
Neutralizing Antibody Induction Associated with a Germline Immunoglobulin Gene Polymorphism in Neutralization-Resistant SIVsmE543-3 Infection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Determination of Germline VH3.33 and TRIM5 Alleles
2.3. Analysis of Plasma Neutralizing Activity
2.4. NGS Analysis of BCR IgG VH cDNAs
3. Results
3.1. Germline Ig VH3.33 Polymorphisms in Rhesus and Cynomolgus Macaques
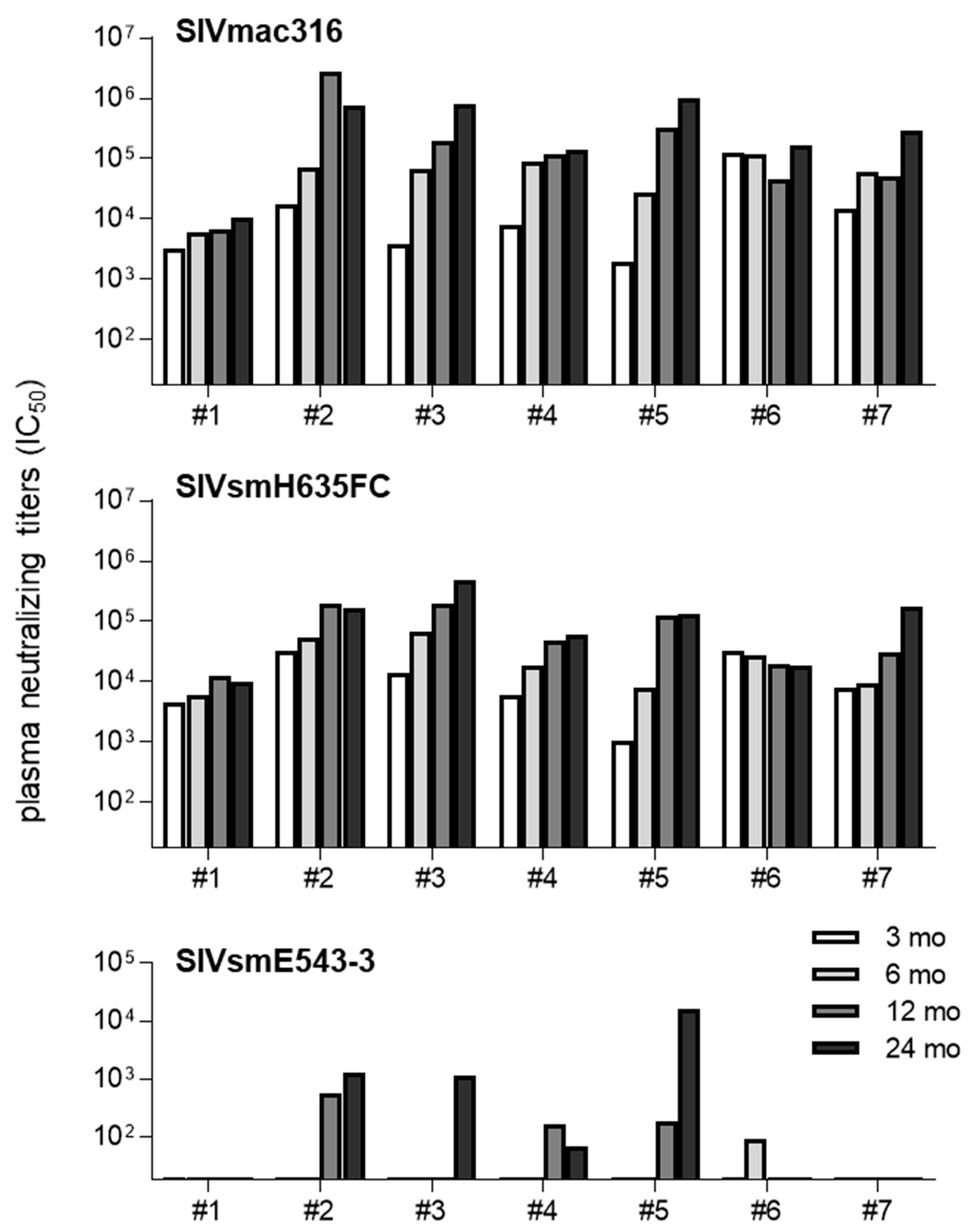
3.2. Neutralizing Antibody Responses in SIVsmE543-3-Infected Rhesus Macaques
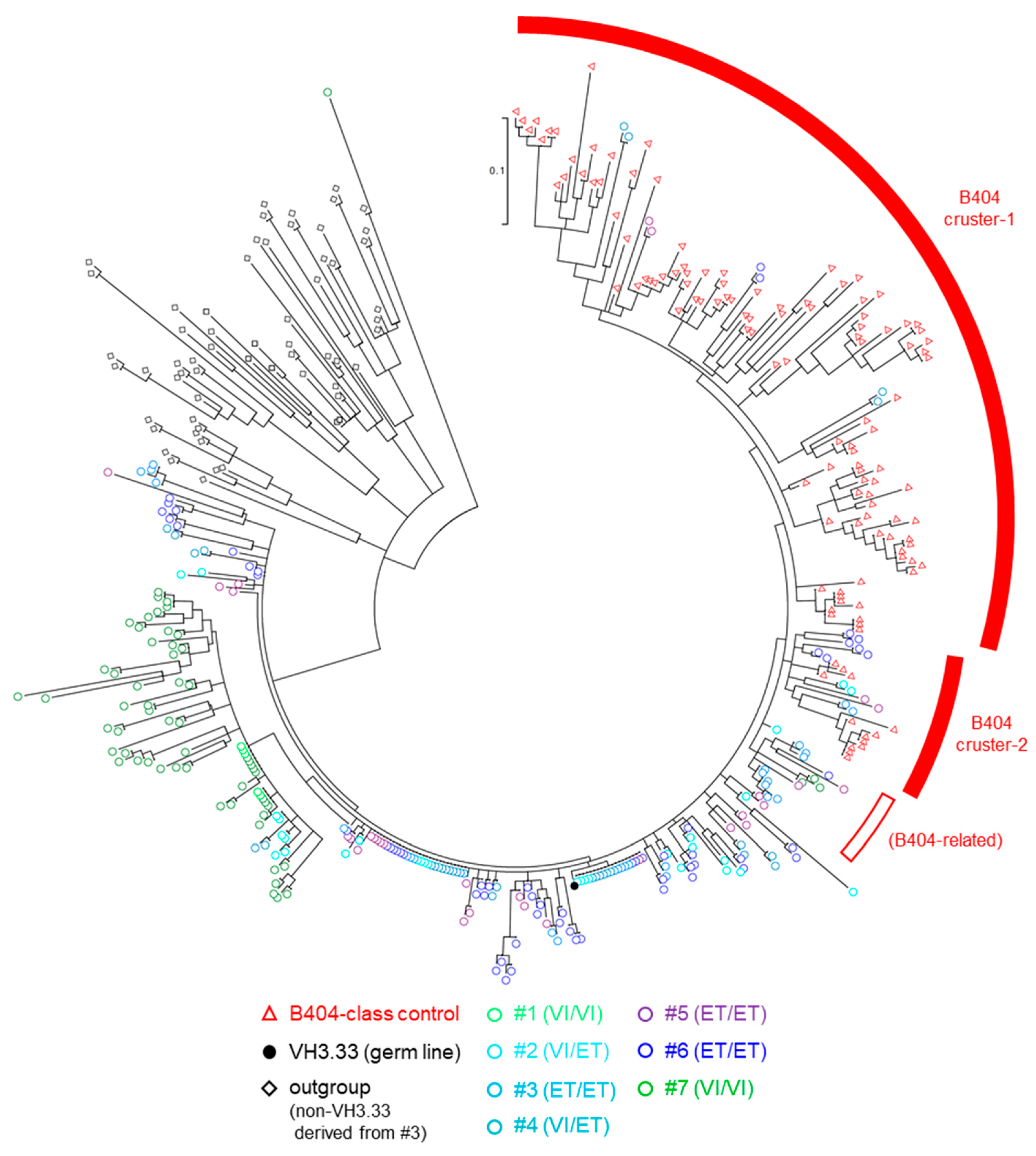
3.3. Analysis of BCR VH3.33 Sequences Derived from PBMCs and LNs by NGS
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiedermann, U.; Garner-Spitzer, E.; Wagner, A. Primary vaccine failure to routine vaccines: Why and what to do? Hum. Vaccin. Immunother. 2016, 12, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Cotugno, N.; Ruggiero, A.; Santilli, V.; Manno, E.C.; Rocca, S.; Zicari, S.; Amodio, D.; Colucci, M.; Rossi, P.; Levy, O.; et al. OMIC Technologies and Vaccine Development: From the Identification of Vulnerable Individuals to the Formulation of Invulnerable Vaccines. J. Immunol. Res. 2019, 8732191. [Google Scholar] [CrossRef] [Green Version]
- Kubba, A.K.; Taylor, P.; Graneek, B.; Strobel, S. Non-responders to hepatitis B vaccination: A review. Commun. Dis. Public Health 2003, 6, 106–112. [Google Scholar] [PubMed]
- Rendi-Wagner, P.; Korinek, M.; Winkler, B.; Kundi, M.; Kollaritsch, H.; Wiedermann, U. Persistence of seroprotection 10 years after primary hepatitis A vaccination in an unselected study population. Vaccine 2007, 25, 927–931. [Google Scholar] [CrossRef]
- Weinberger, B.; Keller, M.; Fischer, K.H.; Stiasny, K.; Neuner, C.; Heinz, F.X.; Grubeck-Loebenstein, B. Decreased antibody titers and booster responses in tick-borne encephalitis vaccinees aged 50–90 years. Vaccine 2010, 28, 3511–3515. [Google Scholar] [CrossRef] [PubMed]
- Haynes, B.F.; Kelsoe, G.; Harrison, S.C.; Kepler, T.B. B-cell-lineage immunogen design in vaccine development with HIV-1 as a case study. Nat. Biotechnol. 2012, 30, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moir, S.; Fauci, A.S. B-cell responses to HIV infection. Immunol. Rev. 2017, 275, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Bonsignori, M.; Zhou, T.; Sheng, Z.; Chen, L.; Gao, F.; Joyce, M.G.; Ozorowski, G.; Chuang, G.Y.; Schramm, C.A.; Wiehe, K.; et al. Maturation Pathway from Germline to Broad HIV-1 Neutralizer of a CD4-Mimic Antibody. Cell 2016, 165, 449–463. [Google Scholar] [CrossRef] [Green Version]
- Bonsignori, M.; Liao, H.X.; Gao, F.; Williams, W.B.; Alam, S.M.; Montefiori, D.C.; Haynes, B.F. Antibody-virus co-evolution in HIV infection: Paths for HIV vaccine development. Immunol. Rev. 2017, 275, 145–160. [Google Scholar] [CrossRef]
- West, A.P., Jr.; Diskin, R.; Nussenzweig, M.C.; Bjorkman, P.J. Structural basis for germ-line gene usage of a potent class of antibodies targeting the CD4-binding site of HIV-1 gp120. Proc. Natl. Acad. Sci. USA. 2012, 109, E2083–E2090. [Google Scholar] [CrossRef] [Green Version]
- Yacoob, C.; Pancera, M.; Vigdorovich, V.; Oliver, B.G.; Glenn, J.A.; Feng, J.; Sather, D.N.; McGuire, A.T.; Stamatatos, L. Differences in Allelic Frequency and CDRH3 Region Limit the Engagement of HIV Env Immunogens by Putative VRC01 Neutralizing Antibody Precursors. Cell Rep. 2016, 17, 1560–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwata, T.; Dehghani, H.; Brown, C.R.; Plishka, R.; Buckler-White, A.; Igarashi, T.; Mattapallil, J.; Roederer, M.; Hirsch, V.M. Infectious molecular clones from a simian immunodeficiency virus-infected rapid-progressor (RP) macaque: Evidence of differential selection of RP-specific envelope mutations in vitro and in vivo. J. Virol. 2006, 80, 1463–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwata, T.; Katsumata, Y.; Takaki, K.; Miura, T.; Igarashi, T. Isolation of potent neutralizing monoclonal antibodies from an SIV-Infected rhesus macaque by phage display. AIDS Res. Hum. Retrovir. 2011, 27, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Sundling, C.; Li, Y.; Huynh, N.; Poulsen, C.; Wilson, R.; O’Dell, S.; Feng, Y.; Mascola, J.R.; Wyatt, R.T.; Karlsson Hedestam, G.B. High-resolution definition of vaccine-elicited B cell responses against the HIV primary receptor binding site. Sci. Transl. Med. 2012, 4, 142ra96. [Google Scholar] [CrossRef] [Green Version]
- Kuwata, T.; Takaki, K.; Yoshimura, K.; Enomoto, I.; Wu, F.; Ourmanov, I.; Hirsch, V.M.; Yokoyama, M.; Sato, H.; Matsushita, S. Conformational epitope consisting of the V3 and V4 loops as a target for potent and broad neutralization of simian immunodeficiency viruses. J. Virol. 2013, 87, 5424–5436. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, S.; Kuwata, T.; Ishii, H.; Sekizuka, T.; Kuroda, M.; Sano, M.; Okazaki, M.; Yamamoto, H.; Shimizu, M.; Matsushita, S.; et al. A Potent anti-Simian Immunodeficiency Virus Neutralizing Antibody Induction Associated with a Germline Immunoglobulin Gene Polymorphism in Rhesus Macaques. J. Virol. 2021, 95, e02455-20. [Google Scholar] [CrossRef]
- Hirsch, V.; Adger-Johnson, D.; Campbell, B.; Goldstein, S.; Brown, C.; Elkins, W.R.; Montefiori, D.C. A molecularly cloned, pathogenic, neutralization-resistant simian immunodeficiency virus, SIVsmE543-3. J. Virol. 1997, 71, 1608–1620. [Google Scholar] [CrossRef] [Green Version]
- Matano, T.; Kano, M.; Nakamura, H.; Takeda, A.; Nagai, Y. Rapid Appearance of Secondary Immune Responses and Protection from Acute CD4 Depletion after a Highly Pathogenic Immunodeficiency Virus Challenge in Macaques Vaccinated with a DNA Prime/Sendai Virus Vector Boost Regimen. J. Virol. 2001, 75, 11891–11896. [Google Scholar] [CrossRef] [Green Version]
- Moriya, C.; Igarashi, H.; Takeda, A.; Tsukamoto, T.; Kawada, M.; Yamamoto, H.; Inoue, M.; Iida, A.; Shu, T.; Hasegawa, M.; et al. Abrogation of AIDS vaccine-induced cytotoxic T-lymphocyte efficacy in vivo due to a change in viral epitope flanking sequences. Microbes Infect 2008, 10, 285–292. [Google Scholar] [CrossRef]
- Science Council of Japan. Guidelines for Proper Conduct of Animal Experiments. Available online: http://www.scj.go.jp/ja/info/kohyo/pdf/kohyo-20-k16-2e.pdf (accessed on 30 April 2021).
- Saito, A.; Kawamoto, Y.; Higashino, A.; Yoshida, T.; Ikoma, T.; Suzaki, Y.; Ami, Y.; Shioda, T.; Nakayama, E.E.; Akari, H. Allele frequency of antiretroviral host factor TRIMCyp in wild-caught cynomolgus macaques (Macaca fascicularis). Front. Microbiol. 2012, 3, 314. [Google Scholar] [CrossRef] [Green Version]
- Mori, K.; Ringler, D.J.; Kodama, T.; Desrosiers, R.C. Complex determinants of macrophage tropism in env of simian immunodeficiency virus. J. Virol. 1992, 66, 2067–2075. [Google Scholar] [CrossRef] [Green Version]
- Marcel, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Aronesty, E. ea-utils: Command-line tools for processing biological sequencing data. Available online: https://github.com/ExpressionAnalysis/ea-utils (accessed on 30 April 2021).
- Lefranc, M.P.; Giudicelli, V.; Ginestoux, C.; Jabado-Michaloud, J.; Folch, G.; Bellahcene, F.; Wu, Y.; Gemrot, E.; Brochet, X.; Lane, J.; et al. IMGT, the international ImMunoGeneTics information system. Nucleic Acids Res. 2009, 37, D1006–D1012. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Ma, N.; Madden, T.L.; Ostell, J.M. IgBLAST: An immunoglobulin variable domain sequence analysis tool. Nucleic Acids Res. 2013, 41, W34–W40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MEGA (Molecular Evolutionary Genetics Analysis). Available online: https://www.megasoftware.net/ (accessed on 30 April 2021).
- Keele, B.F.; Giorgi, E.E.; Salazar-Gonzalez, J.F.; Decker, J.M.; Pham, K.T.; Salazar, M.G.; Sun, C.; Grayson, T.; Wang, S.; Li, H.; et al. Identification and characterization of transmitted and early founder virus envelopes in primary HIV-1 infection. Proc. Natl. Acad. Sci. USA 2008, 105, 7552–7557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ping, L.H.; Joseph, S.B.; Anderson, J.A.; Abrahams, M.R.; Salazar-Gonzalez, J.F.; Kincer, L.P.; Treurnicht, F.K.; Arney, L.; Ojeda, S.; Zhang, M.; et al. Comparison of viral Env proteins from acute and chronic infections with subtype C human immunodeficiency virus type 1 identifies differences in glycosylation and CCR5 utilization and suggests a new strategy for immunogen design. J. Virol. 2013, 87, 7218–7233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, L.M.; Phogat, S.K.; Chan-Hui, P.Y.; Wagner, D.; Phung, P.; Goss, J.L.; Wrin, T.; Simek, M.D.; Fling, S.; Mitcham, J.L.; et al. Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Science 2009, 326, 285–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, L.M.; Simek, M.D.; Priddy, F.; Gach, J.S.; Wagner, D.; Zwick, M.B.; Phogat, S.K.; Poignard, P.; Burton, D.R. A limited number of antibody specificities mediate broad and potent serum neutralization in selected HIV-1 infected individuals. PLoS Pathog. 2010, 6, e1001028. [Google Scholar] [CrossRef] [Green Version]
- Hraber, P.; Seaman, M.S.; Bailer, R.T.; Mascola, J.R.; Montefiori, D.C.; Korber, B.T. Prevalence of broadly neutralizing antibody responses during chronic HIV-1 infection. AIDS 2014, 28, 163–169. [Google Scholar] [CrossRef] [PubMed]
- McGuire, A.T.; Hoot, S.; Dreyer, A.M.; Lippy, A.; Stuart, A.; Cohen, K.W.; Jardine, J.; Menis, S.; Scheid, J.F.; West, A.P.; et al. Engineering HIV envelope protein to activate germline B cell receptors of broadly neutralizing anti-CD4 binding site antibodies. J. Exp. Med. 2013, 210, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Jardine, J.G.; Kulp, D.W.; Havenar-Daughton, C.; Sarkar, A.; Briney, B.; Sok, D.; Sesterhenn, F.; Ereño-Orbea, J.; Kalyuzhniy, O.; Deresa, I.; et al. HIV-1 broadly neutralizing antibody precursor B cells revealed by germline-targeting immunogen. Science 2016, 351, 1458–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macaque | n | VH3.33 Allele b | ||
|---|---|---|---|---|
| VI/VI | VI/ET | ET/ET | ||
| Rhesus | 28 | 13 | 4 | 11 |
| Cynomolgus | 24 | 24 | 0 | 0 |
| Macaques | VH3.33 Allele a | TRIM5 Allele b | Vaccination c | ||
|---|---|---|---|---|---|
| #1 | VI | VI | TFP | Cyp | unvaccinated |
| #2 | VI | ET | TFP | Cyp | unvaccinated |
| #3 | ET | ET | TFP | Cyp | unvaccinated |
| #4 | VI | ET | TFP | Cyp | vaccinated |
| #5 | ET | ET | TFP | TFP | vaccinated |
| #6 | ET | ET | Q | Q | vaccinated |
| #7 | VI | VI | Cyp | Cyp | vaccinated |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nomura, Y.; Matsuoka, S.; Okazaki, M.; Kuwata, T.; Matano, T.; Ishii, H. Neutralizing Antibody Induction Associated with a Germline Immunoglobulin Gene Polymorphism in Neutralization-Resistant SIVsmE543-3 Infection. Viruses 2021, 13, 1181. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061181
Nomura Y, Matsuoka S, Okazaki M, Kuwata T, Matano T, Ishii H. Neutralizing Antibody Induction Associated with a Germline Immunoglobulin Gene Polymorphism in Neutralization-Resistant SIVsmE543-3 Infection. Viruses. 2021; 13(6):1181. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061181
Chicago/Turabian StyleNomura, Yuto, Saori Matsuoka, Midori Okazaki, Takeo Kuwata, Tetsuro Matano, and Hiroshi Ishii. 2021. "Neutralizing Antibody Induction Associated with a Germline Immunoglobulin Gene Polymorphism in Neutralization-Resistant SIVsmE543-3 Infection" Viruses 13, no. 6: 1181. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061181