B.S.; Zhang, Z.; Lu, M.; Harder, O.; Chen, P.; Lu, Z.; Li, A.; Ma, Y.; Xu, Y.; et al. Viral N6-methyladenosine upregulates replication and pathogenesis of human respiratory syncytial virus. Nat. Commun. 2019, 10, 4595. [Google Scholar] [CrossRef]
- Gulati, P.; Avezov, E.; Ma, M.; Antrobus, R.; Lehner, P.; O’Rahilly, S.; Yeo, G.S.H. Fat mass and obesity-related (FTO) shuttles between the nucleus and cytoplasm. Biosci. Rep. 2014, 34, e00144. [Google Scholar] [CrossRef] [PubMed]
- Lesbirel, S.; Viphakone, N.; Parker, M.; Parker, J.; Heath, C.; Sudbery, I.; Wilson, S.A. The m6A-methylase complex recruits TREX and regulates mRNA export. Sci. Rep. 2018, 8, 13827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Wan, J.; Gao, X.; Zhang, X.; Jaffrey, S.R.; Qian, S.-B. Dynamic m6A mRNA methylation directs translational control of heat shock response. Nature 2015, 526, 591–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, M.; Chelysheva, I.; Goebel, I.; Trenkner, T.; Zhou, J.; Mao, Y.; Ignatova, Z. Dynamic m6A methylation facilitates mRNA triaging to stress granules. Life Sci. Alliance 2018, 1, e201800113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, Y.; Toth, J.I.; Petroski, M.D.; Zhang, Z.; Zhao, J.C. N6-methyladenosine modification destabilizes developmental regulators in embryonic stem cells. Nat. Cell Biol. 2014, 16, 191–198. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, Y.; Sun, B.-F.; Shi, Y.; Yang, X.; Xiao, W.; Hao, Y.-J.; Ping, X.-L.; Chen, Y.-S.; Wang, W.-J.; et al. FTO-dependent demethylation of N6-methyladenosine regulates mRNA splicing and is required for adipogenesis. Cell Res. 2014, 24, 1403–1419. [Google Scholar] [CrossRef]
- He, L.; Li, H.; Wu, A.; Peng, Y.; Shu, G.; Yin, G. Functions of N6-methyladenosine and its role in cancer. Mol. Cancer 2019, 18, 176. [Google Scholar] [CrossRef] [Green Version]
- Paramasivam, A.; Priyadharsini, J.V.; Raghunandhakumar, S. Implications of m6A modification in autoimmune disorders. Cell. Mol. Immunol. 2020, 17, 550–551. [Google Scholar] [CrossRef]
- Noack, F.; Calegari, F. Epitranscriptomics: A New Regulatory Mechanism of Brain Development and Function. Front. Neurosci. 2018, 12, 85. [Google Scholar] [CrossRef]
- Manners, O.; Baquero-Perez, B.; Whitehouse, A. m6A: Widespread regulatory control in virus replication. Biochim. Et Biophys. Acta (Bba) Gene Regul. Mech. 2019, 1862, 370–381. [Google Scholar] [CrossRef]
- McIntyre, A.B.R.; Gokhale, N.S.; Cerchietti, L.; Jaffrey, S.R.; Horner, S.M.; Mason, C.E. Limits in the detection of m6A changes using MeRIP/m6A-seq. Sci. Rep. 2020, 10, 6590. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Lu, Z.; Wang, X.; Fu, Y.; Luo, G.-Z.; Liu, N.; Han, D.; Dominissini, D.; Dai, Q.; Pan, T.; et al. High-resolution N6-methyladenosine m6A map using photo-crosslinking-assisted m6A sequencing. Angew. Chem. Int. Ed. Engl. 2015, 54, 1587–1590. [Google Scholar] [CrossRef] [Green Version]
- Molinie, B.; Wang, J.; Lim, K.S.; Hillebrand, R.; Lu, Z.-X.; Van Wittenberghe, N.; Howard, B.D.; Daneshvar, K.; Mullen, A.C.; Dedon, P.; et al. m6A-LAIC-seq reveals the census and complexity of the m6A epitranscriptome. Nat. Methods 2016, 13, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Mikutis, S.; Gu, M.; Sendinc, E.; Hazemi, M.E.; Kiely-Collins, H.; Aspris, D.; Vassiliou, G.S.; Shi, Y.; Tzelepis, K.; Bernardes, G.J.L. meCLICK-Seq, a Substrate-Hijacking and RNA Degradation Strategy for the Study of RNA Methylation. ACS Cent Sci. 2020, 6, 2196–2208. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D. DART-seq: An antibody-free method for global m6A detection. Nat. Methods 2019, 16, 1275–1280. [Google Scholar] [CrossRef]
- Garcia-Campos, M.A.; Edelheit, S.; Toth, U.; Safra, M.; Shachar, R.; Viukov, S.; Winkler, R.; Nir, R.; Lasman, L.; Brandis, A.; et al. Deciphering the “m6A Code” via Antibody-Independent Quantitative Profiling. Cell 2019, 178, 731–747.e716. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiao, Y.; Dong, S.; Yu, Q.; Jia, G. Antibody-free enzyme-assisted chemical approach for detection of N6-methyladenosine. Nat. Chem. Biol. 2020, 16, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.; Cao, J.; Cheng, M.; Xiang, S.; Gao, M.; Li, T.; Ying, X.; Wang, F.; Yue, Y.; Lu, Z.; et al. A metabolic labeling method detects m6A transcriptome-wide at single base resolution. Nat. Chem. Biol. 2020, 16, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Anreiter, I.; Mir, Q.; Simpson, J.T.; Janga, S.C.; Soller, M. New Twists in Detecting mRNA Modification Dynamics. Trends Biotechnol. 2021, 39, 72–89. [Google Scholar] [CrossRef]
- Xu, L.; Seki, M. Recent advances in the detection of base modifications using the Nanopore sequencer. J. Hum. Genet. 2020, 65, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Lee, J.-Y.; Yang, J.-S.; Kim, J.W.; Kim, V.N.; Chang, H. The Architecture of SARS-CoV-2 Transcriptome. Cell 2020, 181, 914–921.e910. [Google Scholar] [CrossRef]
- Viehweger, A.; Krautwurst, S.; Lamkiewicz, K.; Madhugiri, R.; Ziebuhr, J.; Hölzer, M.; Marz, M. Direct RNA nanopore sequencing of full-length coronavirus genomes provides novel insights into structural variants and enables modification analysis. Genome Res. 2019, 29, 1545–1554. [Google Scholar] [CrossRef] [Green Version]
- Price, A.M.; Hayer, K.E.; McIntyre, A.B.; Gokhale, N.S.; Abebe, J.S.; Della Fera, A.N.; Weitzman, M.D. Direct RNA sequencing reveals m6A modifications on adenovirus RNA are necessary for efficient splicing. Nat. Commun. 2020, 11, 6016. [Google Scholar] [CrossRef]
- Lavi, S.; Shatkin, A.J. Methylated simian virus 40-specific RNA from nuclei and cytoplasm of infected BSC-1 cells. Proc. Natl. Acad. Sci. USA 1975, 72, 2012–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, S.I.; Green, M. Multiple methylated cap sequences in adenovirus type 2 early mRNA. J. Virol. 1976, 20, 425–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krug, R.M.; Morgan, M.A.; Shatkin, A.J. Influenza viral mRNA contains internal N6-methyladenosine and 5’-terminal 7-methylguanosine in cap structures. J. Virol. 1976, 20, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayan, P.; Ayers, D.F.; Rottman, F.M.; Maroney, P.A.; Nilsen, T.W. Unequal distribution of N6-methyladenosine in influenza virus mRNAs. Mol. Cell Biol. 1987, 7, 1572–1575. [Google Scholar] [CrossRef] [Green Version]
- Moss, B.; Gershowitz, A.; Stringer, J.R.; Holland, L.E.; Wagner, E.K. 5′-Terminal and internal methylated nucleosides in herpes simplex virus type 1 mRNA. J. Virol. 1977, 23, 234–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoltzfus, C.M.; Dimock, K. Evidence of methylation of B77 avian sarcoma virus genome RNA subunits. J. Virol. 1976, 18, 586–595. [Google Scholar] [CrossRef] [Green Version]
- Beemon, K.; Keith, J. Localization of N6-methyladenosine in the Rous sarcoma virus genome. J. Mol. Biol. 1977, 113, 165–179. [Google Scholar] [CrossRef]
- Kane, S.E.; Beemon, K. Precise localization of m6A in Rous sarcoma virus RNA reveals clustering of methylation sites: Implications for RNA processing. Mol. Cell Biol. 1985, 5, 2298–2306. [Google Scholar] [CrossRef] [Green Version]
- Hesser, C.R.; Karijolich, J.; Dominissini, D.; He, C.; Glaunsinger, B.A. N6-methyladenosine modification and the YTHDF2 reader protein play cell type specific roles in lytic viral gene expression during Kaposi’s sarcoma-associated herpesvirus infection. PLoS Pathog. 2018, 14, e1006995. [Google Scholar] [CrossRef] [Green Version]
- Lichinchi, G.; Gao, S.; Saletore, Y.; Gonzalez, G.M.; Bansal, V.; Wang, Y.; Mason, C.E.; Rana, T.M. Dynamics of the human and viral m6A RNA methylomes during HIV-1 infection of T cells. Nat. Microbiol. 2016, 1, 16011. [Google Scholar] [CrossRef]
- Kennedy, E.M.; Bogerd, H.P.; Kornepati, A.V.R.; Kang, D.; Ghoshal, D.; Marshall, J.B.; Poling, B.C.; Tsai, K.; Gokhale, N.S.; Horner, S.M.; et al. Posttranscriptional m6A Editing of HIV-1 mRNAs Enhances Viral Gene Expression. Cell Host Microbe 2016, 19, 675–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, W.; Tirumuru, N.; St Gelais, C.; Koneru, P.C.; Liu, C.; Kvaratskhelia, M.; He, C.; Wu, L. N6-Methyladenosine–binding proteins suppress HIV-1 infectivity and viral production. J. Biol. Chem. 2018, 293, 12992–13005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirumuru, N.; Zhao, B.S.; Lu, W.; Lu, Z.; He, C.; Wu, L. N6-methyladenosine of HIV-1 RNA regulates viral infection and HIV-1 Gag protein expression. Elife 2016, 5, e15528. [Google Scholar] [CrossRef]
- Jurczyszak, D.; Zhang, W.; Terry, S.N.; Kehrer, T.; Bermúdez González, M.C.; McGregor, E.; Mulder, L.C.F.; Eckwahl, M.J.; Pan, T.; Simon, V. HIV protease cleaves the antiviral m6A reader protein YTHDF3 in the viral particle. PLoS Pathog. 2020, 16, e1008305. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Jin, L.; Wang, Z.; Wang, L.; Chen, Q.; Cui, Y.; Liu, G. N6-methyladenosine regulates PEDV replication and host gene expression. Virology 2020, 548, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Courtney, D.G.; Kennedy, E.M.; Dumm, R.E.; Bogerd, H.P.; Tsai, K.; Heaton, N.S.; Cullen, B.R. Epitranscriptomic Enhancement of Influenza A Virus Gene Expression and Replication. Cell Host Microbe 2017, 22, 377–386.e375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Xu, Y.P.; Li, K.; Ye, Q.; Zhou, H.Y.; Sun, H.; Li, X.; Qin, C.F. The m6A methylome of SARS-CoV-2 in host cells. Cell Res. 2021, 31, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Arcos, S.; Rothamel, K.; Jian, J.; Rose, K.L.; McDonald, W.H.; Bian, Y.; Reasoner, S.; Barrows, N.J.; Bradrick, S.; et al. Discovery of Widespread Host Protein Interactions with the Pre-replicated Genome of CHIKV Using VIR-CLASP. Mol. Cell 2020, 78, 624–640.e627. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Hao, S.; Chen, H.; Chen, Z.; Zhang, Y.; Wang, J.; Wang, H.; Zhang, B.; Qiu, J.; Deng, F.; et al. N6-methyladenosine modification and METTL3 modulate enterovirus 71 replication. Nucleic. Acids Res. 2019, 47, 362–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M.; Zhang, Z.; Xue, M.; Zhao, B.S.; Harder, O.; Li, A.; Liang, X.; Gao, T.Z.; Xu, Y.; Zhou, J.; et al. N6-methyladenosine modification enables viral RNA to escape recognition by RNA sensor RIG-I. Nat. Microbiol. 2020, 5, 584–598. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Zhang, Q.; Zhang, R.; Lu, Y.; Wang, X.; Tian, H.; Yang, P. N6-methyladenosine RNA modification suppresses antiviral innate sensing pathways via reshaping double-stranded RNA. Nat. Commun. 2021, 12, 1582. [Google Scholar] [CrossRef]
- Tsai, K.; Courtney, D.G.; Cullen, B.R. Addition of m6A to SV40 late mRNAs enhances viral structural gene expression and replication. PLoS Pathog. 2018, 14, e1006919. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Chen, E.R.; Nilsen, T.W. Kaposi’s Sarcoma-Associated Herpesvirus Utilizes and Manipulates RNA N6-Adenosine Methylation To Promote Lytic Replication. J. Virol. 2017, 91, e00466-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, B.; Liu, H.; Zhang, S.; da Silva, S.R.; Zhang, L.; Meng, J.; Cui, X.; Yuan, H.; Sorel, O.; Zhang, S.-W.; et al. Viral and cellular N6-methyladenosine and N6,2′-O-dimethyladenosine epitranscriptomes in the KSHV life cycle. Nat. Microbiol. 2018, 3, 108–120. [Google Scholar] [CrossRef]
- Lang, F.; Singh, R.K.; Pei, Y.; Zhang, S.; Sun, K.; Robertson, E.S. EBV epitranscriptome reprogramming by METTL14 is critical for viral-associated tumorigenesis. PLoS Pathog. 2019, 15, e1007796. [Google Scholar] [CrossRef]
- Xia, T.L.; Li, X.; Wang, X.; Zhu, Y.J.; Zhang, H.; Cheng, W.; Zeng, M.S. N(6)-methyladenosine-binding protein YTHDF1 suppresses EBV replication and promotes EBV RNA decay. EMBO Rep. 2021, 22, e50128. [Google Scholar] [CrossRef]
- Imam, H.; Khan, M.; Gokhale, N.S.; McIntyre, A.B.R.; Kim, G.-W.; Jang, J.Y.; Kim, S.-J.; Mason, C.E.; Horner, S.M.; Siddiqui, A. N6-methyladenosine modification of hepatitis B virus RNA differentially regulates the viral life cycle. Proc. Natl. Acad. Sci. USA 2018, 115, 8829–8834. [Google Scholar] [CrossRef] [Green Version]
- Winkler, R.; Gillis, E.; Lasman, L.; Safra, M.; Geula, S.; Soyris, C.; Nachshon, A.; Tai-Schmiedel, J.; Friedman, N.; Le-Trilling, V.T.K.; et al. m6A modification controls the innate immune response to infection by targeting type I interferons. Nat. Immunol. 2019, 20, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; You, Y.; Lu, Z.; Yang, J.; Li, P.; Liu, L.; Xu, H.; Niu, Y.; Cao, X. N6-methyladenosine RNA modification–mediated cellular metabolism rewiring inhibits viral replication. Science 2019, 365, 1171–1176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X.; Zhang, X.; Wang, J.; Ma, Y.; Zhang, L.; Cao, X. RNA-binding protein YTHDF3 suppresses interferon-dependent antiviral responses by promoting FOXO3 translation. Proc. Natl. Acad. Sci. USA 2019, 116, 976–981. [Google Scholar] [CrossRef] [Green Version]
- Rubio, R.M.; Depledge, D.P.; Bianco, C.; Thompson, L.; Mohr, I. RNA m6A modification enzymes shape innate responses to DNA by regulating interferon β. Genes Dev. 2018, 32, 1472–1484. [Google Scholar] [CrossRef] [Green Version]
- Tirumuru, N.; Wu, L. HIV-1 envelope proteins up-regulate N (6)-methyladenosine levels of cellular RNA independently of viral replication. J. Biol. Chem. 2019, 294, 3249–3260. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.-C.; Liu, B.; Plangger, R.; Kreutz, C.; Al-Hashimi, H.M. m6A minimally impacts the structure, dynamics, and Rev ARM binding properties of HIV-1 RRE stem IIB. PLoS ONE 2019, 14, e0224850. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.-W.; Imam, H.; Khan, M.; Siddiqui, A. N6-Methyladenosine modification of hepatitis B and C viral RNAs attenuates host innate immunity via RIG-I signaling. J. Biol. Chem. 2020, 295, 13123–13133. [Google Scholar] [CrossRef]
- Antanaviciute, A.; Baquero-Perez, B.; Watson, C.M.; Harrison, S.M.; Lascelles, C.; Crinnion, L.; Markham, A.F.; Bonthron, D.T.; Whitehouse, A.; Carr, I.M. m6aViewer: Software for the detection, analysis, and visualization of N6-methyladenosine peaks from m6A-seq/ME-RIP sequencing data. RNA 2017, 23, 1493–1501. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Nott, T.J.; Jin, J.; Pawson, T. Deciphering arginine methylation: Tudor tells the tale. Nat. Rev. Mol. Cell Biol. 2011, 12, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Li, S.; Gu, D.; Xu, K.; Du, M.; Zhu, L.; Chu, H.; Zhang, Z.; Wu, Y.; Fu, Z.; et al. Genetic variants in m6A modification genes are associated with colorectal cancer risk. Carcinogenesis 2019, 41, 8–17. [Google Scholar] [CrossRef]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type I interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Wang, X.; Cai, J.; Dai, Q.; Natchiar, S.K.; Lv, R.; Chen, K.; Lu, Z.; Chen, H.; Shi, Y.G.; et al. N6-Methyladenosine methyltransferase ZCCHC4 mediates ribosomal RNA methylation. Nat. Chem. Biol. 2019, 15, 88–94. [Google Scholar] [CrossRef]
- van Tran, N.; Ernst, F.G.M.; Hawley, B.R.; Zorbas, C.; Ulryck, N.; Hackert, P.; Bohnsack, K.E.; Bohnsack, M.T.; Jaffrey, S.R.; Graille, M.; et al. The human 18S rRNA m6A methyltransferase METTL5 is stabilized by TRMT112. Nucleic Acids Res. 2019, 47, 7719–7733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Theler, D.; Kaminska, K.H.; Hiller, M.; de la Grange, P.; Pudimat, R.; Rafalska, I.; Heinrich, B.; Bujnicki, J.M.; Allain, F.H.T.; et al. The YTH domain is a novel RNA binding domain. J. Biol. Chem. 2010, 285, 14701–14710. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, T.; Newton, I.L.G.; Hardy, R.W. Wolbachia elevates host methyltransferase expression to block an RNA virus early during infection. PLoS Pathog. 2017, 13, e1006427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selberg, S.; Blokhina, D.; Aatonen, M.; Koivisto, P.; Siltanen, A.; Mervaala, E.; Kankuri, E.; Karelson, M. Discovery of Small Molecules that Activate RNA Methylation through Cooperative Binding to the METTL3-14-WTAP Complex Active Site. Cell Rep. 2019, 26, 3762–3771.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
The m6A RNA modification and its cellular interactome. The N6-methyladenosine (m6A) modification is installed co-transcriptionally to mRNA transcripts by the METTL3-METTL14-WTAP writer complex and can be removed by the FTO and ALKBH5 erasers. The family of YTH proteins comprises the major readers that specifically recognise and directly bind m6A modifications. YTHDC1 is a nuclear m6A reader and facilitates mRNA splicing and nuclear export. In the cytoplasm, YTHDF1, YTHDF2, YTHDF3 and YTHDC2 bind mRNAs to enhance their translation or induce their degradation in P-bodies. Other multiple secondary readers that also target m6A modifications have been recently elucidated. Some of these bind to m6A through structural RNA switches in the mRNA transcript induced by m6A itself.
Figure 1.
The m6A RNA modification and its cellular interactome. The N6-methyladenosine (m6A) modification is installed co-transcriptionally to mRNA transcripts by the METTL3-METTL14-WTAP writer complex and can be removed by the FTO and ALKBH5 erasers. The family of YTH proteins comprises the major readers that specifically recognise and directly bind m6A modifications. YTHDC1 is a nuclear m6A reader and facilitates mRNA splicing and nuclear export. In the cytoplasm, YTHDF1, YTHDF2, YTHDF3 and YTHDC2 bind mRNAs to enhance their translation or induce their degradation in P-bodies. Other multiple secondary readers that also target m6A modifications have been recently elucidated. Some of these bind to m6A through structural RNA switches in the mRNA transcript induced by m6A itself.

Figure 2.
The most common transcriptome-wide m6A profiling techniques used to locate m6A in viral RNAs. (A) The first step of the m6A-seq technique consists of the chemical fragmentation of the RNA into 100–200 nucleotides-long RNA molecules. An input control (which does not undergo m6A-immunoprecipitation) and an m6A-immunoprecipitated (IP) sample are required to build cDNA libraries for deep-sequencing and subsequent bioinformatic analysis. m6A-seq offers a low-resolution approach to map m6A modifications because of the relatively large size of the starting fragmented RNAs. (B) Photo-crosslinking-assisted m6A-sequencing (PA-m6A-seq) requires incubating cells with 4-thiouridine (4SU), which is incorporated into mRNAs. Full length mRNAs are then immunoprecipitated with m6A antibodies and UV-crosslinked. Next, crosslinked mRNAs are digested into around 30 nucleotides-long molecules that are isolated to prepare the cDNA libraries. Crosslinked 4SU is read as C during reverse transcription, therefore T to C transitions can be identified when compared to the reference genome. (C) In miCLiP-seq, RNA containing m6A-modifications is fragmented, immunoprecipitated (IP) using an anti-m6A antibody and UV-crosslinked to the IP antibody. As similarly to the previous techniques, during library preparation adapters are ligated to the precipitated RNA but in this case the 5′-end of the RNA is radioactively labeled. After purification of the RNA-antibody complexes using SDS-PAGE and membrane transfer, the RNA fragments are then reverse-transcribed leading to either a transition of C to T or truncations in cDNA. Following library construction and high-throughput sequencing localization of m6A modifications at single nucleotide resolution is achieved. (D) Nanopore sequencing of m6A modifications takes advantage of changes in electric current when a nucleotide containing a modification traverses a nanopore. Using specialised software, m6A modifications can then be identified. This allows for fast, accurate and sensitive detection of long sequencing reads reaching up to 2 Mb.
Figure 2.
The most common transcriptome-wide m6A profiling techniques used to locate m6A in viral RNAs. (A) The first step of the m6A-seq technique consists of the chemical fragmentation of the RNA into 100–200 nucleotides-long RNA molecules. An input control (which does not undergo m6A-immunoprecipitation) and an m6A-immunoprecipitated (IP) sample are required to build cDNA libraries for deep-sequencing and subsequent bioinformatic analysis. m6A-seq offers a low-resolution approach to map m6A modifications because of the relatively large size of the starting fragmented RNAs. (B) Photo-crosslinking-assisted m6A-sequencing (PA-m6A-seq) requires incubating cells with 4-thiouridine (4SU), which is incorporated into mRNAs. Full length mRNAs are then immunoprecipitated with m6A antibodies and UV-crosslinked. Next, crosslinked mRNAs are digested into around 30 nucleotides-long molecules that are isolated to prepare the cDNA libraries. Crosslinked 4SU is read as C during reverse transcription, therefore T to C transitions can be identified when compared to the reference genome. (C) In miCLiP-seq, RNA containing m6A-modifications is fragmented, immunoprecipitated (IP) using an anti-m6A antibody and UV-crosslinked to the IP antibody. As similarly to the previous techniques, during library preparation adapters are ligated to the precipitated RNA but in this case the 5′-end of the RNA is radioactively labeled. After purification of the RNA-antibody complexes using SDS-PAGE and membrane transfer, the RNA fragments are then reverse-transcribed leading to either a transition of C to T or truncations in cDNA. Following library construction and high-throughput sequencing localization of m6A modifications at single nucleotide resolution is achieved. (D) Nanopore sequencing of m6A modifications takes advantage of changes in electric current when a nucleotide containing a modification traverses a nanopore. Using specialised software, m6A modifications can then be identified. This allows for fast, accurate and sensitive detection of long sequencing reads reaching up to 2 Mb.
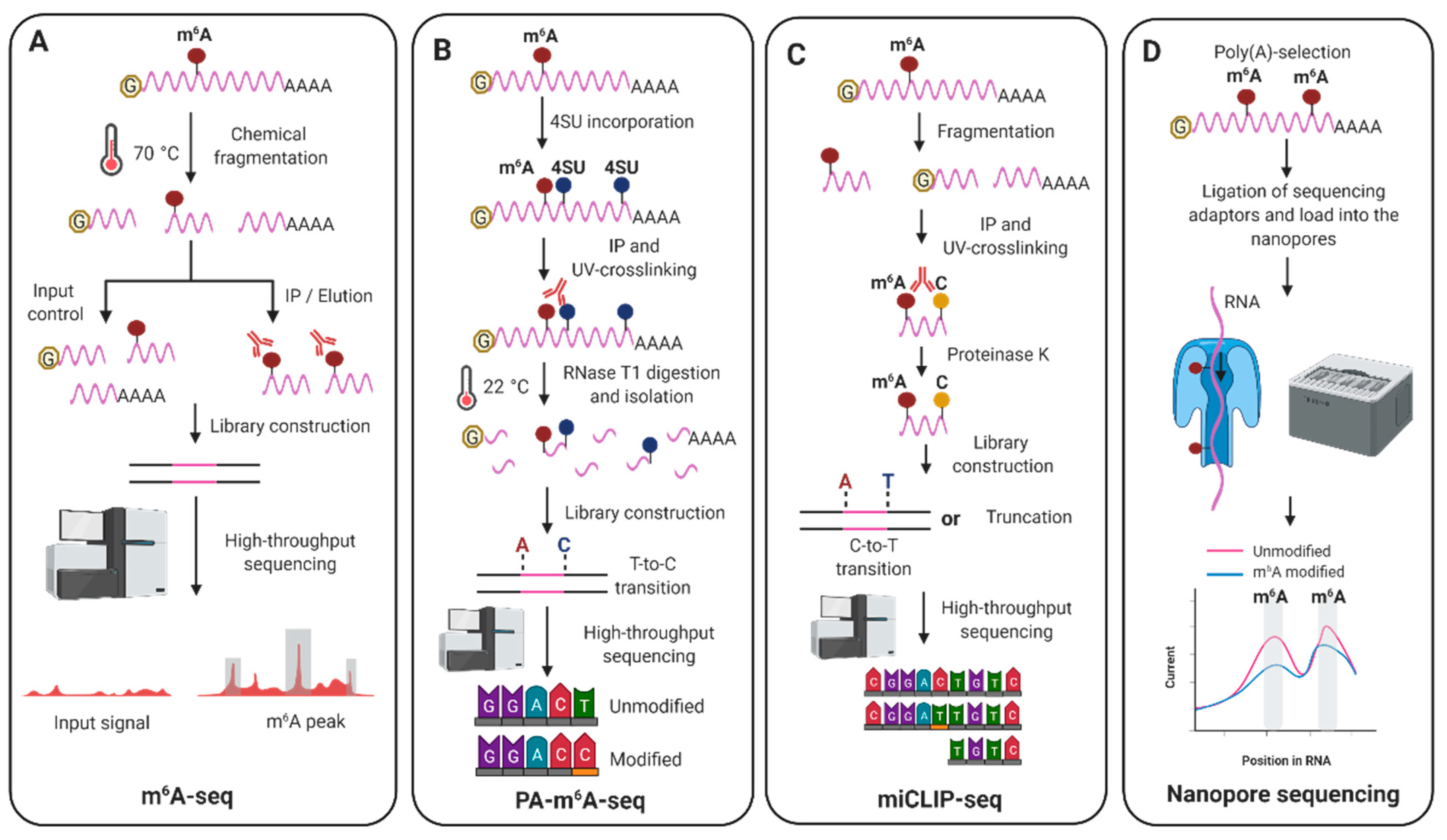
Figure 3.
How viral m6A modifications and the host m6A machinery affect viral replication. Both DNA and RNA viruses are subjected to extensive m6A modification. Importantly, m6A modifications differentially impact the steps of the viral life cycles depending on the virus. (1) m6A modifications enhance the translation of some viral mRNAs. (2) m6A modifications prevent replication through the inhibition of reverse transcription in retroviruses and enhance replication of HBV through the facilitation of reverse transcription. (3) m6A modifications reduce or increase viral mRNA stability. (4 and 5) m6A modifications affect the splicing and nuclear export of some viral RNAs that replicate in the nucleus. (6) The encapsidation of genomic RNA or assembly of viral components can be regulated by m6A modifications. (7) m6A modifications reduce or increase infectious viral titers. (8) Viral m6A decorations can prevent virus-sensing by RIG-I or MDA5 receptors and consequently inhibit type I IFN induction and anti-viral immunity.
Figure 3.
How viral m6A modifications and the host m6A machinery affect viral replication. Both DNA and RNA viruses are subjected to extensive m6A modification. Importantly, m6A modifications differentially impact the steps of the viral life cycles depending on the virus. (1) m6A modifications enhance the translation of some viral mRNAs. (2) m6A modifications prevent replication through the inhibition of reverse transcription in retroviruses and enhance replication of HBV through the facilitation of reverse transcription. (3) m6A modifications reduce or increase viral mRNA stability. (4 and 5) m6A modifications affect the splicing and nuclear export of some viral RNAs that replicate in the nucleus. (6) The encapsidation of genomic RNA or assembly of viral components can be regulated by m6A modifications. (7) m6A modifications reduce or increase infectious viral titers. (8) Viral m6A decorations can prevent virus-sensing by RIG-I or MDA5 receptors and consequently inhibit type I IFN induction and anti-viral immunity.


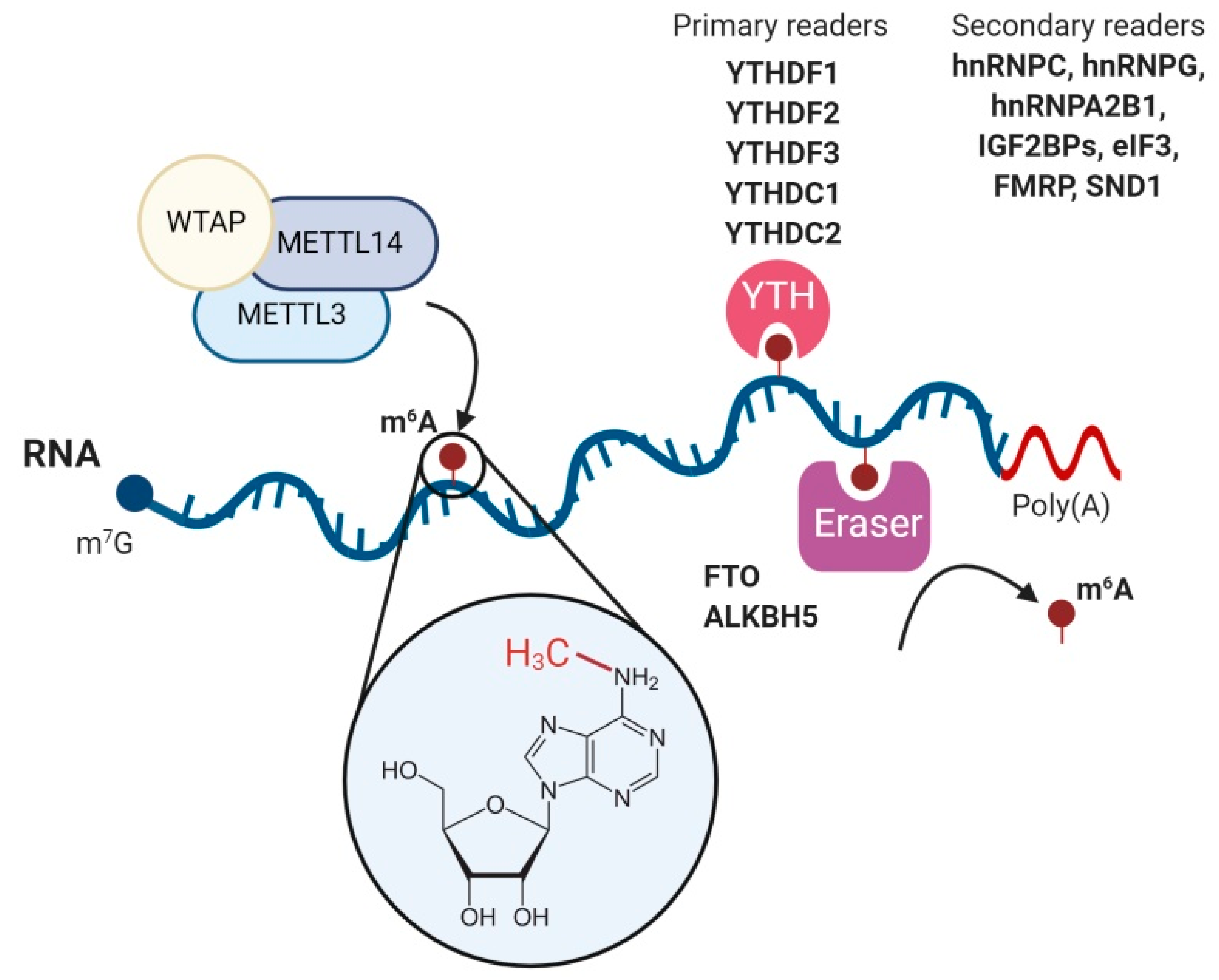{kind=link}
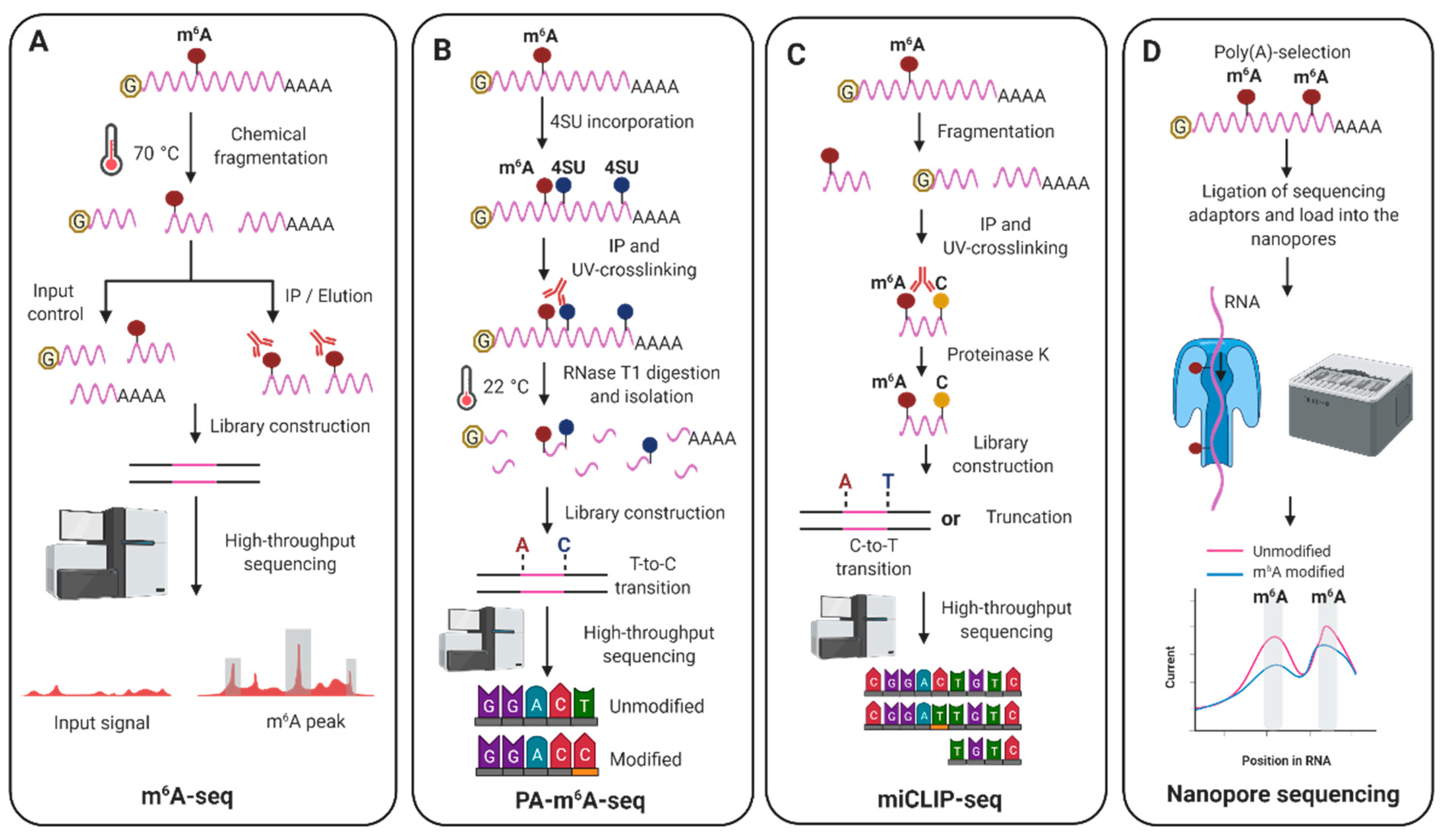{kind=link}
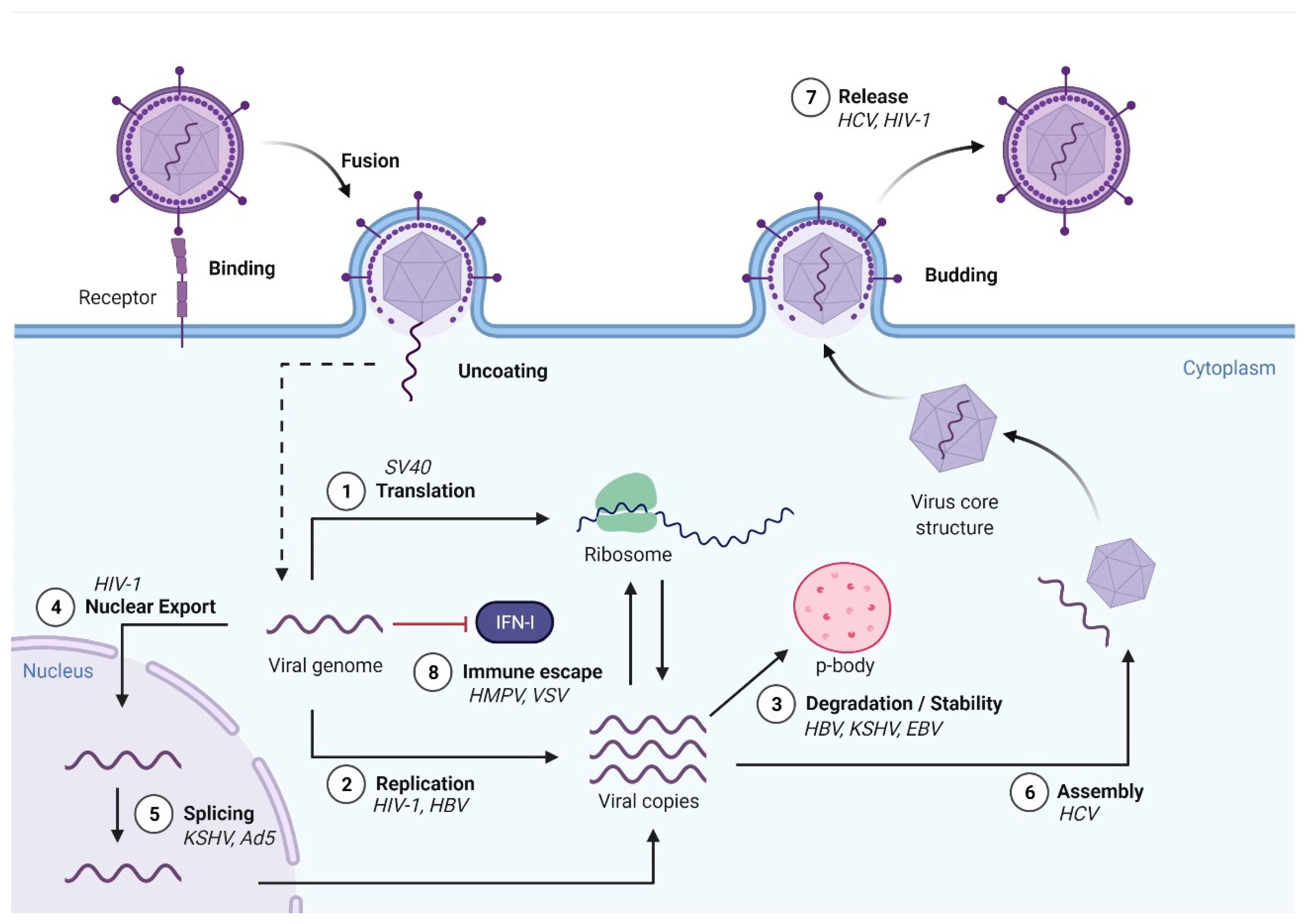{kind=link}
Table 1.
Literature on viral m6A epitranscriptomic studies.
| Virus | Detection | Phenotype | Main Function for m6A | Reference |
|---|---|---|---|---|
| RNA Viruses | ||||
| HIV-1 | m6A-seq | Proviral | vRNA nuclear export | [103] |
| PA-m6A-seq | Proviral | vRNA abundance and protein expression | [104] | |
| m6A-seq | Antiviral | YTHDF1–3 proteins inhibit HIV-1 infection by decreasing HIV-1 reverse transcription | [105,106] | |
| - | Antiviral | YTHDF3 inhibits HIV-1 infection at the step of reverse transcription | [107] | |
| HCV | m6A-seq | Antiviral | m6A modifications in E1 are bound by YTHDF proteins to negatively regulate HCV packaging | [67] |
| ZIKV | m6A-seq | Antiviral | vRNA abundance and protein expression | [68] |
| DENV WNV | m6A-seq | - | - | [67] |
| PEDV | m6A-seq | Antiviral | m6A modifications reduce viral RNA production and viral titers | [108] |
| IAV | PA-m6A-seq | Proviral | m6A modifications increase IAV RNA expression | [109] |
| RSV | m6A-seq | Proviral | m6A modifications enhance RSV replication and gene expression | [70] |
| SARS-CoV2 | m6A-seq and miCLIP | Antiviral | m6A inhibits SARS-CoV2 replication | [110] |
| CHIKV | m6A-IP | Proviral and antiviral | YTHDF1 and YTHDF3 restrict CHIKV replication, while YTHDF2 promotes it | [111] |
| EV71 | m6A-seq | Proviral | METTL3 induces enhanced sumoylation and ubiquitination of the viral RNA polymerase to facilitate viral replication | [112] |
| HMPV | m6A-seq | Proviral | m6A modifications enable vRNA to escape recognition by RIG-I | [113] |
| VSV | miCLIP | Proviral | m6A modifications reduce viral dsRNA formation leading to reduced virus-sensing by innate receptors | [114] |
| DNA VIRUSES | ||||
| SV40 | PA-m6A-seq | Proviral | m6A enhances the translation of viral late transcripts | [115] |
| KSHV | m6A-IP | Proviral | Splicing of ORF50 pre-mRNA | [116] |
| m6A-seq | Antiviral | vRNA stability of latent and lytic transcripts | [117] | |
| m6A-seq | Antiviral and proviral | vRNA abundance and protein expression | [102] | |
| m6A-seq | Proviral | vRNA abundance and protein expression | [59] | |
| EBV | m6A-seq | Both | vRNA stability of latent and lytic transcripts | [118] |
| m6A-seq and PA-m6A-seq | Antiviral | YTHDF1 promotes EBV RNA decay | [119] | |
| Ad5 | m6A-seq and nanopore | Proviral | m6A is required for splicing of adenoviral late transcripts | [93] |
| HBV | m6A-seq | Antiviral and proviral | vRNA stability and viral reverse-transcription | [120] |
Table 2.
Effects observed on viral replication after manipulation of the cellular m6A machinery. Boxes shaded in green indicate enhanced viral replication, while in red indicate reduced viral replication. Boxes shaded in orange indicate that there was no significant effect on viral replication. DNA viruses are shaded in grey boxes. HIV-1: [103,104,105,106,107]. HCV: [67]. ZIKV: [68]. CHIKV: [111]. PEDV: [108] IAV: [109,121]. RSV: [70]. SARS-CoV2: [110]. EV71: [112]. HMPV: [113]. VSV: [114,122,123]. SV40: Tsai et al., 2018. KSHV: [59,102,116,117]. EBV: [118,119]. Ad5: [93]. HBV: [120]. HCMV: [121,124].
Table 2.
Effects observed on viral replication after manipulation of the cellular m6A machinery. Boxes shaded in green indicate enhanced viral replication, while in red indicate reduced viral replication. Boxes shaded in orange indicate that there was no significant effect on viral replication. DNA viruses are shaded in grey boxes. HIV-1: [103,104,105,106,107]. HCV: [67]. ZIKV: [68]. CHIKV: [111]. PEDV: [108] IAV: [109,121]. RSV: [70]. SARS-CoV2: [110]. EV71: [112]. HMPV: [113]. VSV: [114,122,123]. SV40: Tsai et al., 2018. KSHV: [59,102,116,117]. EBV: [118,119]. Ad5: [93]. HBV: [120]. HCMV: [121,124].
| Virus | Depletion/Knockout | Overexpression | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| METTL3 | METTL14 | FTO | ALKBH5 | YTHDF1 | YTHDF2 | YTHDF3 | METTL3 | METTL14 | FTO | ALKBH5 | YTHDF1 | YTHDF2 | YTHDF3 | ||||||||||
| HIV-1 | [103,106] | [103,106] | [106] | [103,106] | [105,106] | [105,106] | [104,105] | [105,106,107] | [104] | [105,106] | [104] | [105,106] | [104] | [105,106] | |||||||||
| HCV | |||||||||||||||||||||||
| ZIKA | |||||||||||||||||||||||
| CHIKV | |||||||||||||||||||||||
| PEDV | |||||||||||||||||||||||
| IAV | [109,121] | [109] | [109] | [109] | |||||||||||||||||||
| RSV | |||||||||||||||||||||||
| SARS-2 | |||||||||||||||||||||||
| EV71 | |||||||||||||||||||||||
| HMPV | |||||||||||||||||||||||
| VSV | [122] | [114] | [122] | [122] | [122] | [123] | [123] | [123] | [114] | [122] | |||||||||||||
| SV40 | |||||||||||||||||||||||
| KSHV | [102] | [59,102,116] | [59,116] | [102,117] | [59] | [102,117] | [59,102] | [102,117] | [59] | [117] | |||||||||||||
| EBV | [119] | [118] | [118,119] | [118] | [119] | [119] | [118] | [118,119] | [119] | ||||||||||||||
| Ad5 | |||||||||||||||||||||||
| HBV | |||||||||||||||||||||||
| HCMV | [121,124] | [121,124] | [121,124] | [121,124] | [121] | [121] | [121] | ||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Baquero-Perez, B.; Geers, D.; Díez, J. From A to m6A: The Emerging Viral Epitranscriptome. Viruses 2021, 13, 1049. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061049
AMA Style
Baquero-Perez B, Geers D, Díez J. From A to m6A: The Emerging Viral Epitranscriptome. Viruses. 2021; 13(6):1049. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061049
Chicago/Turabian StyleBaquero-Perez, Belinda, Daryl Geers, and Juana Díez. 2021. "From A to m6A: The Emerging Viral Epitranscriptome" Viruses 13, no. 6: 1049. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061049
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.