
R-BPMV-Mediated Resistance to Bean pod mottle virus in Phaseolus vulgaris L. Is Heat-Stable but Elevated Temperatures Boost Viral Infection in Susceptible Genotypes

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
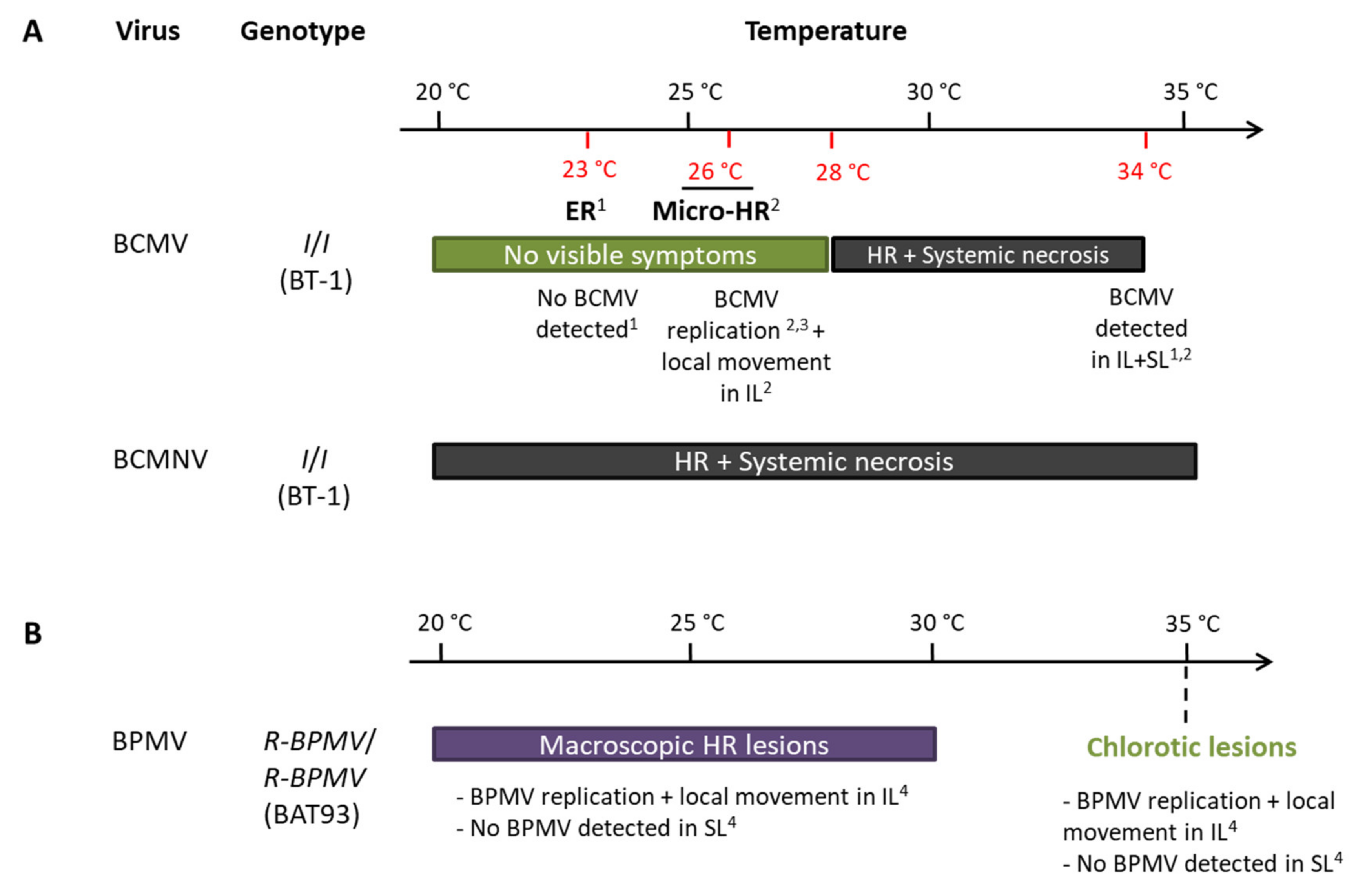{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Common Bean Material and Growing Conditions
2.2. Viral Material
2.3. Viral Rub-Inoculation of P. vulgaris Plants and High-Temperature Assays
2.4. Cell Death Assays
2.5. Detection of GFP Fluorescence in Planta
2.6. RNA Isolation and RT-PCR Analyses
3. Results
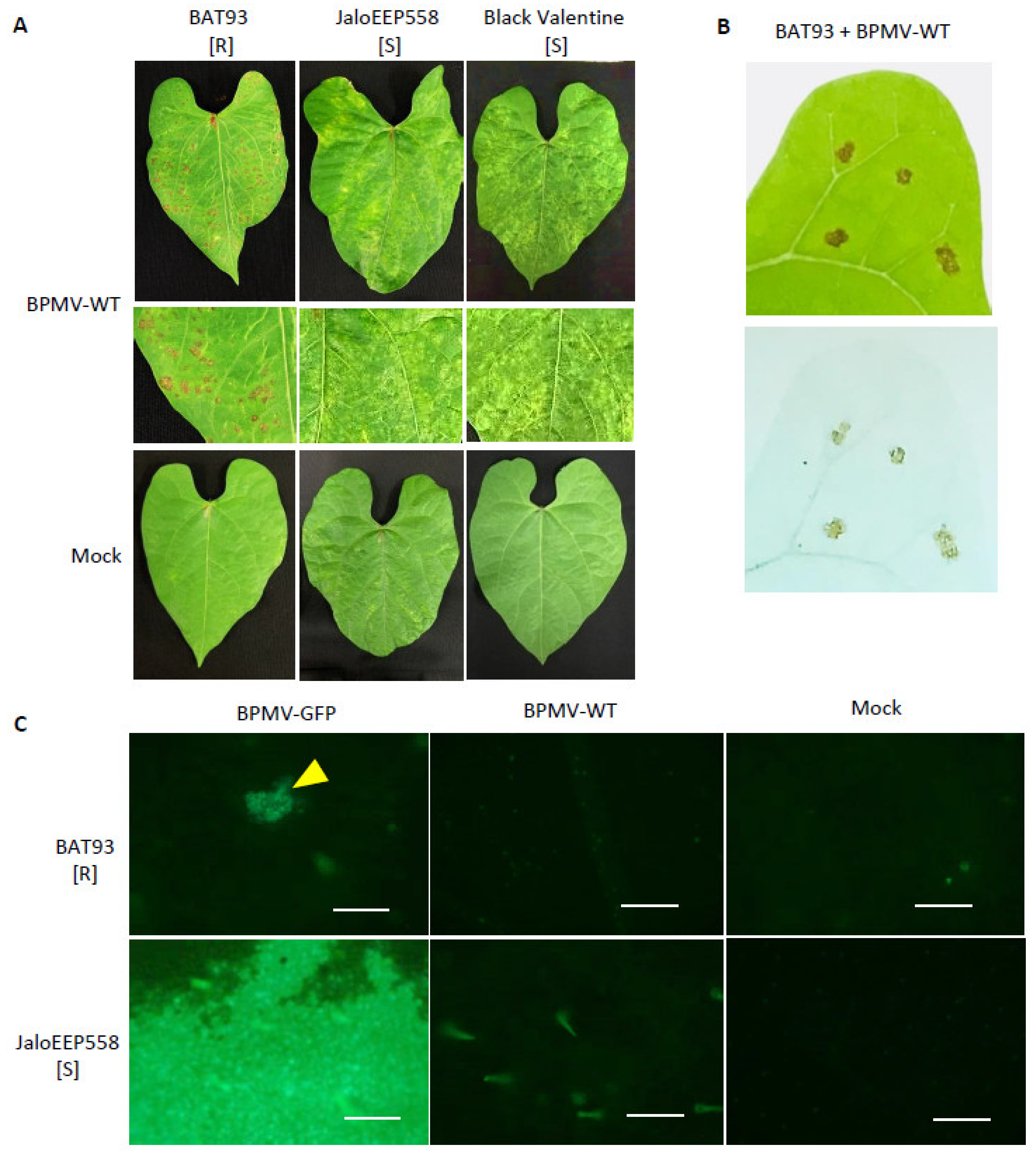
3.1. R-BPMV-Mediated Resistance Is Associated with Local HR Lesions at 20 °C Where BPMV Is Able to Multiply in a First Step
3.2. The R-BPMV Gene Segregates as an Incompletely Dominant Gene
3.3. At 25 and 30 °C, R-BPMV Induces More Expanded Local HR Lesions
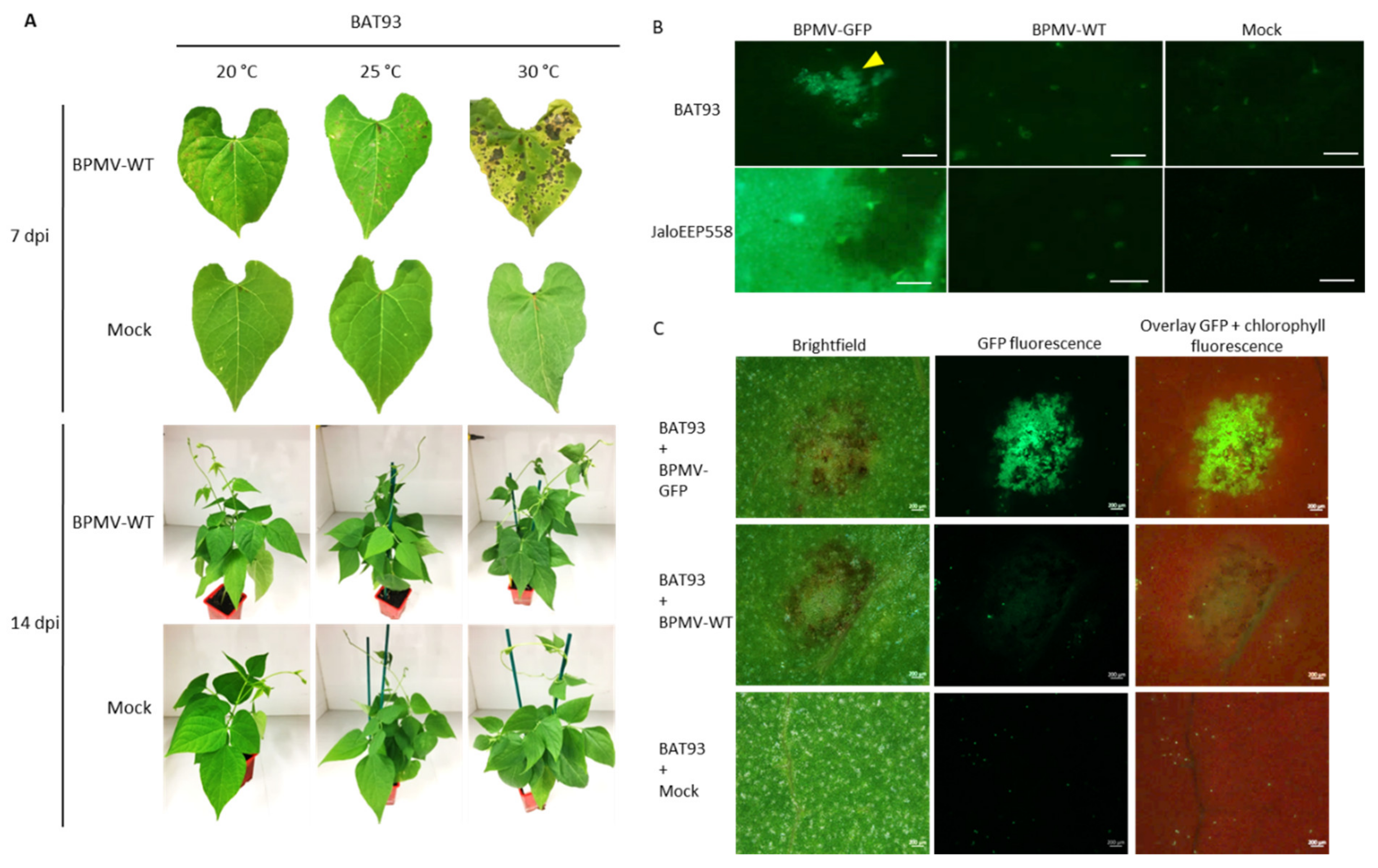
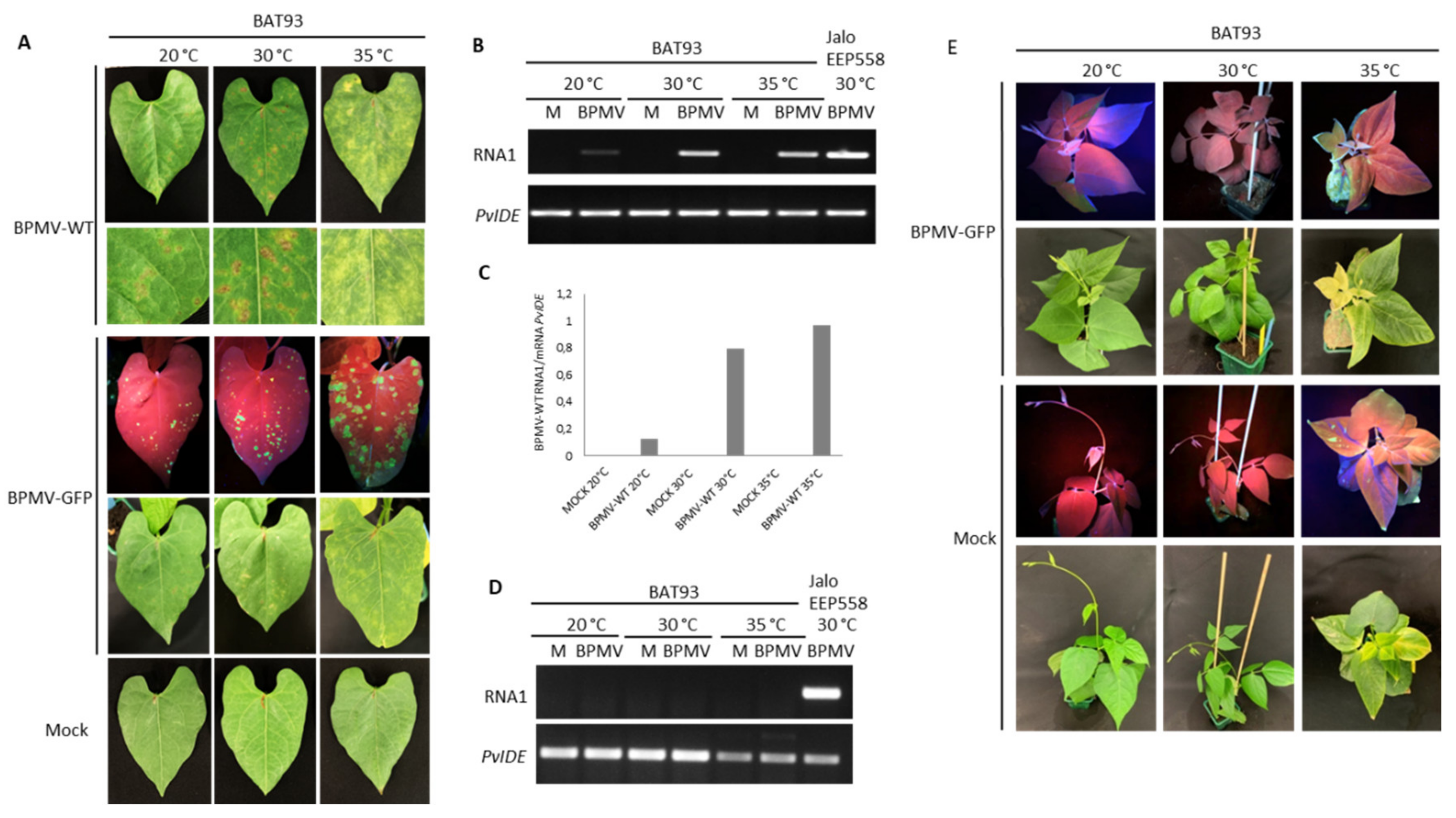
3.4. The Resistance Mediated by R-BPMV Is Heat-Stable up to 35 °C in BAT93, But Local HR Lesions Are Replaced by Chlorotic Lesions
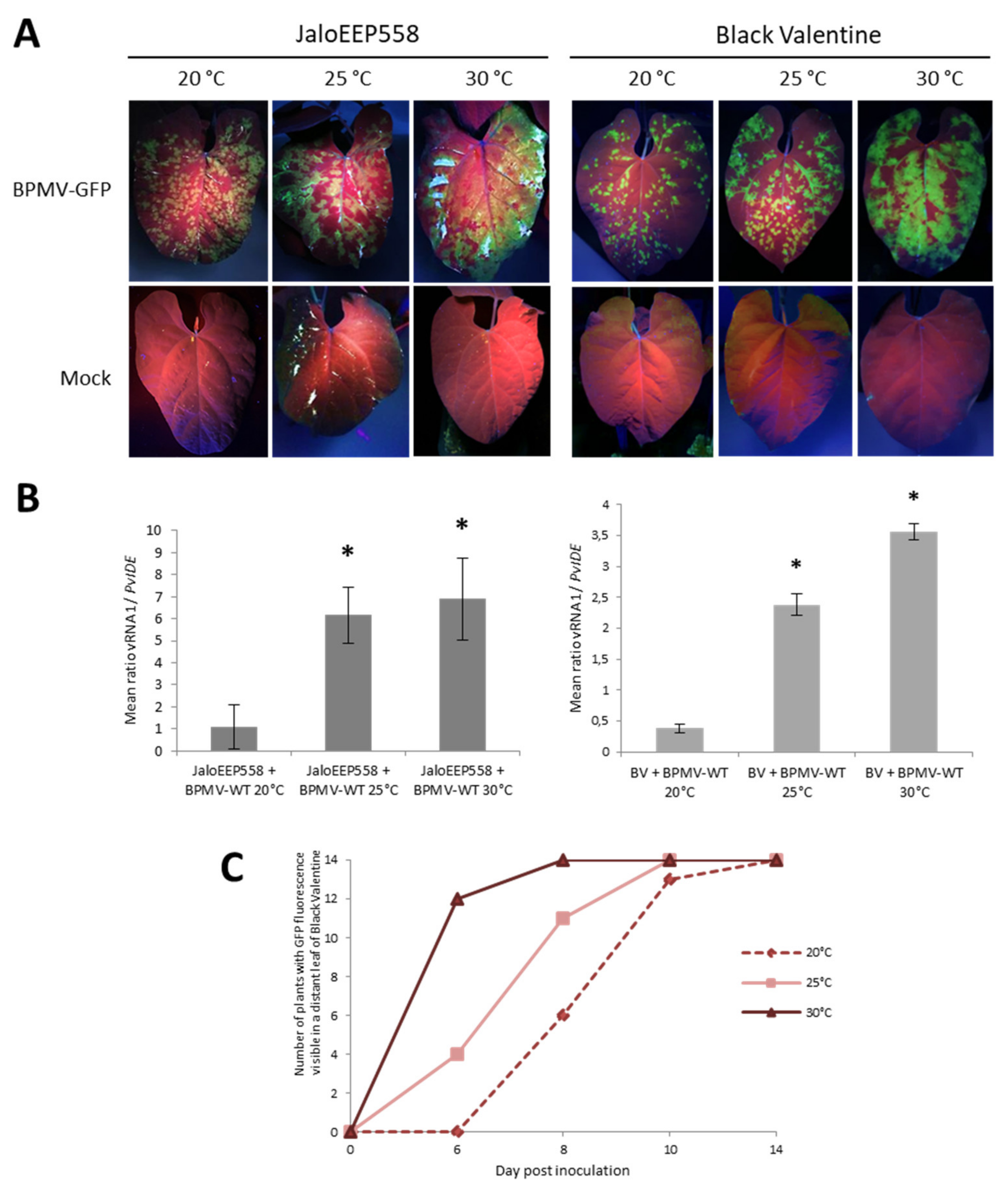
3.5. Elevated Temperatures Boost BPMV Infection in Susceptible Genotypes
3.6. BT-1 and BT-2, Two Near Isogenic Lines for the I. locus, Are Both Resistant to BPMV Systemic Movement
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abreviations
| Avr factor | avirulence factor |
| CO2 | carbon dioxide |
| dpi | day(s) post-inoculation |
| ETI | effector-triggered immunity |
| HR | hypersensitive reaction |
| NLR | nucleotide-binding domain leucin-rich repeat containing receptors |
| ppm | parts per million |
| R | resistance gene/protein |
References
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef]
- Sastry, K.S.; Zitter, T.A. Plant Virus and Viroid Diseases in the Tropics. Volume 2: Epidemiology and Management; Springer: Dordrecht, The Netherlands, 2014. [Google Scholar]
- Amari, K.; Niehl, A. Nucleic acid-mediated PAMP-triggered immunity in plants. Curr. Opin. Virol. 2020, 42, 32–39. [Google Scholar] [CrossRef]
- Ratcliff, F.; Harrison, B.D.; Baulcombe, D.C. A similarity between viral defense and gene silencing in plants. Science 1997, 276, 1558–1560. [Google Scholar] [CrossRef] [PubMed]
- Kørner, C.; Klauser, D.; Niehl, A.; Domínguez-Ferreras, A.; Chinchilla, D.; Boller, T.; Heinlein, M.; Hann, D.R. The immunity regulator BAK1 contributes to resistance against diverse RNA viruses. Mol. Plant Microbe Interact. 2013, 26, 1271–1280. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.; Park, Y.S.; Lee, S.; Song, G.C.; Ryu, C.M. Bacterial RNAs activate innate immunity in Arabidopsis. New Phytol. 2016, 209, 785–797. [Google Scholar] [CrossRef]
- Niehl, A.; Wyrsch, I.; Boller, T.; Heinlein, M. Double-stranded RNAs induce a pattern-triggered immune signaling pathway in plants. New Phytol. 2016, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csorba, T.; Kontra, L.; Burgyán, J. Viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479–480, 85–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zvereva, A.S.; Golyaev, V.; Turco, S.; Gubaeva, E.G.; Rajeswaran, R.; Schepetilnikov, M.V.; Srour, O.; Ryabova, L.A.; Boller, T.; Pooggin, M.M. Viral protein suppresses oxidative burst and salicylic acid-dependent autophagy and facilitates bacterial growth on virus-infected plants. New Phytol. 2016, 211, 1020–1034. [Google Scholar] [CrossRef] [Green Version]
- Nicaise, V.; Candresse, T. Plum pox virus capsid protein suppresses plant pathogen-associated molecular pattern (PAMP)-triggered immunity. Mol. Plant Pathol. 2017, 18, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Wei, M.; Li, G.; Lei, R.; Qiu, Y.; Wang, C.; Li, Z.H.; Zhu, S. The cucumber mosaic virus movement protein suppresses PAMP-triggered immune responses in Arabidopsis and tobacco. Biochem. Biophys. Res. Commun. 2018, 498, 395–401. [Google Scholar] [CrossRef]
- Palukaitis, P.; Yoon, J.Y. R gene mediated defense against viruses. Curr. Opin. Virol. 2020, 45, 1–7. [Google Scholar] [CrossRef]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1955, 1971, 275–296. [Google Scholar] [CrossRef]
- Gouveia, B.C.; Calil, I.P.; Machado, J.P.B.; Santos, A.A.; Fontes, E.P.B. Immune Receptors and Co-receptors in Antiviral Innate Immunity in Plants. Front. Microbiol. 2017, 7, 2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ronde, D.; Butterbach, P.; Kormelink, R. Dominant resistance against plant viruses. Front. Plant Sci. 2014, 5, 307. [Google Scholar] [CrossRef] [Green Version]
- Kou, Y.; Wang, S. Broad-spectrum and durability: Understanding of quantitative disease resistance. Curr. Opin. Plant Biol. 2010, 13, 181–185. [Google Scholar] [CrossRef]
- Schmitt-Keichinger, C. Manipulating cellular factors to combat viruses: A case study from the plant eukaryotic translation initiation factors eIF4. Front. Microbiol. 2019, 10, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatesh, J.; Kang, B.C. Current views on temperature-modulated R gene-mediated plant defense responses and tradeoffs between plant growth and immunity. Curr. Opin. Plant Biol. 2019, 50, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Trebicki, P. Climate change and plant virus epidemiology. Virus Res. 2020, 286, 198059. [Google Scholar] [CrossRef]
- Tenllado, F.; Canto, T. Effects of a changing environment on the defenses of plants to viruses. Curr. Opin. Virol. 2020, 42, 40–46. [Google Scholar] [CrossRef]
- Desaint, H.; Aoun, N.; Deslandes, L.; Vailleau, F.; Roux, F.; Berthomé, R. Fight hard or die trying: When plants face pathogens under heat stress. New Phytol. 2021, 229, 712–734. [Google Scholar] [CrossRef] [PubMed]
- Weststeijn, E. Lesion growth and virus localization in leaves of Nicotiana tabacum cv. Xanthi nc. after inoculation with tobacco mosaic virus and incubation alternately at 22 °C and 32 °C. Physiol. Plant Pathol. 1981, 18, 357–368. [Google Scholar] [CrossRef]
- Chung, B.N.; Lee, J.H.; Kang, B.C.; Koh, S.W.; Joa, J.H.; Choi, K.S.; Ahn, J.J. HR-mediated defense response is overcome at high temperatures in capsicum species. Plant Pathol. J. 2018, 34, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Richard, M.M.S.; Knip, M.; Aalders, T.; Beijaert, M.S.; Takken, F.L.W. Unlike Many Disease Resistances, Rx1-Mediated Immunity to Potato Virus X Is Not Compromised at Elevated Temperatures. Front. Genet. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- Kabbage, M.; Kessens, R.; Bartholomay, L.C.; Williams, B. The Life and Death of a Plant Cell. Annu. Rev. Plant Biol. 2017, 68, 375–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.S.; del Toro, F.; Tenllado, F.; Canto, T.; Chung, B.N. A model to explain temperature dependent systemic infection of potato plants by Potato virus Y. Plant Pathol. J. 2017, 33, 206–211. [Google Scholar] [CrossRef] [Green Version]
- Nancarrow, N.; Constable, F.E.; Finlay, K.J.; Freeman, A.J.; Rodoni, B.C.; Trebicki, P.; Vassiliadis, S.; Yen, A.L.; Luck, J.E. The effect of elevated temperature on barley yellow dwarf virus-PAV in wheat. Virus Res. 2014, 186, 97–103. [Google Scholar] [CrossRef]
- Kido, K.; Tanaka, C.; Mochizuki, T.; Kubota, K.; Ohki, T.; Ohnishi, J.; Knight, L.M.; Tsuda, S. High temperatures activate local viral multiplication and cell-to-cell movement of Melon necrotic spot virus but restrict expression of systemic symptoms. Phytopathology 2008, 98, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.N.; Choi, K.S.; Ahn, J.J.; Joa, J.H.; Do, K.S.; Park, K.S. Effects of temperature on systemic infection and symptom expression of turnip mosaic virus in Chinese cabbage (Brassica campestris). Plant Pathol. J. 2015, 31, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Del Toro, F.J.; Aguilar, E.; Hernández-Walias, F.J.; Tenllado, F.; Chung, B.N.; Canto, T. High temperature, high ambient CO2 affect the interactions between three positive-sense RNA viruses and a compatible host differentially, but not their silencing suppression efficiencies. PLoS ONE 2015, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Dahal, G.; Hughes, J.D.; Thottappilly, G.; Lockhart, B.E.L. Effect of temperature on symptom expression and reliability of banana streak badnavirus detection in naturally infected plantain and banana (Musa spp.). Plant Dis. 1998, 82, 16–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chellappan, P.; Vanitharani, R.; Ogbe, F.; Fauquet, C.M. Effect of temperature on geminivirus-induced RNA silencing in plants. Plant Physiol. 2005, 138, 1828–1841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broughton, W.J.; Hernandez, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef] [Green Version]
- Beebe, S.; Ramirez, J.; Jarvis, A.; Rao, I.M.; Mosquera, G.; Bueno, J.M.; Blair, M.W. Genetic Improvement of Common Beans and the Challenges of Climate Change. Crop Adapt. Clim. Chang. 2011, 356–369. [Google Scholar] [CrossRef]
- Julian R—Climate Change and Bean Production. Available online: https://pt.slideshare.net/ciatdapa/julian-r-climate-change-and-bean-production/2 (accessed on 25 April 2021).
- Omae, H.; Kumar, A.; Kashiwaba, K.; Shono, M. Adaptation to high temperature and water deficit in the common bean (Phaseolus vulgaris L.) during the reproductive period. J. Bot. 2012. [Google Scholar] [CrossRef] [Green Version]
- Suarez, J.C.; Polania, J.A.; Contreras, A.T.; Rodriguez, L.; Machado, L.; Ordonez, C.; Beebe, S.; Rao, I. Adaptation of common bean lines to high temperature conditions: Genotypic differences in phenological and agronomic performance Adaptation of common bean lines to high temperature conditions: Genotypic differences in phenological and agronomic performan. Euphytica 2020, 216, 28. [Google Scholar] [CrossRef]
- Meziadi, C.; Blanchet, S.; Geffroy, V.; Pflieger, S. Genetic resistance against viruses in Phaseolus vulgaris L.: State of the art and future prospects. Plant Sci. 2017, 265, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Morales, F.J.; Castano, M. Increased disease severity induced by some comoviruses in bean genotypes possessing monogenic dominant resistance to Bean common mosaic potyvirus. Plant Dis. 1992, 76, 570–573. [Google Scholar] [CrossRef]
- Morales, F.J.; Singh, S.P. Inheritance of the mosaic and necroses reactions induced by bean severe mosaic comoviruses in Phaseolus vulgaris L. Euphytica 1997, 93, 223–226. [Google Scholar] [CrossRef]
- Pflieger, S.; Blanchet, S.; Meziadi, C.; Richard, M.M.S.; Thareau, V.; Mary, F.; Mazoyer, C.; Geffroy, V. The “ one-step ” Bean pod mottle virus (BPMV)—Derived vector is a functional genomics tool for efficient overexpression of heterologous protein, virus-induced gene silencing and genetic mapping of BPMV R-gene in common bean ( Phaseolus vulgaris L.). BMC Plant Biol. 2014, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Provvidenti, R. Two useful selections of the bean cultivar Black Turtle Soup for viral identification. Bean Improv. Coop. Annu. Rep. 1983, 26, 73–75. [Google Scholar]
- Zhang, C.; Bradshaw, J.D.; Whitham, S.A.; Hill, J.H. The Development of an Efficient Multipurpose Bean Pod Mottle Virus Viral Vector Set for Foreign Gene Expression and RNA silencing. Plant Physiol. 2010, 153, 52–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pflieger, S.; Blanchet, S.; Meziadi, C.; Richard, M.M.S.; Geffroy, V. Bean pod mottle virus (BPMV) viral inoculation procedure in common bean (Phaseolus vulgaris L.). Bio-Protocol 2015, 5, e1524. [Google Scholar] [CrossRef]
- Richard, M.M.S.; Gratias, A.; Alvarez Diaz, J.C.; Thareau, V.; Pflieger, S.; Meziadi, C.; Blanchet, S.; Marande, W.; Bitocchi, E.; Papa, R.; et al. A common bean truncated CRINKLY4 kinase controls gene-for-gene resistance to the fungus Colletotrichum lindemuthianum. J. Exp. Bot. 2021, 72, 3569–3581. [Google Scholar] [CrossRef]
- Ali, M.A. Genetics of resistance to the common Bean mosaic virus in the bean (Phaseolus vulgaris L). Phytopathology 1950, 40, 69–79. [Google Scholar]
- Bos, L. Bean common mosaic virus. C. Descr. Plant Viruses 1971, 73. [Google Scholar]
- Drijfhout, E. Genetic Interaction between Phaseolus vulgaris and bean Common Mosaic Virus with Implications for Strain Identification and Breeding for Resistance; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1978. [Google Scholar]
- Whitham, S.; McCormick, S.; Baker, B. The N gene of tobacco confers resistance to tobacco mosaic virus in transgenic tomato. Proc. Natl. Acad. Sci. USA 1996, 93, 8776–8781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collmer, C.W.; Marston, M.F.; Taylor, J.C.; Jahn, M. The I gene of bean: A dosage-dependent allele conferring extreme resistance, hypersensitive resistance, or spreading vascular necrosis in response to the potyvirus Bean common mosaic virus. Mol. Plant Microbe Interact. 2000, 13, 1266–1270. [Google Scholar] [CrossRef] [Green Version]
- Cadle-Davidson, M.M.; Jahn, M.M. Resistance conferred against bean common mosaic virus by the incompletely dominant I locus of Phaseolus vulgaris is active at the single cell level. Arch. Virol. 2005, 150, 2601–2608. [Google Scholar] [CrossRef]
- Cadle-Davidson, M.M.; Jahn, M.M. Patterns of accumulation of Bean common mosaic virus in Phaseolus vulgaris genotypes nearly isogenic for the I locus. Ann. Appl. Biol. 2006, 148, 179–185. [Google Scholar] [CrossRef]
- Tomita, R.; Sekine, K.-T.; Mizumoto, H.; Sakamoto, M.; Murai, J.; Kiba, A.; Hikichi, Y.; Suzuki, K.; Kobayashi, K. Genetic Basis for the Hierarchical Interaction between Tobamovirus spp. and L Resistance Gene Alleles from Different Pepper Species. Mol. Plant Microbe Interact. 2011, 24, 108–117. [Google Scholar] [CrossRef] [Green Version]
- Veremis, J.C.; van Heusden, A.W.; Roberts, P.A. Mapping a novel heat-stable resistance to Meloidogyne in Lycopersicon peruvianum. Theor. Appl. Genet. 1999, 98, 274–280. [Google Scholar] [CrossRef]
- Ammiraju, J.S.S.; Veremis, J.C.; Huang, X.; Roberts, P.A.; Kaloshian, I. The heat-stable root-knot nematode resistance gene Mi-9 from Lycopersicon peruvianum is localized on the short arm of chromosome 6. Theor. Appl. Genet. 2003, 106, 478–484. [Google Scholar] [CrossRef]
- Jablonska, B.; Ammiraju, J.S.S.; Bhattarai, K.K.; Mantelin, S.; de Ilarduya, O.M.; Roberts, P.A.; Kaloshian, I. The Mi-9 gene from Solanum arcanum conferring heat-stable resistance to root-knot nematodes is a homolog of Mi-1. Plant Physiol. 2007, 143, 1044–1054. [Google Scholar] [CrossRef] [Green Version]
- Djian-Caporalino, C.; Pijarowski, L.; Januel, A.; Lefebvre, V.; Daubèze, A.; Palloix, A.; Dalmasso, A.; Abad, P. Spectrum of resistance to root-knot nematodes and inheritance of heat-stable resistance in in pepper (Capsicum annuum L.). Theor. Appl. Genet. 1999, 99, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Djian-Caporalino, C.; Pijarowski, L.; Fazari, A.; Samson, M.; Gaveau, L.; O’Byrne, C.; Lefebvre, V.; Caranta, C.; Palloix, A.; Abad, P. High-resolution genetic mapping of the pepper (Capsicum annuum L.) resistance loci Me3 and Me4 conferring heat-stable resistance to root-knot nematodes (Meloidogyne spp.). Theor. Appl. Genet. 2001, 103, 592–600. [Google Scholar] [CrossRef]
- Bradshaw, J.; Ramsey, G. Utilisation of the Commonwealth Potato Collection in potato breeding. Euphytica 2005, 146, 9–19. [Google Scholar] [CrossRef]
- Solomon-Blackburn, R.; Bradshaw, J. Resistance to Potato virus Y in a Multitrait Potato Breeding Scheme without Direct Selection in Each Generation. Potato Res. 2007, 50, 87–95. [Google Scholar] [CrossRef]
- Moury, B.; Selassie, K.G.; Marchoux, G.; Daub, A.; Palloix, A. High temperature effects on hypersensitive resistance to Tomato Spotted Wilt Tospovirus (TSWV) in pepper (Capsicum chinense Jacq.). Eur. J. Plant Pathol. 1998, 104, 489–498. [Google Scholar] [CrossRef]
- De Carvalho, L.; Benda, N.; Vaughan, M.; Cabrera, A.; Hung, K.; Cox, T.; Abdo, Z.; Allen, L.; Teal, P. Mi-1-Mediated Nematode Resistance in Tomatoes is Broken by Short-Term Heat Stress but Recovers Over Time. J. Nematol. 2015, 47, 133–140. [Google Scholar]
- Romero, A.M.; Kousik, C.S.; Ritchie, D.F. Temperature Sensitivity of the Hypersensitive Response of Bell Pepper to Xanthomonas axonopodis pv. vesicatoria. Phytopathology 2002, 92, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Takken, F.; Joosten, M.; de Wit, P. Specific recognition of AVR4 and AVR9 results in distinct patterns of hypersensitive cell death in tomato, but similar patterns of defence-related gene expression. Mol. Plant Pathol. 2001, 2, 77–86. [Google Scholar] [CrossRef] [PubMed]
- De Jong, C.F.; Takken, F.L.; Cai, X.; de Wit, P.J.; Joosten, M.H. Attenuation of Cf-Mediated Defense Responses at Elevated Temperatures Correlates With a Decrease in Elicitor-Binding Sites. Mol. Plant Microbe Interact. 2002, 15, 1040–1049. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Gao, X.; Feng, B.; Sheen, J.; Shan, L.; He, P. Differential temperature operation of plant immune responses. Nat. Commun. 2013, 4, 2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menna, A.; Nguyen, D.; Guttman, D.S.; Desveaux, D. Elevated temperature differentially influences effector-triggered immunity outputs in Arabidopsis. Front. Plant Sci. 2015, 6, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoun, N.; Tauleigne, L.; Lonjon, F.; Deslandes, L.; Vailleau, F.; Roux, F.; Berthomé, R. Quantitative disease resistance under elevated temperature: Genetic basis of new resistance mechanisms to Ralstonia solanacearum. Front. Plant Sci. 2017, 8, 1387. [Google Scholar] [CrossRef]
- Yang, S.; Hua, J. A haplotype-specific Resistance gene regulated by BONZAI1 mediates temperature-dependent growth control in Arabidopsis. Plant Cell 2004, 16, 1060–1071. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Qian, W.; Hua, J. Temperature modulates plant defense responses through NB-LRR proteins. PLoS Pathog. 2010, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mang, H.G.; Qian, W.; Zhu, Y.; Qian, J.; Kang, H.G.; Klessig, D.F.; Hua, J. Abscisic acid deficiency antagonizes high-temperature inhibition of disease resistance through enhancing nuclear accumulation of resistance proteins SNC1 and RPS4 in Arabidopsis. Plant Cell 2012, 24, 1271–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fesenko, I.; Spechenkova, N.; Mamaeva, A.; Makhotenko, A.V.; Love, A.J.; Kalinina, N.O.; Taliansky, M. Role of the methionine cycle in the temperature-sensitive responses of potato plants to potato virus Y. Mol. Plant Pathol. 2021, 22, 77–91. [Google Scholar] [CrossRef]
- Amari, K.; Huang, C.; Heinlein, M. Potential Impact of Global Warming on Virus Propagation in Infected Plants and Agricultural Productivity. Front. Plant Sci. 2021, 12, 649768. [Google Scholar] [CrossRef]
- Whitham, S.A.; Yang, C.; Goodin, M.M. Global impact: Elucidating plant responses to viral infection. Mol. Plant Microbe Interact. 2006, 19, 1207–1215. [Google Scholar] [CrossRef] [Green Version]
- Lewsey, M.; Palukaitis, P.; Carr, J. Plant–Virus Interactions: Defence and Counter-Defence. In Annual Plant Reviews Volume 34: Molecular Aspects of Plant Disease Resistance; Wiley-Blackwell: Oxford, UK, 2009; pp. 134–176. [Google Scholar] [CrossRef]
- Baebler, Š.; Witek, K.; Petek, M.; Stare, K.; Tušek-Žnidarič, M.; Pompe-Novak, M.; Renaut, J.; Szajko, K.; Strzelczyk-Zyta, D.; Marczewski, W.; et al. Salicylic acid is an indispensable component of the Ny-1 resistance-gene-mediated response against Potato virus y infection in potato. J. Exp. Bot. 2014, 65, 1095–1109. [Google Scholar] [CrossRef]
- Chandra-Shekara, A.; Navarre, D.; Kachroo, A.; Kang, H.; Klessig, D.; Kachroo, P. Signaling requirements and role of salicylic acid in HRT—And rrt -mediated resistance to turnip crinkle virus in Arabidopsis. Plant J. 2004, 40, 647–659. [Google Scholar] [CrossRef]
- Lukan, T.; Baebler, S.; Pompe-Novak, M.; Gucek, K.; Zagoršcak, M.; Coll, A.; Gruden, K. Cell Death Is Not Sufficient for the Restriction of Potato Virus Y Spread in Hypersensitive Response-Conferred Resistance in Potato. Front. Plant Sci. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.; Yanan, L.; Lijuan, C.; Lisha, Z.; Han, R.; Honghui, L.; Dehui, X. Temperature dependent defence of Nicotiana tabacum against Cucumber mosaic virus and recovery occurs with the formation of dark green islands. J. Plant Biol. 2016, 59, 293–301. [Google Scholar] [CrossRef]
- Makarova, S.; Makhotenko, A.; Spechenkova, N.; Love, A.J.; Kalinina, N.O.; Taliansky, M. Interactive Responses of Potato (Solanum tuberosum L.) Plants to Heat Stress and Infection With Potato Virus Y. Front. Microbiol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, M.; Desvoyes, B.; Turina, M.; Noad, R.; Scholthof, H.B. Genetic dissection of tomato bushy stunt virus p19-protein-mediated host-dependent symptom induction and systemic invasion. Virology 2000, 266, 79–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cawly, J.; Cole, A.B.; Király, L.; Qiu, W.; Schoelz, J.E. The plant gene CCD1 selectively blocks cell death during the hypersensitive response to cauliflower mosaic virus infection. Mol. Plant Microbe Interact. 2005, 18, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Masuda, K.; Naito, S.; Meshi, T.; Ishikawa, M. An inhibitor of viral RNA replication is encoded by a plant resistance gene. Proc. Natl. Acad. Sci. USA 2007, 104, 13833–13838. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, K.; Hashimoto, M.; Ozeki, J.; Yamaji, Y.; Maejima, K.; Senshu, H.; Himeno, M.; Okano, Y.; Kagiwada, S.; Namba, S. Viral-Induced systemic necrosis in plants involves both programmed cell death and the inhibition of viral multiplication, which are regulated by independent pathways. Mol. Plant Microbe Interact. 2010, 23, 283–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.P.; Bhattacharjee, S.; Klessig, D.F.; Moffett, P. Systemic acquired resistance is induced by R gene-mediated responses independent of cell death. Mol. Plant Pathol. 2010, 11, 155–160. [Google Scholar] [CrossRef]
- Hafez, Y.M.; Bacsó, R.; Király, Z.; Künstler, A.; Király, L. Up-regulation of antioxidants in tobacco by low concentrations of H 2O 2 suppresses necrotic disease symptoms. Phytopathology 2012, 102, 848–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, S.; Obinata, A.; Takahashi, H. WRKY70 interacting with RCY1 disease resistance protein is required for resistance to Cucumber mosaic virus in Arabidopsis thaliana. Physiol. Mol. Plant Pathol. 2014, 85, 8–14. [Google Scholar] [CrossRef]
- Harris, J.M.; Balint-kurti, P.; Bede, J.C.; Day, B.; Gold, S.; Goss, E.M.; Grenville-briggs, L.J.; Jones, K.M.; Wang, A.; Wang, Y.; et al. What are the Top 10 Unanswered Questions in Molecular Plant-Microbe Interactions? Mol. Plant Microbe Interact. 2020, X, 1–12. [Google Scholar] [CrossRef]
- Zheng, C.; Chen, P.; Gergerich, R. Characterization of Resistance to Soybean mosaic virus in Diverse Soybean Germplasm. Crop Sci. 2005, 45, 2503–2509. [Google Scholar] [CrossRef]
- Meziadi, C.; (IPS2, Orsay, France). Personal communication, 2021.
- Vallejos, C.E.; Astua-monge, G.; Jones, V.; Plyler, T.R.; Sakiyama, N.S.; Mackenzie, S.A. Genetic and Molecular Characterization of the I Locus of Phaseolus vulgaris. Genetics 2006, 172, 1229–1242. [Google Scholar] [CrossRef] [Green Version]
- Nodari, R.O.; Tsai, S.M.; Gilbertson, R.L.; Gepts, P. Towards an integrated linkage map of common bean. 2. Development of an RFLP-based linkage map. Theor. Appl. Genet. 1993, 85, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Kyle, M.M.; Provvidenti, R. Inheritance of resistance to potato y viruses in Phaseolus vulgaris L: 1. Two independent genes for resistance to watermelon mosaic virus-2. Theor. Appl. Genet. 1987, 74, 595–600. [Google Scholar] [CrossRef]
- Provvidenti, R. Inheritance of resistance to watermelon mosaic virus 2 in Phaseolus vulgaris. Phytopathology 1974, 64, 1448–1450. [Google Scholar] [CrossRef]
- Provvidenti, R.; Gonsalves, D.; Taiwo, M.A. Inheritance of resistance to Blackeye cowpea mosaic and Cowpea aphid-borne mosaic viruses in Phaseolus vulgaris. J. Hered. 1983, 74, 60–61. [Google Scholar] [CrossRef] [Green Version]
- Provvidenti, R.; Gonsalves, D.; Ranalli, P. Inheritance of resistance to soybean mosaic virus in Phaseolus vulgaris. J. Hered. 1982, 73, 302–303. [Google Scholar] [CrossRef] [Green Version]
- Kyle, M.M.; Provvidenti, R. Inheritance of resistance to potyviruses in Phaseolus vulgaris L. 2. Linkage relations and utility of a dominant gene for lethal systemic necrosis to Soybean mosaic virus. Theor. Appl. Genet. 1993, 86, 189–196. [Google Scholar] [CrossRef]
- Provvidenti, R.; Chirco, E.M. Inheritance of resistance to peanut mottle virus in Phaseolus vulgaris. J. Hered. 1987, 78, 402–403. [Google Scholar]
- Li, R.H.; Zettler, F.W.; Elliott, M.S.; Petersen, M.A.; Still, P.E.; Baker, C.A.; Mink, G.I. A strain of Peanut Mottle Virus Seedborne in Bambarra Groundnut. Plant Dis. 1991, 75, 130–133. [Google Scholar] [CrossRef]
- Fisher, M.L.; Kyle, M.M. Inheritance of resistance to potyviruses in Phaseolus vulgaris L. III. Cosegregation of phenotypically similar dominant responses to nine potyviruses. Theor. Appl. Genet. 1994, 89, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Hart, J.P.; Griffiths, P.D. Genotyping-by-Sequencing Enabled Mapping and Marker Development for the By-2 Potyvirus Resistance Allele in Common Bean. Plant Genome 2015, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickson, M.H.; Natti, J.J. Inheritance of resistance of Phaseolus vulgaris to bean yellow mosaic virus. Phytopathology 1968, 58, 1450. [Google Scholar]
- Bello, M.H.; Moghaddam, S.M.; Massoudi, M.; Mcclean, P.E.; Cregan, P.B.; Miklas, P.N. Application of in silico bulked segregant analysis for rapid development of markers linked to Bean common mosaic virus resistance in common bean. BMC Genom. 2014, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Chung, B.N.; Canto, T.; Tenllado, F.; Choi, K.S.; Joa, J.H.; Ahn, J.J.; Kim, C.H.; Do, K.S. The effects of high temperature on infection by Potato virus Y, Potato virus A, and Potato leafroll virus. Plant Pathol. J. 2016, 32, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, E.; Allende, L.; Del Toro, F.J.; Chung, B.N.; Canto, T.; Tenllado, F. Effects of elevated CO2 and temperature on pathogenicity determinants and virulence of potato virus X/Potyvirus-Associated synergism. Mol. Plant-Microbe Interact. 2015, 28, 1364–1373. [Google Scholar] [CrossRef] [Green Version]
- Szittya, G.; Silhavy, D.; Molnar, A.; Havelda, Z.; Lovas, A.; Lakatos, L.; Banfaldi, Z.; Burgyan, J. Low temperature inhibits RNA silencing-mediated defence by the control of siRNA generation. EMBO J. 2003, 22, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Toro, F.; Rakhshandehroo, F.; Larruy, B.; Aguilar, E.; Tenllado, F.; Canto, T. Effects of simultaneously elevated temperature and CO2 levels on Nicotiana benthamiana and its infection by different positive-sense RNA viruses are cumulative and virus type-specific. Virology 2017, 511, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.N.; Koh, S.W.; Choi, K.S.; Joa, J.H.; Kim, C.H.; Selvakumar, G. Temperature and CO2 Level Influence Potato leafroll virus Infection in Solanum tuberosum. Plant Pathol. J. 2017, 33, 522–527. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, E.; del Toro, F.J.; Figueira-Galán, D.; Hou, W.; Canto, T.; Tenllado, F. Virus infection induces resistance to Pseudomonas syringae and to drought in both compatible and incompatible bacteria—Host interactions, which are compromised under conditions of elevated temperature and CO2 levels. J. Gen. Virol. 2020, 101, 122–135. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meziadi, C.; Lintz, J.; Naderpour, M.; Gautier, C.; Blanchet, S.; Noly, A.; Gratias-Weill, A.; Geffroy, V.; Pflieger, S. R-BPMV-Mediated Resistance to Bean pod mottle virus in Phaseolus vulgaris L. Is Heat-Stable but Elevated Temperatures Boost Viral Infection in Susceptible Genotypes. Viruses 2021, 13, 1239. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071239
Meziadi C, Lintz J, Naderpour M, Gautier C, Blanchet S, Noly A, Gratias-Weill A, Geffroy V, Pflieger S. R-BPMV-Mediated Resistance to Bean pod mottle virus in Phaseolus vulgaris L. Is Heat-Stable but Elevated Temperatures Boost Viral Infection in Susceptible Genotypes. Viruses. 2021; 13(7):1239. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071239
Chicago/Turabian StyleMeziadi, Chouaïb, Julie Lintz, Masoud Naderpour, Charlotte Gautier, Sophie Blanchet, Alicia Noly, Ariane Gratias-Weill, Valérie Geffroy, and Stéphanie Pflieger. 2021. "R-BPMV-Mediated Resistance to Bean pod mottle virus in Phaseolus vulgaris L. Is Heat-Stable but Elevated Temperatures Boost Viral Infection in Susceptible Genotypes" Viruses 13, no. 7: 1239. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071239