
Aerobic Conditions and Endogenous Reactive Oxygen Species Reduce the Production of Infectious MS2 Phage by Escherichia coli

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Production and Purification of Phage Suspension
2.2. Phage Replication Conditions during Plaque Formation
2.3. Extraction of MS2 Phages from Lysis Plaques
2.4. Quantification of Infectious Phages in Individual Lysis Plaques
2.5. Quantification of MS2-RNA in Plaques, Using RT-qPCR
2.6. MS2-RNA Quantification Using Digital Droplet PCR to Establish the Standard Curve for RT-qPCR
2.7. Visualization of E. coli Growth on Semi-Agar Media
2.8. Data Analysis
3. Results
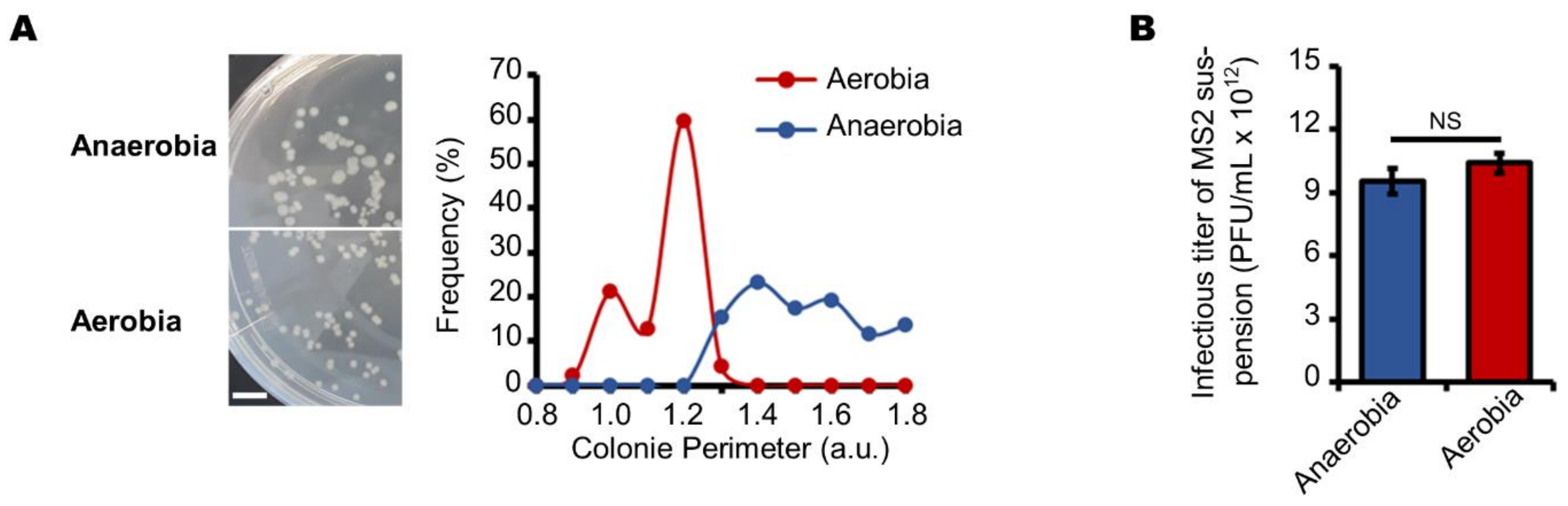
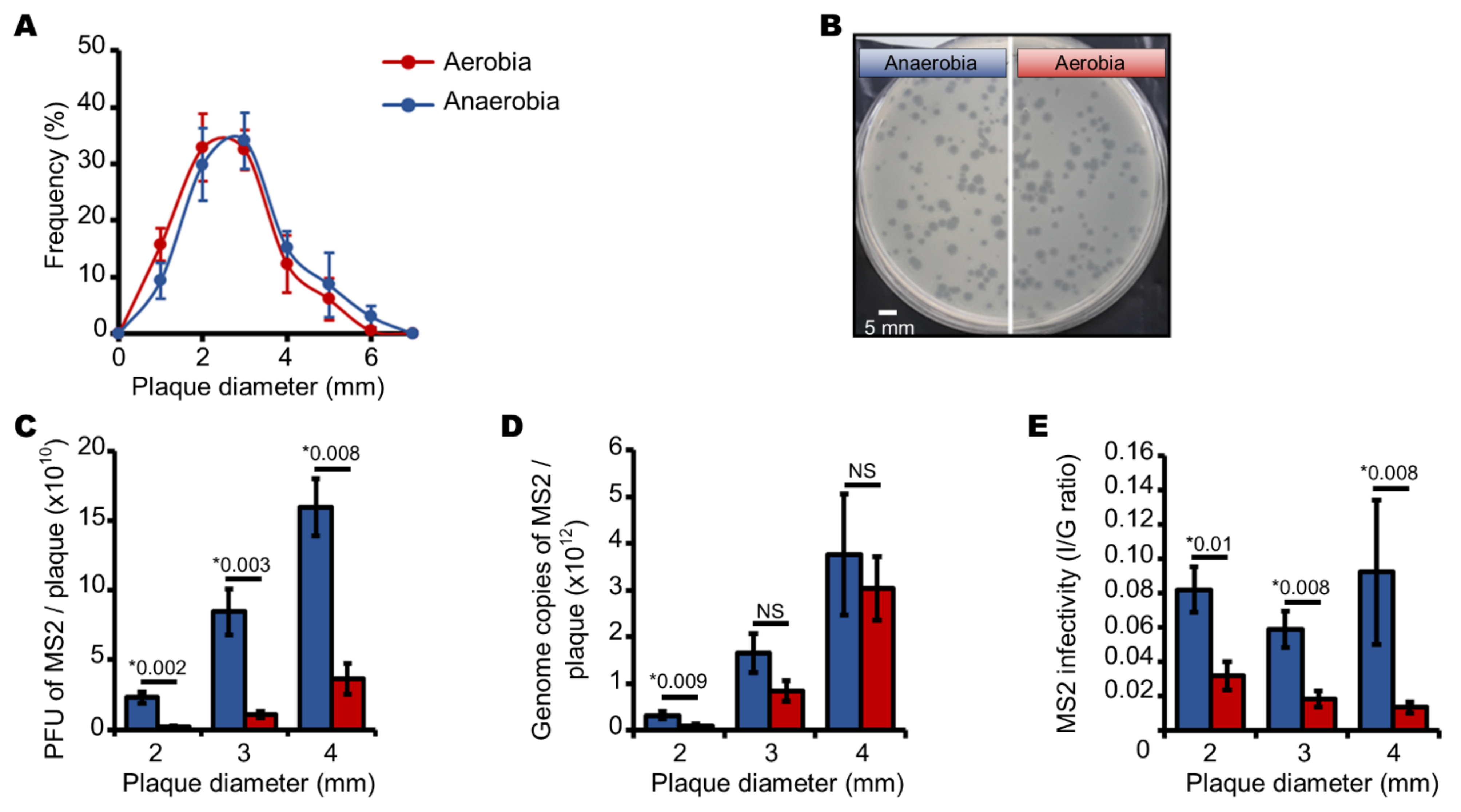
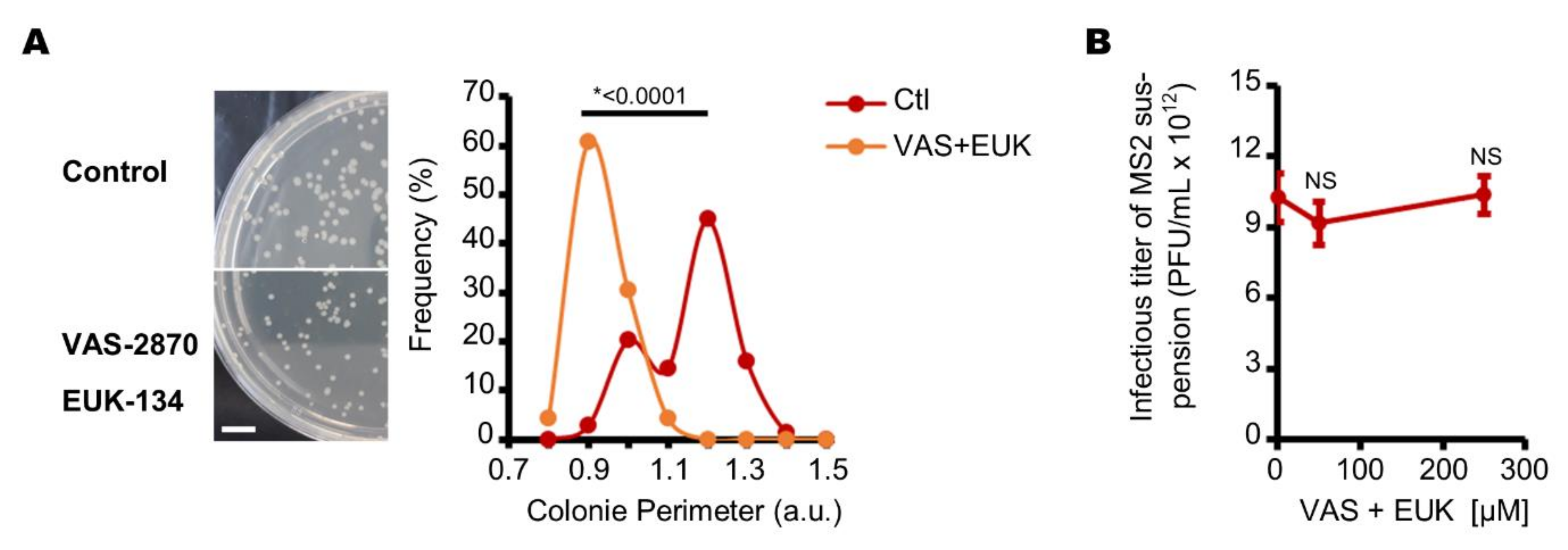
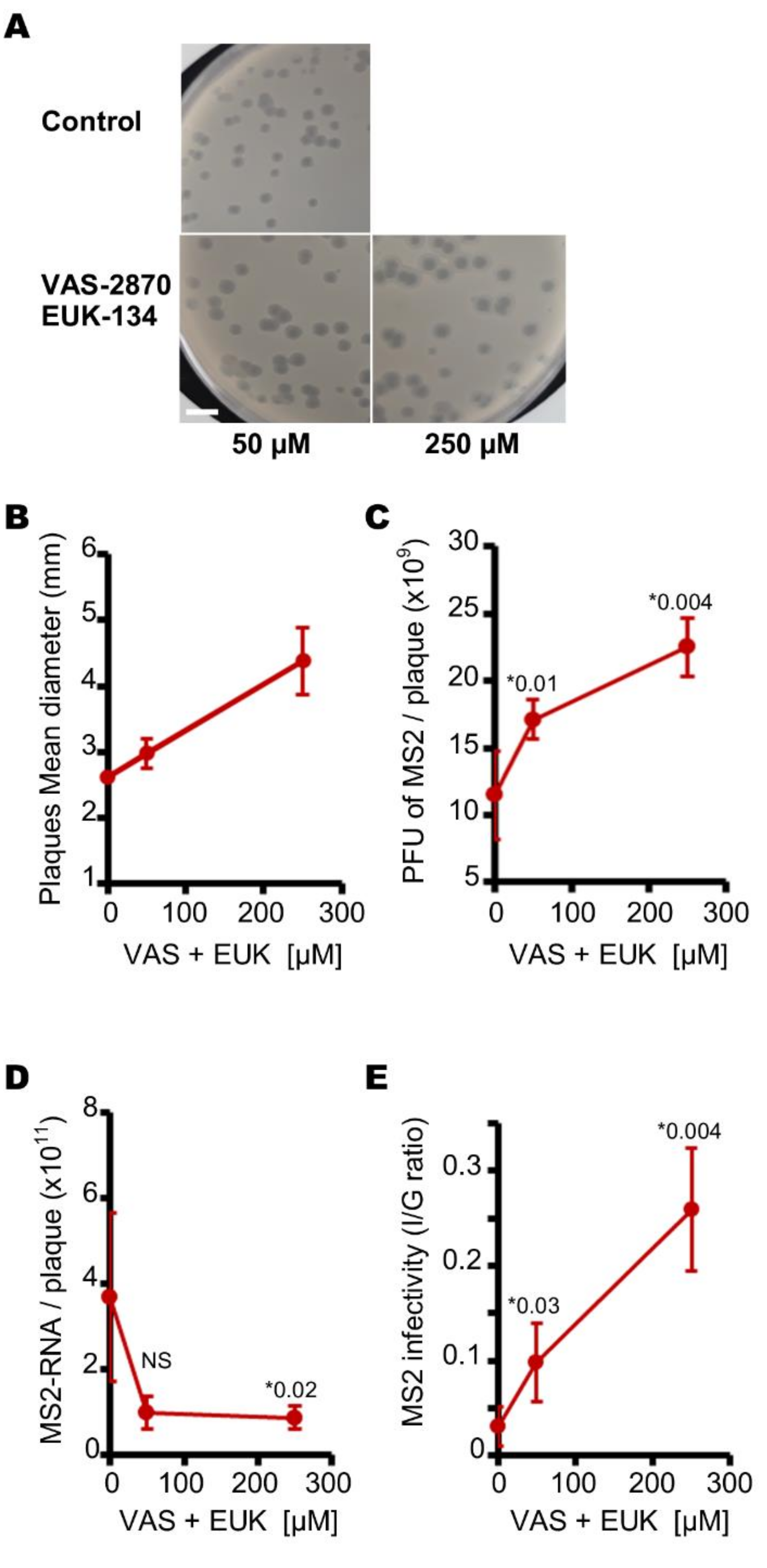
3.1. Aerobic Culture Conditions Decrease the Production of Infectious MS2 Phages
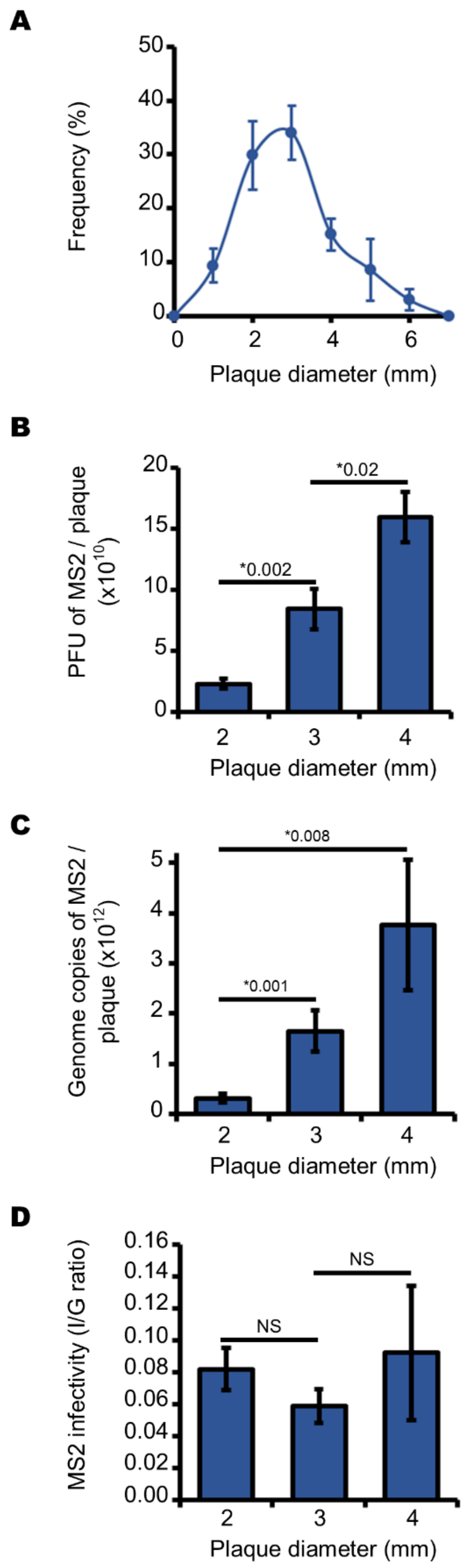
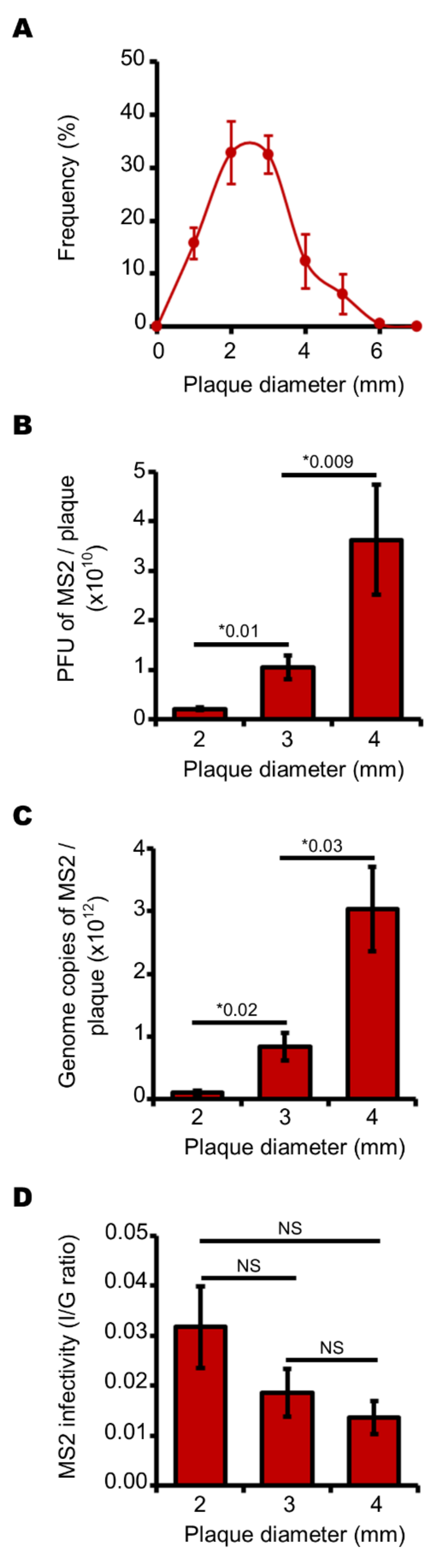
3.2. Endogenous Reactive Oxygen Species Produced by E. coli Decrease the Production of Infectious MS2 Phages
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mikel, P.; Vasickova, P.; Kralik, P. Methods for Preparation of MS2 Phage-Like Particles and Their Utilization as Process Control Viruses in RT-PCR and qRT-PCR Detection of RNA Viruses From Food Matrices and Clinical Specimens. Food Environ. Virol. 2015, 7, 96–111. [Google Scholar] [CrossRef]
- Brié, A.; Gantzer, C.; Boudaud, N.; Bertrand, I. The impact of chlorine and heat on the infectivity and physicochemical properties of bacteriophage MS2. FEMS Microbiol. Ecol. 2018, 94, fiy106. [Google Scholar] [CrossRef]
- Hartard, C.; Banas, S.; Loutreul, J.; Rincé, A.; Benoit, F.; Boudaud, N.; Gantzer, C. Relevance of F-Specific RNA Bacteriophages in Assessing Human Norovirus Risk in Shellfish and Environmental Waters. Appl. Environ. Microbiol. 2016, 82, 5709–5719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, B.V.V.; Hardy, M.E.; Dokland, T.; Bella, J.; Rossmann, M.G.; Estes, M.K. X-ray Crystallographic Structure of the Norwalk Virus Capsid. Science 1999, 286, 287–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, X.; Li, Z.; Lai, M.; Shu, S.; Du, Y.; Zhou, Z.H.; Sun, R. In situ structures of the genome and genome-delivery apparatus in a single-stranded RNA virus. Nature 2017, 541, 112–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fauvel, B.; Cauchie, H.M.; Gantzer, C.; Ogorzaly, L. Influence of physico-chemical characteristics of sediment on the in situ spatial distribution of F-specific RNA phages in the riverbed. FEMS Microbiol. Ecol. 2019, 95, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Elmahdy, M.E.I.; Fongaro, G.; Magri, M.E.; Petruccio, M.M.; Barardi, C.R.M. Spatial distribution of enteric viruses and somatic coliphages in a Lagoon used as drinking water source and recreation in Southern Brazil. Int. J. Hyg. Environ. Health 2016, 219, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Straub, T.M.; Pepper, I.L.; Gerba, C.P. Cell culture and PCR determination of poliovirus inactivation by disinfectants. Appl. Environ. Microbiol. 1994, 60, 4203–4206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobsey, M.D.; Battigelli, D.A.; Shin, G.A.; Newland, S. RT-PCR amplification detects inactivated viruses in water and wastewater. Water Sci. Technol. 1998, 38, 91–94. [Google Scholar] [CrossRef]
- Simonet, J.; Gantzer, C. Inactivation of poliovirus 1 and F-specific RNA phages and degradation of their genomes by UV irradiation at 254 nanometers. Appl. Environ. Microbiol. 2006, 72, 7671–7677. [Google Scholar] [CrossRef] [Green Version]
- Sigstam, T.; Gannon, G.; Cascella, M.; Pecson, B.M.; Wigginton, K.R.; Kohn, T. Subtle differences in virus composition affect disinfection kinetics and mechanisms. Appl. Environ. Microbiol. 2013, 79, 3455–3467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wigginton, K.R.; Menin, L.; Sigstam, T.; Gannon, G.; Cascella, M.; Hamidane, H.B.; Tsybin, Y.O.; Waridel, P.; Kohn, T. UV Radiation Induces Genome-Mediated, Site-Specific Cleavage in Viral Proteins. ChemBioChem 2012, 13, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, M.; Tohya, Y.; Matsubara, K.; Haramoto, E.; Utagawa, E.; Katayama, H. Chlorine inactivation of human norovirus, murine norovirus and poliovirus in drinking water. Lett. Appl. Microbiol. 2010, 51, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Shin, G.A.; Sobsey, M.D. Inactivation of norovirus by chlorine disinfection of water. Water Res. 2008, 42, 4562–4568. [Google Scholar] [CrossRef] [PubMed]
- Tung, G.; Macinga, D.; Arbogast, J. Efficacy of Commonly Used Disinfectants for Inactivation of Human Noroviruses and Their Surrogates. J. Food Prot. 2013, 76, 1210–1217. [Google Scholar] [CrossRef]
- Wigginton, K.R.; Kohn, T. Virus disinfection mechanisms: The role of virus composition, structure, and function. Curr. Opin. Virol. 2012, 2, 84–89. [Google Scholar] [CrossRef]
- Friedman, E.S.; Bittinger, K.; Esipova, T.V.; Hou, L.; Chau, L.; Jiang, J.; Mesaros, C.; Lund, P.J.; Liang, X.; Fitzgerald, G.A.; et al. Microbes vs. chemistry in the origin of the anaerobic gut lumen. Proc. Natl. Acad. Sci. USA 2018, 115, 4170–4175. [Google Scholar] [CrossRef] [Green Version]
- Bundy, B.C.; Swartz, J.R. Efficient disulfide bond formation in virus-like particles. J. Biotechnol. 2011, 154, 230–239. [Google Scholar] [CrossRef]
- Buccigrossi, V.; Laudiero, G.; Russo, C.; Miele, E.; Sofia, M.; Monini, M.; Ruggeri, F.M.; Guarino, A. Chloride secretion induced by rotavirus is oxidative stress-dependent and inhibited by Saccharomyces boulardii in human enterocytes. PLoS ONE 2014, 9, e99830. [Google Scholar] [CrossRef] [Green Version]
- Bai, Z.; Zhao, X.; Li, C.; Sheng, C.; Li, H. EV71 virus reduces Nrf2 activation to promote production of reactive oxygen species in infected cells. Gut Pathog. 2020, 12, 1–12. [Google Scholar] [CrossRef]
- Reshi, M.L.; Su, Y.; Hong, J. RNA Viruses: ROS-Mediated Cell Death. Int. J. Cell Biol. 2014, 2014, 467452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, T.G.; Dong, S.; Catalano, C.; Moore, R.; Liang, X.; Mekalanos, J.J. Generation of reactive oxygen species by lethal attacks from competing microbes. Proc. Natl. Acad. Sci. USA 2015, 112, 2181–2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, A.V.; Valuev-elliston, V.T.; Ivanova, O.N.; Kochetkov, S.N.; Starodubova, E.S.; Bartosch, B.; Isaguliants, M.G. Oxidative Stress during HIV Infection: Mechanisms and Consequences. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Casteel, M.J.; Schmidt, C.E.; Sobsey, M.D. Chlorine disinfection of produce to inactivate hepatitis A virus and coliphage MS2. Int. J. Food Microbiol. 2008, 125, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro, R.; Misawa, T.; Sakudo, A. Key role of singlet oxygen and peroxynitrite in viral RNA damage during virucidal effect of plasma torch on feline calicivirus. Sci. Rep. 2018, 8, 17947. [Google Scholar] [CrossRef] [PubMed]
- Simonet, J.; Gantzer, C. Degradation of the Poliovirus 1 genome by chlorine dioxide. J. Appl. Microbiol. 2006, 100, 862–870. [Google Scholar] [CrossRef]
- Wigginton, K.; Pecson, B.M.; Sigstam, T.; Bosshard, F.; Kohn, T. Virus inactivation mechanisms: Impact of Disinfectants on Virus Function and Structural Integrity. Environ. Sci. Technol. 2012, 46, 12069–12078. [Google Scholar] [CrossRef]
- Young, M.; Kim, J.; Ko, G. Disinfection kinetics of murine norovirus using chlorine and chlorine dioxide. Water Res. 2010, 44, 3243–3251. [Google Scholar] [CrossRef]
- Cromeans, T.L.; Kahler, A.M.; Hill, V.R. Inactivation of Adenoviruses, Enteroviruses, and Murine Norovirus in Water by Free Chlorine and Monochloramine. Appl. Environ. Microbiol. 2010, 76, 1028–1033. [Google Scholar] [CrossRef] [Green Version]
- Padalko, E.; Ohnishi, T.; Matsushita, K.; Sun, H.; Fox-talbot, K.; Bao, C.; Baldwin, W.M.; Lowenstein, C.J. Peroxynitrite inhibition of Coxsackievirus infection by prevention of viral RNA entry. Proc. Natl. Acad. Sci. USA 2004, 101, 11731–11736. [Google Scholar] [CrossRef] [Green Version]
- Sano, D.; Pintó, R.M.; Omura, T.; Bosch, A. Detection of oxidative damages on viral capsid protein for evaluating structural integrity and infectivity of human norovirus. Environ. Sci. Technol. 2010, 44, 808–812. [Google Scholar] [CrossRef]
- Araud, E.; Shisler, J.L.; Nguyen, T.H. Inactivation Mechanisms of Human and Animal Rotaviruses by Solar UVA and Visible Light. Environ. Sci. Technol. 2018, 52, 5682–5690. [Google Scholar] [CrossRef]
- Bastin, G.; Loison, P.; Vernex-Loset, L.; Dupire, F.; Challant, J.; Majou, D.; Boudaud, N.; Krier, G.; Gantzer, C. Structural Organizations of Qβ and MS2 Phages Affect Capsid Protein Modifications by Oxidants Hypochlorous Acid and Peroxynitrite. Front. Microbiol. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Richardson, S.D.; Thruston, A.D., Jr.; Caughran, T.V.; Chen, P.H.; Collette, T.W.; Schenck, K.M.; Lykins, B.W., Jr.; Rav-Acha, C.; Glezer, V. Identification of new drinking water disinfection by-products from ozone, chlorine dioxide, chloramine, and chlorine. Environ. Prot. 2000, 123, 95–102. [Google Scholar] [CrossRef]
- Page, M.A.; Shisler, J.L.; Marin, B.J. Mechanistic Aspects of Adenovirus Serotype 2 Inactivation with Free Chlorine. Appl. Environ. Microbiol. 2010, 76, 2946–2954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogata, N. Inactivation of influenza virus haemagglutinin by chlorine dioxide: Oxidation of the conserved tryptophan 153 residue in the receptor-binding site. J. Gen. Virol. 2012, 93, 2558–2563. [Google Scholar] [CrossRef] [Green Version]
- Ward, R.L.; Knowlton, D.R.; Pierce, M.J. Efficiency of human rotavirus propagation in cell culture. J. Clin. Microbiol. 1984, 19, 748–753. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, R.A.; Pepper, I.L.; Gerba, C.P. Application of PCR-Based Method to Assess the Infectivity of Enteric Viruses in Environmental Samples. Appl Env. Microbiol 2009, 75, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Jiang, S.C. Real-time PCR quantification of human adenoviruses in urban rivers indicates genome prevalence but low infectivity. Appl. Environ. Microbiol. 2005, 71, 7426–7433. [Google Scholar] [CrossRef] [Green Version]
- Loison, P.; Majou, D.; Gelhaye, E.; Boudaud, N.; Gantzer, C. Impact of reducing and oxidizing agents on the infectivity of Qβ phage and the overall structure of its capsid. FEMS Microbiol. Ecol. 2016, 92, 1–11. [Google Scholar] [CrossRef] [Green Version]
- International Organization for Standardization. ISO 10705-1. Water Quality: Detection and Enumeration of Bacteriophages. Part 1: Enumeration of F-Specific RNA Bacteriophages; International Organization for Standardization: Geneva, Switzerland, 2001. [Google Scholar]
- Dao, V.T.V.; Elbatreek, M.H.; Altenhöfer, S.; Casas, A.I.; Pachado, M.P.; Neullens, C.T.; Knaus, U.G.; Schmidt, H.H.H.W. Isoform-selective NADPH oxidase inhibitor panel for pharmacological target validation. Free Radic. Biol. Med. 2020, 148, 60–69. [Google Scholar] [CrossRef]
- Reis, J.; Massari, M.; Marchese, S.; Ceccon, M.; Aalbers, F.S.; Corana, F.; Valente, S.; Mai, A.; Magnani, F.; Mattevi, A. A closer look into NADPH oxidase inhibitors: Validation and insight into their mechanism of action. Redox Biol. 2020, 32, 101466. [Google Scholar] [CrossRef]
- Brain, I.; Baker, K.; Marcus, C.B.; Huffman, K.; Kruk, H.; Malfroy, B.; Doctrow, S.R. Synthetic Combined Superoxide Dismutase/ Catalase Mimetics Are Protective as a Delayed Treatment in a Rat Stroke Model: A Key Role for Reactive Oxygen Species in Ischemic Brain Injury. J. Pharmacol. Exp. Ther. 1998, 284, 215–221. [Google Scholar]
- Munroe, W.; Kingsley, C.; Durazo, A.; Butler Gralla, E.; Imlay, J.A.; Srinivasan, C.; Selverstone Valentine, J. Only one of a wide assortment of manganese-containing SOD mimicking compounds rescues the slow aerobic growth phenotypes of both Escherichia coli and Saccharomyces cerevisiae strains lacking superoxide dismutase enzymes. J. Inorg. Biochem. 2007, 101, 1875–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogorzaly, L.; Gantzer, C. Erratum to “Development of real-time RT-PCR methods for specific detection of F-specific RNA bacteriophage genogroups: Application to urban raw wastewater”. J. Virol. Methods 2007, 143, 122. [Google Scholar] [CrossRef]
- Mendoza-Chamizo, B.; Løbner-Olesen, A.; Charbon, G. Coping with reactive oxygen species to ensure genome stability in escherichia coli. Genes 2018, 9, 565. [Google Scholar] [CrossRef] [Green Version]
- Charbon, G.; Bjørn, L.; Mendoza-Chamizo, B.; Frimodt-Møller, J.; Løbner-Olesen, A. Oxidative DNA damage is instrumental in hyperreplication stress-induced inviability of Escherichia coli. Nucleic Acids Res. 2014, 42, 13228–13241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamison, C.S.; Adler, H.I. Mutations in Escherichia coli that effect sensitivity to oxygen. J. Bacteriol. 1987, 169, 5087–5094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Na, Y.A.; Lee, J.Y.; Bang, W.J.; Lee, H.J.; Choi, S.I.; Kwon, S.K.; Jung, K.H.; Kim, J.F.; Kim, P. Growth retardation of Escherichia coli by artificial increase of intracellular ATP. J. Ind. Microbiol. Biotechnol. 2015, 42, 915–924. [Google Scholar] [CrossRef]
- Murashko, O.N.; Lin-Chao, S. Escherichia coli responds to environmental changes using enolasic degradosomes and stabilized DicF sRNA to alter cellular morphology. Proc. Natl. Acad. Sci. USA 2017, 114, E8025–E8034. [Google Scholar] [CrossRef] [Green Version]
- Magnani, F.; Mattevi, A. Structure and mechanisms of ROS generation by NADPH oxidases. Curr. Opin. Struct. Biol. 2019, 59, 91–97. [Google Scholar] [CrossRef]
- Hajjar, C.; Cherrier, M.V.; Mirandela, G.D.; Petit-Hartlein, I.; Stasia, M.J.; Fontecilla-Camps, J.C.; Fieschi, F.; Dupuy, J. The NOX family of proteins is also present in bacteria. MBio 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Imlay, J.A. The fumarate reductase of Bacteroides thetaiotaomicron, unlike that of Escherichia coli, is configured so that it does not generate reactive oxygen species. MBio 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meehan, B.M.; Malamy, M.H. Fumarate reductase is a major contributor to the generation of reactive oxygen species in the anaerobe Bacteroides fragilis. Microbiology 2012, 158, 539–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asad, N.R.; Buarque Oliveira Asad, L.M.; Bonacossa De Almeida, C.E.; Felzenszwalb, I.; Bispo Cabral-Neto, J.; Leitão, A.C. Several pathways of hydrogen peroxide action that damage the E. coli genome. Genet. Mol. Biol. 2004, 27, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Gallet, R.; Kannoly, S.; Wang, I. Effects of bacteriophage traits on plaque formation. BMC Microbiol. 2011, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Biebricher, C.K.; Duker, E.M. F and type 1 piliation of Escherichia coli. J. Gen. Microbiol. 1984, 130, 951–957. [Google Scholar] [CrossRef] [Green Version]
- Roychoudhury, P.; Shrestha, N.; Wiss, V.R.; Krone, S.M. Fitness benefits of low infectivity in a spatially structured population of bacteriophages. Proc. R. Soc. B 2013, 281, 20132563. [Google Scholar] [CrossRef] [Green Version]
- Sobota, J.M.; Imlay, J.A. Iron enzyme ribulose-5-phosphate 3-epimerase in Escherichia coli is rapidly damaged by hydrogen peroxide but can be protected by manganese. Proc. Natl. Acad. Sci. USA 2011, 108, 5402–5407. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.F.; Mashino, T.; Fridovich, I. α,β-Dihydroxyisovalerate dehydratase. A superoxide-sensitive enzyme. J. Biol. Chem. 1987, 262, 4724–4727. [Google Scholar] [CrossRef]
- Hyodo, K.; Hashimoto, K.; Kuchitsu, K.; Suzuki, N.; Okuno, T. Harnessing host ROS-generating machinery for the robust genome replication of a plant RNA virus. Proc. Natl. Acad. Sci. USA 2017, 114, E1282–E1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camini, F.C.; da Silva Caetano, C.C.; Almeida, L.T.; de Brito Magalhães, C.L. Implications of oxidative stress on viral pathogenesis. Arch. Virol. 2017, 162, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, R.; Ploen, D.; Hildt, E. HCV and Oxidative Stress: Implications for HCV Life Cycle and HCV-Associated Pathogenesis. Oxid. Med. Cell. Longev. 2016, 2016, 9012580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C. Therapeutic Modulation of Virus-Induced Oxidative Stress via the Nrf2-Dependent Antioxidative Pathway. Oxid. Med. Cell. Longev. 2018, 2018, 6208067. [Google Scholar] [CrossRef] [PubMed]
- Chamakura, R.K.; Tran S, J.; Young, R. MS2 Lysis of Escherichia coli Depends on Host Chaperone DnaJ. J. Bacteriol. 2017, 199, e00058-17. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bastin, G.; Galmiche, A.; Talfournier, F.; Mazon, H.; Challant, J.; Robin, M.; Majou, D.; Boudaud, N.; Gantzer, C. Aerobic Conditions and Endogenous Reactive Oxygen Species Reduce the Production of Infectious MS2 Phage by Escherichia coli. Viruses 2021, 13, 1376. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071376
Bastin G, Galmiche A, Talfournier F, Mazon H, Challant J, Robin M, Majou D, Boudaud N, Gantzer C. Aerobic Conditions and Endogenous Reactive Oxygen Species Reduce the Production of Infectious MS2 Phage by Escherichia coli. Viruses. 2021; 13(7):1376. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071376
Chicago/Turabian StyleBastin, Guillaume, Aurélie Galmiche, François Talfournier, Hortense Mazon, Julie Challant, Maëlle Robin, Didier Majou, Nicolas Boudaud, and Christophe Gantzer. 2021. "Aerobic Conditions and Endogenous Reactive Oxygen Species Reduce the Production of Infectious MS2 Phage by Escherichia coli" Viruses 13, no. 7: 1376. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071376