
Lymphocytic Choriomeningitis Virus Alters the Expression of Male Mouse Scent Proteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
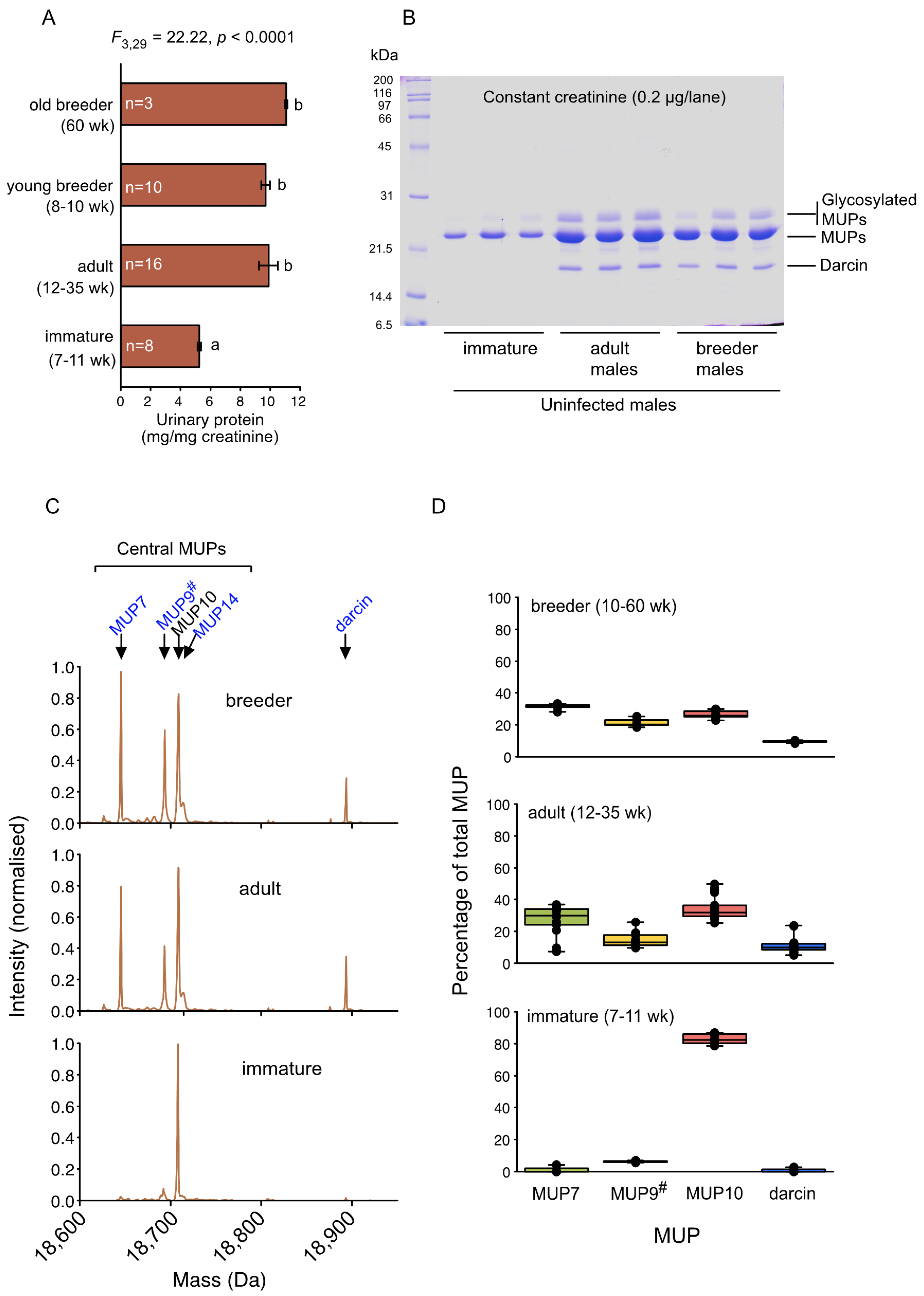
3.1. Normal Male MUP Pattern and Output
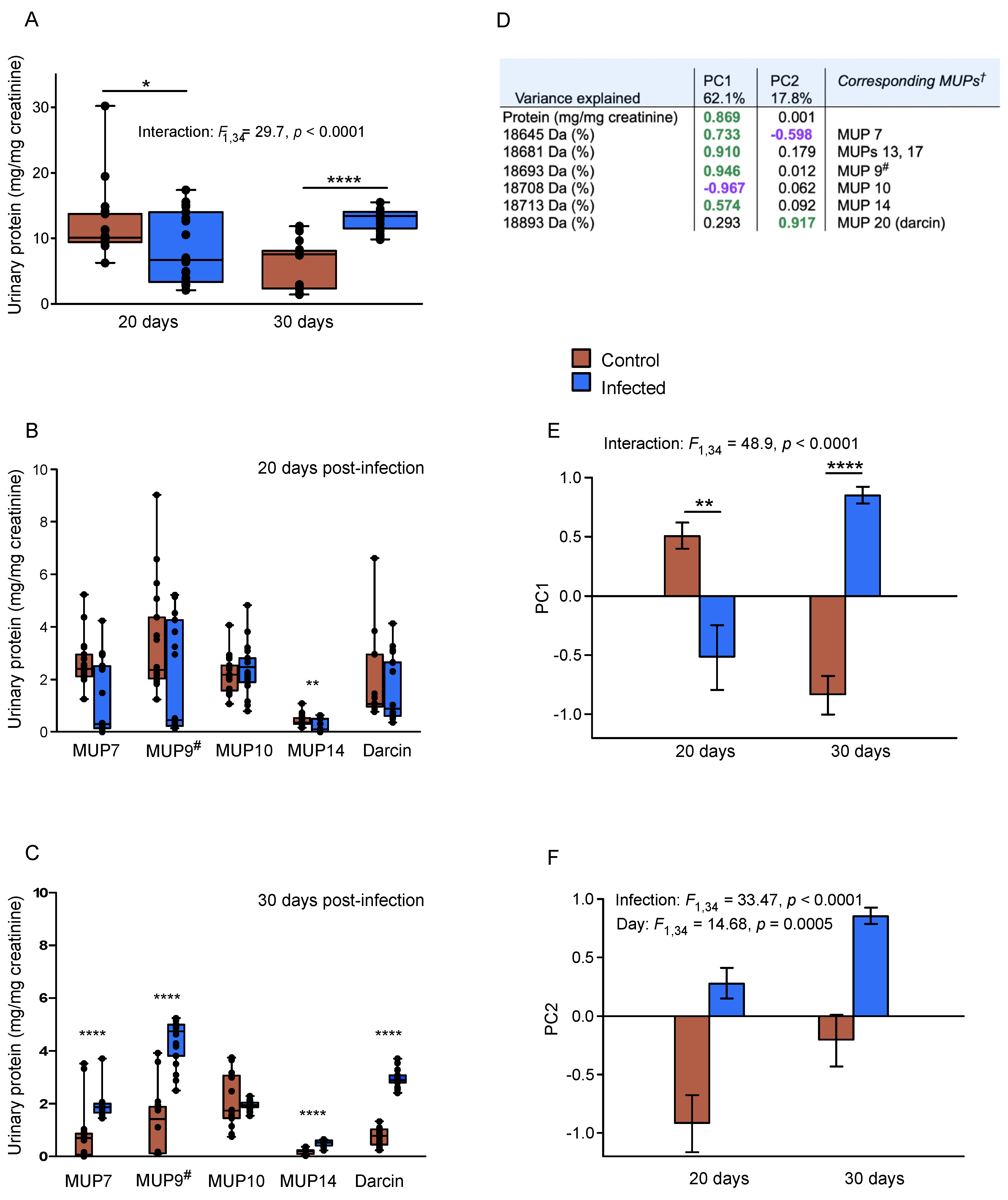
3.2. Clearance of LCMV Infection Leads to Enhanced MUP Output
3.3. Enhancement of Adult Male MUP Pattern and Darcin
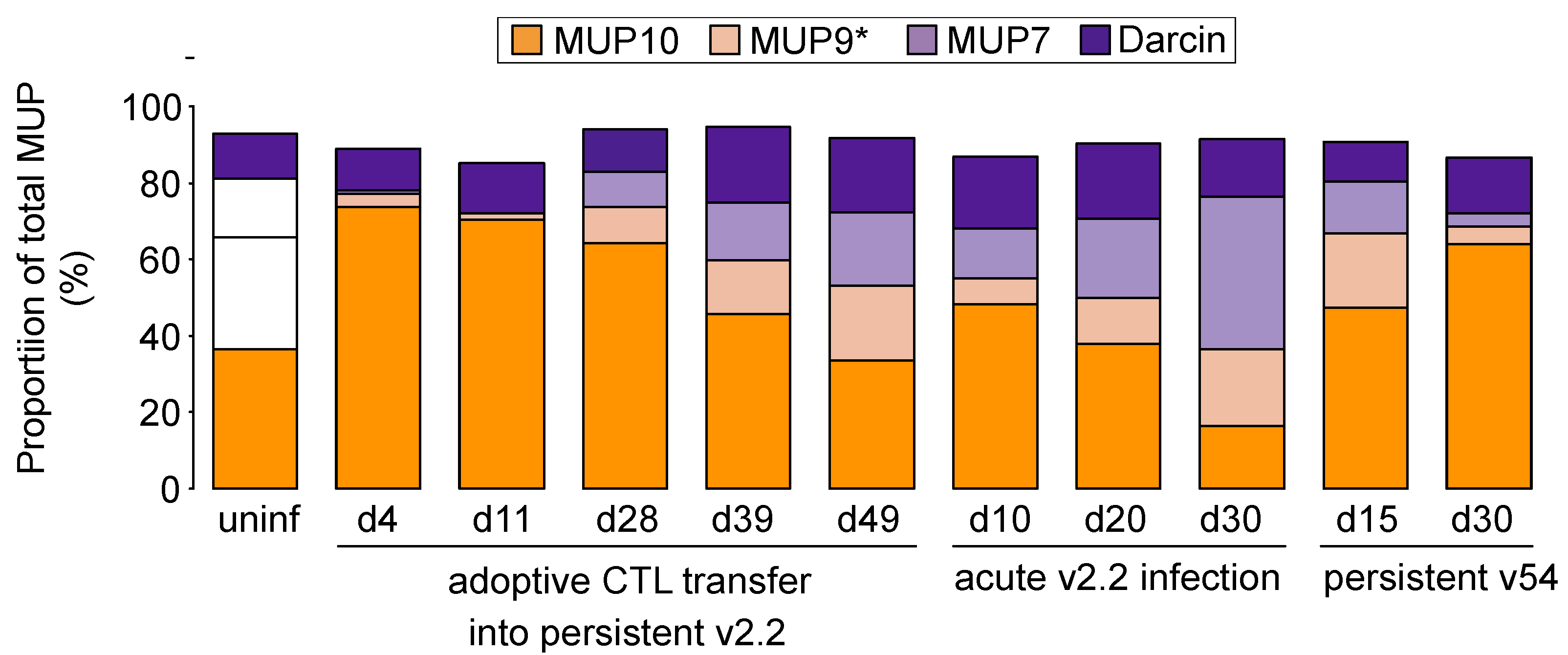
3.4. Adoptive Transfer of CTL during Persistent Infection
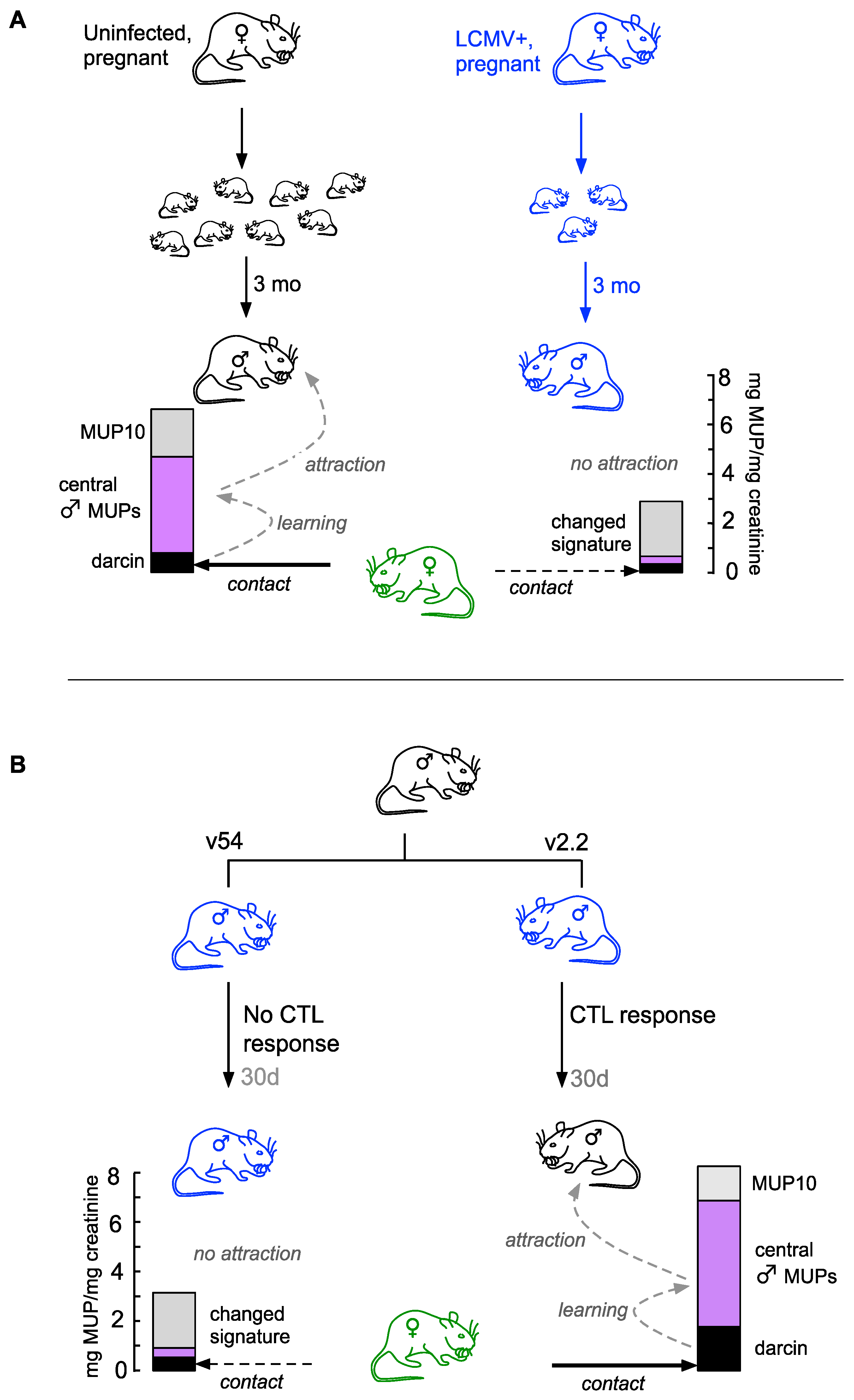
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Finlayson, J.S.; Asofsky, R.; Potter, M.; Runner, C.C. Major urinary protein complex of normal mice: Origin. Science 1965, 149, 981–982. [Google Scholar] [CrossRef] [PubMed]
- Beynon, R.J.; Hurst, J.L. Multiple roles of major urinary proteins in the house mouse, Mus domesticus. Biochem. Soc. Trans. 2003, 31, 142–146. [Google Scholar] [CrossRef] [Green Version]
- Beynon, R.J.; Hurst, J.L. Urinary proteins and the modulation of chemical scents in mice and rats. Peptides 2004, 25, 1553–1563. [Google Scholar] [CrossRef] [PubMed]
- Beynon, R.J.; Armstrong, S.D.; Gómez-Baena, G.; Lee, V.; Simpson, D.; Unsworth, J.; Hurst, J.L. The complexity of protein semiochemistry in mammals. Biochem. Soc. Trans. 2014, 42, 837–845. [Google Scholar] [CrossRef]
- Green, J.P.; Holmes, A.M.; Davidson, A.J.; Paterson, S.; Stockley, P.; Beynon, R.J.; Hurst, J.L. The Genetic Basis of Kin Recognition in a Cooperatively Breeding Mammal. Curr. Biol. 2015, 25, 2631–2641. [Google Scholar] [CrossRef] [Green Version]
- Hurst, J.L.; Beynon, R.J. Rodent urinary proteins: Genetic identity signals and pheromones. In Chemical Signals in Vertebrates 12; Springer: Berlin/Heidelberg, Germany, 2013; pp. 117–133. [Google Scholar]
- Hurst, J.L.; Payne, C.E.; Nevison, C.M.; Marie, A.D.; Humphries, R.E.; Robertson, D.H.; Cavaggioni, A.; Beynon, R.J. Individual recognition in mice mediated by major urinary proteins. Nature 2001, 414, 631–634. [Google Scholar] [CrossRef]
- Hurst, J.L.; Beynon, R.J.; Armstrong, S.D.; Davidson, A.J.; Roberts, S.A.; Gómez-Baena, G.; Smadja, C.M.; Ganem, G. Molecular heterogeneity in major urinary proteins of Mus musculus subspecies: Potential candidates involved in speciation. Sci. Rep. 2017, 7, 44992. [Google Scholar] [CrossRef] [Green Version]
- Kaur, A.W.; Ackels, T.; Kuo, T.H.; Cichy, A.; Dey, S.; Hays, C.; Kateri, M.; Logan, D.W.; Marton, T.F.; Spehr, M.; et al. Murine pheromone proteins constitute a context-dependent combinatorial code governing multiple social behaviors. Cell 2014, 157, 676–688. [Google Scholar] [CrossRef] [Green Version]
- Lopes, P.C.; König, B. Choosing a healthy mate: Sexually attractive traits as reliable indicators of current disease status in house mice. Anim. Behav. 2016, 111, 119–126. [Google Scholar] [CrossRef]
- Nelson, A.C.; Cunningham, C.B.; Ruff, J.S.; Potts, W.K. Protein pheromone expression levels predict and respond to the formation of social dominance networks. J. Evol. Biol. 2015, 28, 1213–1224. [Google Scholar] [CrossRef] [Green Version]
- Roberts, S.A.; Prescott, M.C.; Davidson, A.J.; Mclean, L.; Beynon, R.J.; Hurst, J.L. Individual odour signatures that mice learn are shaped by involatile major urinary proteins (MUPs). BMC Biol. 2018, 16, 48. [Google Scholar] [CrossRef]
- Stopka, P.; Janotova, K.; Heyrovsky, D. The advertisement role of major urinary proteins in mice. Physiol. Behav. 2007, 91, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Thoß, M.; Luzynski, K.C.; Enk, V.M.; Razzazi-Fazeli, E.; Kwak, J.; Ortner, I.; Penn, D.J. Regulation of volatile and non-volatile pheromone attractants depends upon male social status. Sci. Rep. 2019, 9, 489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demir, E.; Li, K.; Bobrowski-Khoury, N.; Sanders, J.I.; Beynon, R.J.; Hurst, J.L.; Kepecs, A.; Axel, R. The pheromone darcin drives a circuit for innate and reinforced behaviours. Nature 2020, 578, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, E.; Pickavance, L.; Thippeswamy, T.; Beynon, R.J.; Hurst, J.L. The male sex pheromone darcin stimulates hippocampal neurogenesis and cell proliferation in the subventricular zone in female mice. Front. Behav. Neurosci. 2015, 9, 106. [Google Scholar] [CrossRef] [Green Version]
- Phelan, M.M.; Mclean, L.; Simpson, D.M.; Hurst, J.L.; Beynon, R.J.; Lian, L.Y. 1H, 15N and 13C resonance assignment of darcin, a mouse major urinary protein. Biomol. NMR Assign. 2010, 4, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.A.; Simpson, D.M.; Armstrong, S.D.; Davidson, A.J.; Robertson, D.H.; Mclean, L.; Beynon, R.J.; Hurst, J.L. Darcin: A male pheromone that stimulates female memory and sexual attraction to an individual male’s odour. BMC Biol. 2010, 8, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austen, J. Pride and Prejudice; T. Egerton: Whitehall, UK, 1813. [Google Scholar]
- Roberts, S.A.; Davidson, A.J.; Mclean, L.; Beynon, R.J.; Hurst, J.L. Pheromonal induction of spatial learning in mice. Science 2012, 338, 1462–1465. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, M.J.; Lee, V.; Corbett-Detig, R.; Bi, K.; Beynon, R.J.; Hurst, J.L.; Nachman, M.W. Selection on Coding and Regulatory Variation Maintains Individuality in Major Urinary Protein Scent Marks in Wild Mice. PLoS Genet. 2016, 12, e1005891. [Google Scholar] [CrossRef] [Green Version]
- Beynon, R.J.; Armstron, S.D.; Claydon, A.J.; Davidson, A.; Eyers, C.E.; Langridge, J.I.; Gómez-Baena, G.; Harman, V.M.; Hurst, J.L.; Victoria, L.; et al. Mass spectrometry for structural analysis and quantification of the Major Urinary Proteins of the house mouse. Int. J. Mass Spectrom. 2015, 391, 146–156. [Google Scholar] [CrossRef]
- Mudge, J.M.; Armstrong, S.D.; Mclaren, K.; Beynon, R.J.; Hurst, J.L.; Nicholson, C.; Robertson, D.H.; Wilming, L.G.; Harrow, J.L. Dynamic instability of the major urinary protein gene family revealed by genomic and phenotypic comparisons between C57 and 129 strain mice. Genome Biol. 2008, 9, R91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janotova, K.; Stopka, P. The level of major urinary proteins is socially regulated in wild Mus musculus musculus. J. Chem. Ecol. 2011, 37, 647–656. [Google Scholar] [CrossRef]
- Stockley, P.; Bottell, L.; Hurst, J.L. Wake up and smell the conflict: Odour signals in female competition. Philos. Trans. R. Soc. Lond B Biol. Sci. 2013, 368, 20130082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, A.C.; Cauceglia, J.W.; Merkley, S.D.; Youngson, N.A.; Oler, A.J.; Nelson, R.J.; Cairns, B.R.; Whitelaw, E.; Potts, W.K. Reintroducing domesticated wild mice to sociality induces adaptive transgenerational effects on MUP expression. Proc. Natl. Acad. Sci. USA 2013, 110, 19848–19853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, M. Sexual Selection; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Wyatt, T.D. Pheromones and Animal Behavior; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Teng, M.N.; Borrow, P.; Oldstone, M.B.; De La Torre, J.C. A single amino acid change in the glycoprotein of lymphocytic choriomeningitis virus is associated with the ability to cause growth hormone deficiency syndrome. J. Virol. 1996, 70, 8438–8443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, M.N.; Oldstone, M.B.; De La Torre, J.C. Suppression of lymphocytic choriomeningitis virus—Induced growth hormone deficiency syndrome by disease-negative virus variants. Virology 1996, 223, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Ware, B.C.; Sullivan, B.M.; Lavergne, S.; Marro, B.S.; Egashira, T.; Campbell, K.P.; Elder, J.; Oldstone, M.B.A. A unique variant of lymphocytic choriomeningitis virus that induces pheromone binding protein MUP: Critical role for CTL. Proc. Natl. Acad. Sci. USA 2019, 116, 18001–18008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipkin, W.I.; Villarreal, L.P.; Oldstone, M.B. Whole animal section in situ hybridization and protein blotting: New tools in molecular analysis of animal models for human disease. Curr. Top. Microbiol. Immunol. 1989, 143, 33–54. [Google Scholar] [CrossRef] [PubMed]
- Oldstone, M.B. Biology and pathogenesis of lymphocytic choriomeningitis virus infection. Curr. Top. Microbiol. Immunol. 2002, 263, 83–117. [Google Scholar] [CrossRef]
- Roberts, S.C.; Little, A.C.; Burriss, R.P.; Cobey, K.D.; Klapilová, K.; Havlíček, J.; Jones, B.C.; Debruine, L.; Petrie, M. Partner choice, relationship satisfaction, and oral contraception: The congruency hypothesis. Psychol. Sci. 2014, 25, 1497–1503. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, R.; Oldstone, M.B. Organ-specific selection of viral variants during chronic infection. J. Exp. Med. 1988, 167, 1719–1724. [Google Scholar] [CrossRef]
- Berger, D.P.; Homann, D.; Oldstone, M.B. Defining parameters for successful immunocytotherapy of persistent viral infection. Virology 2000, 266, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Kunz, S.; Sevilla, N.; Mcgavern, D.B.; Campbell, K.P.; Oldstone, M.B. Molecular analysis of the interaction of LCMV with its cellular receptor [alpha]-dystroglycan. J. Cell Biol. 2001, 155, 301–310. [Google Scholar] [CrossRef]
- Cheetham, S.A.; Thom, M.D.; Jury, F.; Ollier, W.E.; Beynon, R.J.; Hurst, J.L. The genetic basis of individual-recognition signals in the mouse. Curr. Biol. 2007, 17, 1771–1777. [Google Scholar] [CrossRef] [Green Version]
- Evershed, R.P.; Robertson, D.H.; Beynon, R.J.; Green, B.N. Application of electrospray ionization mass spectrometry with maximum-entropy analysis to allelic ‘fingerprinting’ of major urinary proteins. Rapid Commun. Mass Spectrom. 1993, 7, 882–886. [Google Scholar] [CrossRef]
- Robertson, D.H.; Hurst, J.L.; Searle, J.B.; Gündüz, I.; Beynon, R.J. Characterization and comparison of major urinary proteins from the house mouse, Mus musculus domesticus, and the aboriginal mouse, Mus macedonicus. J. Chem. Ecol. 2007, 33, 613–630. [Google Scholar] [CrossRef] [PubMed]
- Cheetham, S.A.; Smith, A.L.; Armstrong, S.D.; Beynon, R.J.; Hurst, J.L. Limited variation in the major urinary proteins of laboratory mice. Physiol. Behav. 2009, 96, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Clissold, P.M.; Bishop, J.O. Variation in mouse major urinary protein (MUP) genes and the MUP gene products within and between inbred lines. Gene 1982, 18, 211–220. [Google Scholar] [CrossRef]
- Armstrong, S.D.; Robertson, D.H.; Cheetham, S.A.; Hurst, J.L.; Beynon, R.J. Structural and functional differences in isoforms of mouse major urinary proteins: A male-specific protein that preferentially binds a male pheromone. Biochem. J. 2005, 391, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.; Khan, A.; Curley, J.P. Major urinary protein levels are associated with social status and context in mouse social hierarchies. Proc. Biol. Sci. 2017, 284. [Google Scholar] [CrossRef]
- Berger, F.G.; Szoka, P. Biosynthesis of the major urinary proteins in mouse liver: A biochemical genetic study. Biochem. Genet. 1981, 19, 1261–1273. [Google Scholar] [CrossRef]
- Derous, D.; Mitchell, S.E.; Wang, L.; Green, C.L.; Wang, Y.; Chen, L.; Han, J.J.; Promislow, D.E.L.; Lusseau, D.; Douglas, A.; et al. The effects of graded levels of calorie restriction: XI. Evaluation of the main hypotheses underpinning the life extension effects of CR using the hepatic transcriptome. Aging 2017, 9, 1770–1824. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, S.J.; Madrigal-Matute, J.; Scheibye-Knudsen, M.; Fang, E.; Aon, M.; González-Reyes, J.A.; Cortassa, S.; Kaushik, S.; Gonzalez-Freire, M.; Patel, B.; et al. Effects of Sex, Strain, and Energy Intake on Hallmarks of Aging in Mice. Cell Metab. 2016, 23, 1093–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallauf, K.; Günther, I.; Chin, D.; Rimbach, G. In Contrast to Dietary Restriction, Application of Resveratrol in Mice Does not Alter Mouse Major Urinary Protein Expression. Nutrients 2020, 12, 815. [Google Scholar] [CrossRef] [Green Version]
- Hui, X.; Zhu, W.; Wang, Y.; Lam, K.S.; Zhang, J.; Wu, D.; Kraegen, E.W.; Li, Y.; Xu, A. Major urinary protein-1 increases energy expenditure and improves glucose intolerance through enhancing mitochondrial function in skeletal muscle of diabetic mice. J. Biol. Chem. 2009, 284, 14050–14057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isseroff, H.; Sylvester, P.W.; Held, W.A. Effects of Schistosoma mansoni on androgen regulated gene expression in the mouse. Mol. Biochem. Parasitol. 1986, 18, 401–412. [Google Scholar] [CrossRef]
- Manivannan, B.; Rawson, P.; Jordan, T.W.; Secor, W.E.; La Flamme, A.C. Differential patterns of liver proteins in experimental murine hepatosplenic schistosomiasis. Infect. Immun. 2010, 78, 618–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garratt, M.; Stockley, P.; Armstrong, S.D.; Beynon, R.J.; Hurst, J.L. The scent of senescence: Sexual signalling and female preference in house mice. J. Evol. Biol. 2011, 24, 2398–2409. [Google Scholar] [CrossRef] [PubMed]
- Lanuza, E.; Martín-Sánchez, A.; Marco-Manclús, P.; Cádiz-Moretti, B.; Fortes-Marco, L.; Hernández-Martínez, A.; Mclean, L.; Beynon, R.J.; Hurst, J.L.; Martínez-García, F. Sex pheromones are not always attractive: Changes induced by learning and illness in mice. Anim. Behav. 2014, 97, 265–272. [Google Scholar] [CrossRef]
- Berry, A.; Bellisario, V.; Capoccia, S.; Tirassa, P.; Calza, A.; Alleva, E.; Cirulli, F. Social deprivation stress is a triggering factor for the emergence of anxiety- and depression-like behaviours and leads to reduced brain BDNF levels in C57BL/6J mice. Psychoneuroendocrinology 2012, 37, 762–772. [Google Scholar] [CrossRef]
- Garratt, M.; Mcardle, F.; Stockley, P.; Vasilaki, A.; Beynon, R.J.; Jackson, M.J.; Hurst, J.L. Tissue-dependent changes in oxidative damage with male reproductive effort in house mice. Funct. Ecol. 2012, 26, 423–433. [Google Scholar] [CrossRef]
- Rivière, Y.; Gresser, I.; Guillon, J.C.; Bandu, M.T.; Ronco, P.; Morel-Maroger, L.; Verroust, P. Severity of lymphocytic choriomeningitis virus disease in different strains of suckling mice correlates with increasing amounts of endogenous interferon. J. Exp. Med. 1980, 152, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Childs, J.E.; Glass, G.E.; Korch, G.W.; Ksiazek, T.G.; Leduc, J.W. Lymphocytic choriomeningitis virus infection and house mouse (Mus musculus) distribution in urban Baltimore. Am. J. Trop. Med. Hyg. 1992, 47, 27–34. [Google Scholar] [CrossRef]
- Blasdell, K.R.; Becker, S.D.; Hurst, J.; Begon, M.; Bennett, M. Host range and genetic diversity of arenaviruses in rodents, United Kingdom. Emerg. Infect. Dis. 2008, 14, 1455–1458. [Google Scholar] [CrossRef] [PubMed]
- Morita, C.; Matsuura, Y.; Kawashima, E.; Takahashi, S.; Kawaguchi, J.; Iida, S.; Yamanaka, T.; Jitsukawa, W. Seroepidemiological survey of lymphocytic choriomeningitis virus in wild house mouse (Mus musculus) in Yokohama Port, Japan. J. Vet. Med. Sci. 1991, 53, 219–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, J.; Grigsby, C.C.; Rizki, M.M.; Preti, G.; Köksal, M.; Josue, J.; Yamazaki, K.; Beauchamp, G.K. Differential binding between volatile ligands and major urinary proteins due to genetic variation in mice. Physiol. Behav. 2012, 107, 112–120. [Google Scholar] [CrossRef]
- Rich, T.; Hurst, J.L. The competing countermarks hypothesis: Reliable assessment of competitive ability by potential mates. Anim. Behav. 1999, 58, 1027–1037. [Google Scholar] [CrossRef] [Green Version]
- Martínez-García, F.; Martínez-Ricós, J.; Agustín-Pavón, C.; Martínez-Hernández, J.; Novejarque, A.; Lanuza, E. Refining the dual olfactory hypothesis: Pheromone reward and odour experience. Behav. Brain Res. 2009, 200, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Ramm, S.A.; Cheetham, S.A.; Hurst, J.L. Encoding choosiness: Female attraction requires prior physical contact with individual male scents in mice. Proc. Biol. Sci. 2008, 275, 1727–1735. [Google Scholar] [CrossRef] [Green Version]
- Kreshover, S.J.; Hancock, J.A. The effect of lymphocytic choriomeningitis on pregnancy and dental tissues in mice. J. Dent. Res. 1956, 35, 467–478. [Google Scholar] [CrossRef]
- Oldstone, M.B.A.; Ware, B.C.; Horton, L.E.; Welch, M.J.; Aiolfi, R.; Zarpellon, A.; Ruggeri, Z.M.; Sullivan, B.M. Lymphocytic choriomeningitis virus Clone 13 infection causes either persistence or acute death dependent on IFN-1, cytotoxic T lymphocytes (CTLs), and host genetics. Proc. Natl. Acad. Sci. USA 2018, 115, E7814–E7823. [Google Scholar] [CrossRef] [PubMed]
- Asano, M.S.; Ahmed, R. CD8 T cell memory in B cell-deficient mice. J. Exp. Med. 1996, 183, 2165–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, H.M. An exteroceptive block to pregnancy in the mouse. Nature 1959, 184, 105. [Google Scholar] [CrossRef] [PubMed]
- Brennan, P.; Kaba, H.; Keverne, E.B. Olfactory recognition: A simple memory system. Science 1990, 250, 1223–1226. [Google Scholar] [CrossRef] [PubMed]
- Kaba, H.; Rosser, A.; Keverne, B. Neural basis of olfactory memory in the context of pregnancy block. Neuroscience 1989, 32, 657–662. [Google Scholar] [CrossRef]
- Becker, S.D.; Hurst, J.L. Female behaviour plays a critical role in controlling murine pregnancy block. Proc. Biol. Sci. 2009, 276, 1723–1729. [Google Scholar] [CrossRef] [Green Version]
- Brennan, P.A. Outstanding issues surrounding vomeronasal mechanisms of pregnancy block and individual recognition in mice. Behav. Brain Res. 2009, 200, 287–294. [Google Scholar] [CrossRef]
- Brennan, P.A. The nose knows who’s who: Chemosensory individuality and mate recognition in mice. Horm. Behav. 2004, 46, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Stokes, R.H.; Sandel, A.A. Data quality and the comparative method: The case of pregnancy failure in rodents. J. Mammal. 2019, 100, 1436–1446. [Google Scholar] [CrossRef]
- Leinders-Zufall, T.; Brennan, P.; Widmayer, P.; Chandramani, S.P.; Maul-Pavicic, A.; Jäger, M.; Li, X.H.; Breer, H.; Zufall, F.; Boehm, T. MHC class I peptides as chemosensory signals in the vomeronasal organ. Science 2004, 306, 1033–1037. [Google Scholar] [CrossRef] [Green Version]
- Thomas, K.J.; Preeji, K.P.; Ranjith, S. Imprinting of a Nonpheromonal Cue and Its Protective Effect on Alien Male-Induced Implantation Failure in Mice. Chem. Sens. 2018, 43, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, K.; Beauchamp, G.K.; Wysocki, C.J.; Bard, J.; Thomas, L.; Boyse, E.A. Recognition of H-2 types in relation to the blocking of pregnancy in mice. Science 1983, 221, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Peele, P.; Salazar, I.; Mimmack, M.; Keverne, E.B.; Brennan, P.A. Low molecular weight constituents of male mouse urine mediate the pregnancy block effect and convey information about the identity of the mating male. Eur. J. Neurosci. 2003, 18, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Hattori, T.; Osakada, T.; Masaoka, T.; Ooyama, R.; Horio, N.; Mogi, K.; Nagasawa, M.; Haga-Yamanaka, S.; Touhara, K.; Kikusui, T. Exocrine Gland-Secreting Peptide 1 Is a Key Chemosensory Signal Responsible for the Bruce Effect in Mice. Curr. Biol. 2017, 27, 3197–3201.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaba, H.; Fujita, H.; Agatsuma, T.; Matsunami, H. Maternally inherited peptides as strain-specific chemosignals. Proc. Natl. Acad. Sci. USA 2020, 117, 30738–30743. [Google Scholar] [CrossRef] [PubMed]
- Coopersmith, C.B.; Lenington, S. Pregnancy block in house mice (Mus domesticus) as a function of t-complex genotype: Examination of the mate choice and male infanticide hypotheses. J. Comp. Psychol. 1998, 112, 82–91. [Google Scholar] [CrossRef]
- Labov, J.B. Pregnancy blocking in rodents: Adaptive advantages for females. Am. Nat. 1981, 118, 361–371. [Google Scholar] [CrossRef]
- Rülicke, T.; Guncz, N.; Wedekind, C. Early maternal investment in mice: No evidence for compatible-genes sexual selection despite hybrid vigor. J. Evolut. Biol. 2006, 19, 922–928. [Google Scholar] [CrossRef]
- Sturm, T.; Leinders-Zufall, T.; Maček, B.; Walzer, M.; Jung, S.; Pömmerl, B.; Stevanović, S.; Zufall, F.; Overath, P.; Rammensee, H.G. Mouse urinary peptides provide a molecular basis for genotype discrimination by nasal sensory neurons. Nat. Commun. 2013, 4, 1616. [Google Scholar] [CrossRef]
- Fazakerley, J.K.; Southern, P.; Bloom, F.; Buchmeier, M.J. High resolution in situ hybridization to determine the cellular distribution of lymphocytic choriomeningitis virus RNA in the tissues of persistently infected mice: Relevance to arenavirus disease and mechanisms of viral persistence. J. Gen. Virol. 1991, 72, 1611–1625. [Google Scholar] [CrossRef]
- Haga, S.; Hattori, T.; Sato, T.; Sato, K.; Matsuda, S.; Kobayakawa, R.; Sakano, H.; Yoshihara, Y.; Kikusui, T.; Touhara, K. The male mouse pheromone ESP1 enhances female sexual receptive behaviour through a specific vomeronasal receptor. Nature 2010, 466, 118–122. [Google Scholar] [CrossRef]
- Ng, C.T.; Sullivan, B.M.; Oldstone, M.B. The role of dendritic cells in viral persistence. Curr. Opin. Virol. 2011, 1, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Zinkernagel, R.M.; Doherty, P.C. Restriction of in vitro T cell-mediated cytotoxicity in lymphocytic choriomeningitis within a syngeneic or semiallogeneic system. Nature 1974, 248, 701–702. [Google Scholar] [CrossRef]
- Zinkernagel, R.M.; Oldstone, M.B. Cells that express viral antigens but lack H-2 determinants are not lysed by immune thymus-derived lymphocytes but are lysed by other antiviral immune attack mechanisms. Proc. Natl. Acad. Sci. USA 1976, 73, 3666–3670. [Google Scholar] [CrossRef] [Green Version]
- Singer, A.G.; Tsuchiya, H.; Wellington, J.L.; Beauchamp, G.K.; Yamazaki, K. Chemistry of odortypes in mice: Fractionation and bioassay. J. Chem. Ecol. 1993, 19, 569–579. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Yamazaki, K.; Beauchamp, G.K.; Bard, J.; Thomas, L.; Boyse, E.A. Distinctive urinary odors governed by the major histocompatibility locus of the mouse. Proc. Natl. Acad. Sci. USA 1981, 78, 5817–5820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, K.; Beauchamp, G.K.; Kupniewski, D.; Bard, J.; Thomas, L.; Boyse, E.A. Familial imprinting determines H-2 selective mating preferences. Science 1988, 240, 1331–1332. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, K.; Singer, A.; Beauchamp, G.K. Origin, functions and chemistry of H-2 regulated odorants. Genetica 1999, 104, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, K.; Beauchamp, G.K.; Curran, M.; Bard, J.; Boyse, E.A. Parent-progeny recognition as a function of MHC odortype identity. Proc. Natl. Acad. Sci. USA 2000, 97, 10500–10502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, L.S.; Penn, D.J.; Potts, W.K. Discrimination of MHC-derived odors by untrained mice is consistent with divergence in peptide-binding region residues. Proc. Natl. Acad. Sci. USA 2002, 99, 2187–2192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, J.; Willse, A.; Preti, G.; Yamazaki, K.; Beauchamp, G.K. In search of the chemical basis for MHC odourtypes. Proc. Biol. Sci. 2010, 277, 2417–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overath, P.; Sturm, T.; Rammensee, H.G. Of volatiles and peptides: In search for MHC-dependent olfactory signals in social communication. Cell Mol. Life Sci. 2014, 71, 2429–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willse, A.; Kwak, J.; Yamazaki, K.; Preti, G.; Wahl, J.H.; Beauchamp, G.K. Individual odortypes: Interaction of MHC and background genes. Immunogenetics 2006, 58, 967–982. [Google Scholar] [CrossRef] [PubMed]
- Sherborne, A.L.; Thom, M.D.; Paterson, S.; Jury, F.; Ollier, W.E.; Stockley, P.; Beynon, R.J.; Hurst, J.L. The genetic basis of inbreeding avoidance in house mice. Curr. Biol. 2007, 17, 2061–2066. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oldstone, M.B.A.; Ware, B.C.; Davidson, A.; Prescott, M.C.; Beynon, R.J.; Hurst, J.L. Lymphocytic Choriomeningitis Virus Alters the Expression of Male Mouse Scent Proteins. Viruses 2021, 13, 1180. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061180
Oldstone MBA, Ware BC, Davidson A, Prescott MC, Beynon RJ, Hurst JL. Lymphocytic Choriomeningitis Virus Alters the Expression of Male Mouse Scent Proteins. Viruses. 2021; 13(6):1180. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061180
Chicago/Turabian StyleOldstone, Michael B. A., Brian C. Ware, Amanda Davidson, Mark C. Prescott, Robert J. Beynon, and Jane L. Hurst. 2021. "Lymphocytic Choriomeningitis Virus Alters the Expression of Male Mouse Scent Proteins" Viruses 13, no. 6: 1180. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061180