
Fungal F8-Culture Filtrate Induces Tomato Resistance against Tomato Yellow Leaf Curl Thailand Virus




1
Agricultural Biotechnology Research Center, Academia Sinica, Taipei 11529, Taiwan
2
Department of Entomology, National Taiwan University, Taipei 10617, Taiwan
3
Department of Plant Pathology and Microbiology, National Taiwan University, Taipei 10617, Taiwan
4
Institute of Biotechnology, National Taiwan University, Taipei 10617, Taiwan
*
Author to whom correspondence should be addressed.
Viruses 2021, 13(8), 1434; https://0-doi-org.brum.beds.ac.uk/10.3390/v13081434
Submission received: 11 June 2021
/
Revised: 18 July 2021
/
Accepted: 20 July 2021
/
Published: 23 July 2021
(This article belongs to the Special Issue State-of-the-Art Plant-Virus Interactions in Asia)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Tomato (Solanum lycopersicum) is an important economic crop worldwide. However, tomato production is jeopardized by the devastating tomato yellow leaf curl disease caused by whitefly-transmitted begomoviruses (WTBs). In this study, we evaluated the efficacy of our previously developed plant antiviral immunity inducer, fungal F8-culture filtrate, on tomato to combat tomato yellow leaf curl Thailand virus (TYLCTHV), the predominant WTB in Taiwan. Our results indicated that F8-culture filtrate treatment induced strong resistance, did not reduce the growth of tomato, and induced prominent resistance against TYLCTHV both in the greenhouse and in the field. Among TYLCTHV-inoculated Yu-Nu tomato grown in the greenhouse, a greater percentage of plants treated with F8-culture filtrate (43–100%) were healthy-looking compared to the H2O control (0–14%). We found that TYLCTHV cannot move systemically only on the F8-culture filtrate pretreated healthy-looking plants. Tracking the expression of phytohormone-mediated immune maker genes revealed that F8-culture filtrate mainly induced salicylic acid-mediated plant immunity. Furthermore, callose depositions and the expression of the pathogen-induced callose synthase gene, POWDERY MILDEW RESISTANT 4 were only strongly induced by TYLCTHV on tomato pretreated with F8-culture filtrate. This study provides an effective way to induce tomato resistance against TYLCTHV.
1. Introduction
Tomato (Solanum lycopersicum) is an economically important vegetable crop, which ranks third in this class in terms of volume of production and first in terms of processing volume [1]. However, tomato yellow leaf curl disease (TYLCD) caused by whitefly-transmitted begomoviruses (WTBs) has a devastating effect on tomato production worldwide [2,3,4,5]. One of the main approaches to managing TYLCD relies on controlling the whitefly vector using insecticide [6,7]. However, it has become increasingly difficult to control the whitefly vector population in this way because of the rapid evolution of whitefly’s resistance to insecticides [2,7,8].
Screening of TYLCD-resistant tomato cultivars revealed that most commercially cultivated tomato cultivars are susceptible to the disease [9]. To breed TYLCD-resistant cultivars, six TYLCD resistance gene loci (Ty gene family) were identified from wild tomato species [6,10,11,12]. However, the breeding process requires several backcross steps and is usually time consuming. As a large number of tomato cultivars are planted for commercial purposes, transferring the resistance loci to these various commercial tomato cultivars remains a challenge. Transgenic approaches that mainly use RNAi strategies have been successfully applied to plants to manage TYLCD infection with promising results [13,14,15,16,17,18,19,20,21,22,23,24]. However, the debate over using genetically modified organisms (GMO) in agriculture has hindered the distribution of these transgenic tomatoes. One way to address this issue would be to induce plant innate immunity to combat TYLCD.
Plant innate immunity consists of two distinctive pathways: microbe/pathogen-associated molecular pattern (MAMP/PAMP)-triggered immunity (PTI) and effector-triggered immunity (ETI) [25,26,27,28,29]. PTI is triggered by MAMPs/PAMPs via cell surface-localized pattern-recognition receptors (PRRs), whereas ETI is activated by pathogen effector proteins predominantly through direct or indirect perception of intracellular nucleotide-binding, leucine-rich repeat receptors (NLRs) [25,30,31,32]. At the first layer of plant innate immunity, PTI, signals are transmitted through activation of mitogen-activated protein kinases (MAPKs), inducing signaling and defense processes such as reactive oxygen species (ROS) production, ion fluxes, salicylic acid (SA) and ethylene (ET) accumulation, resistance gene activation such as genes encoding pathogenesis-related proteins (PRs), and callose deposition [33,34,35,36,37,38]. However, adapted pathogens can deliver diverse effectors into plant cells to interfere with PTI and enhance virulence [39,40,41,42,43]. To counteract the invasion, plants utilize intracellular polymorphic resistance (R) proteins to perceive these effectors either through direct binding or monitoring perturbations of host proteins caused by effectors, to initiate a more robust defense ETI [29,44]. ETI often leads to hypersensitive response (HR), a type of programmed cell death that has been hypothesized to restrict pathogen growth at the local infection site [32,45,46,47].
The application of ETI in plant protection requires introduction of R protein into susceptible cultivars by breeding or transgenic approaches, which is time consuming, as mentioned above. In addition, WTBs evolve quickly which leads to the occurrence of resistance breaking strains as R protein usually recognizes specific effectors. In comparison to ETI, PTI can be induced in all cultivars, and has the potential to be applied to all tomato cultivars. Indeed, several plant immunity inducers from biological sources have been reported including proteins [48,49,50,51,52,53], oligosaccharides [54,55], microbial inducers [56,57,58], and oil and related compounds [59]. However, the application of plant immunity inducers on crop protection requires the need to overcome several challenges, such as the high cost of the inducers, effectiveness in plant protection, and reduced crop growth and quality [60,61]. Therefore, practical application of plant immunity inducers on crops for effective protection mostly remains to be explored.
In our previous study, we screened culture filtrates derived from soil microorganisms cultured with medium made of weeds and cheaply obtainable vegetables (may contain pathogen/damage-associated molecular patterns) for induction of effective plant defense against viruses [62]. We obtained a culture filtrate derived from Trichosporon sp. (F8-culture filtrate) that induced strong resistance to different viruses in various plants [62]. The infection rate of tobacco mosaic virus (TMV) reduced dramatically on Nicotiana benthamiana pretreated with F8-culture filtrate [62]. The symptoms were also considerably reduced on F8-culture filtrate-pretreated N. benthamiana and Brassica juncea inoculated with cucumber mosaic virus (CMV) and turnip mosaic virus (TuMV), respectively. A fraction of the polysaccharides, designated F8-polysaccharide, with estimated molecular weight of 4260–12,000 kDa is responsible for induction of the plant antiviral immunity. The F8-polysaccharide mainly induced priming of the SA immune pathway, and therefore, has great potential for application on tomato against viruses as previous reports have also indicated that SA-mediated plant immunity is important in tomato resistance to viral diseases including TYLCD [63].
In this study, we applied F8-culture filtrate to tomato to combat tomato yellow leaf curl Thailand virus (TYLCTHV), the predominant WTB in Taiwan [64,65,66]. Our results indicated that F8-culture filtrate treatment in tomatoes induced strong resistance to TYLCTHV both in the greenhouse and in the field. Furthermore, we resolved the main immune responses in F8-culture filtrate-treated tomato. Our results not only provide an effective method to manage TYLCD, but also provide important knowledge for further design strategies to combat the effect of WTBs on crops.
2. Materials and Methods
2.1. Whiteflies, Virus and Plants
The sources of our whitefly (Bemisia tabaci Middle East–Asia Minor 1, MEAM1) and TYLCTHV isolate Ly5 (GenBank Accession No. GU723742 and GU723754) are described in Weng et al. [67].
The non-viruliferous whitefly was reared on Chinese kale (Brassica oleracea cv. Alboglabra Group) in whitefly-proof net cages (30 × 30 × 30 cm3) at 26 °C, under a 16-h light/8-h dark cycle.
TYLCTHV isolate Ly5 was maintained, as previously described [68], in tomato plants (Solanum lycopersicum cv. ANT22) and used for vector transmission, using B. tabaci, under the aforementioned conditions.
Tomato cultivar Yu-Nu (S. lycopersicum) was purchased from Known-You Seed (Kaohsiung, Taiwan), and grown and maintained in a glass-covered greenhouse. Xiao-Nu (S. lycopersicum) was purchased from Known-You Seed and maintained in the field (Alian District, Kaohsiung City, Taiwan) for field analysis.
2.2. Treatment and Inoculation
Yu-Nu tomatoes (2–3 weeks after sowing) were sprayed with H2O (0.4 mL per plant) or F8-culture filtrate (0.4 mL per plant) for a total of three times with a 24-h interval between each spray as previously described [62]. Twenty-four hours after the final treatment, plants were inoculated with TYLCTHV by viruliferous B. tabaci as previously described [69] with modifications. Briefly, non-viruliferous B. tabaci was transferred to the net cage enclosed with TYLCTHV-infected ANT22 tomato and allowed 3-day acquisition access periods (AAP). The viruliferous B. tabaci was used to inoculate tomato plants through feeding for 96 h (approximately 20 whiteflies on one leaf per plant). The inoculated-leaves were cut and collected at 4 dpi. The number of replicated experimental sets and the number of biological replicates for each assay are specified in the Results section.
2.3. Disease Index for Evaluation of Tomato Yellow Leaf Curl Disease (TYLCD)
Tomatoes were evaluated for TYLCD severity according to a disease index of 5 levels. Level 0 = no visible symptoms, inoculated plants show the same growth and development as non-inoculated plants; Level 1 = slight yellowing and minor curling of leaflet ends; Llevel 2 = wide range of leaf yellowing, curling and cupping, with some reduction in size; Level 3 = wide range of leaf yellowing, pronounced leaf cupping and curling; Level 4 = very severe plant stunting and leaf yellowing, pronounced leaf cupping and curling, and plant ceased to grow.
2.4. Nucleic Acid Isolation and Gene Expression Analysis
Total DNA and RNA were extracted using an optimized CTAB method [70]. Leaf tissue (100 mg) was frozen with liquid nitrogen and ground into a fine powder with a pestle and mortar. The powder was then dissolved in 750 μL of CTAB buffer (2% CTAB, 2% PVP, 2.0 M NaCl, 0.1 M Tris–HCl, 25 mM EDTA, 4% β-mercaptoethanol) and incubated at 65 °C for 30 min. Next, 750 μL of chloroform:IAA (24:1) was added to the mixture and centrifuged at 16,000× g for 10 min. To precipitate DNA and RNA, the upper phase was transferred to a new tube, mixed with 2 volumes of ethanol (99.9% v/v) and 1/4 volume of 10 M LiCl, and centrifuged at 16,000× g for 10 min. The DNA and RNA supernatant were discarded and the pellet was washed with 1 mL of ethanol (70% v/v). After centrifugation, the supernatant was discarded and the pellet was dissolved in 200 μL nuclease-free water (Invitrogen, Carlsbad, CA, USA).
Residual DNA was removed by using the TURBO DNA-free kit (Ambion, Austin, TX, USA) before RT-PCR. cDNA was synthesized by using the PrimeScript RT-PCR Kit (Takara Bio, Shiga, Japan). Quantitative PCR (q-PCR) and quantitative reverse transcription PCR (qRT-PCR) were conducted with the SYBR protocol (Life Technologies, Carlsbad, CA, USA) and the ABI 7100 real-time PCR system (Applied Biosystems, Carlsbad, CA, USA). For quantitative analysis, the common tomato endogenous control, β-actin [71,72,73], was verified for expression stability among different samples and the primer efficiency compared to specified target gene(s) was conducted prior to use. Relative fold change in DNA expression was determined by calculating 2−ΔΔCt. The sequences of primers used for analysis are listed in Table S1.
2.5. Measurement of Tomato Plant Fresh Weight
Tomato Yu-Nu (3 weeks after sowing) were pretreated with H2O or F8-culture filtrate as mentioned above. All shoot tissues of individual plants were measured for fresh weight.
2.6. Staining of Callose Depositions
The tomato plants were pretreated with either H2O or F8-culture filtrate. Leaf samples were collected 24 h post treatment. One leaf per plant (n = 7) with similar growth stage were collected for each treatment group. In addition, for inoculation, viruliferous B. tabaci was used to inoculate on 1 leaf per tomato plant. One inoculated leaf per plant (n = 7) was also collected at 4 days post inoculation for staining of callose deposits. The collected leaves were fixated and de-stained with 1:3 of acetic acid (100%)/ethanol (100%) overnight. After washing in 150 mM K2HPO4 for 30 min, leaves were incubated for 2 h in staining solution (150 mM K2HPO4 and 0.01% aniline blue). Zeiss Imager Z1 fluorescence microscope (Carl Zeiss AG, Oberkochen, Germany) using a DAPI filter was used for imaging. The microscopic field photos were printed and the number of callose depositions were counted manually.
2.7. Field Trial
The field trials were conducted in Yuchi Township, Nantou County, Taiwan, and Alian District, Kaohsiung City, Taiwan. Three-week-old (after sowing) Yu-Nu and Xiao-Nu tomato cultivar seedlings were sprayed with H2O or F8-culture filtrate 3 times with a 24 h interval between each spray. Twenty-four hours after the final treatment, plants were transferred to an open field located at the specified location. The tomato plants were evaluated for disease severity at 2 months after transplantation. Evaluation of disease severity was conducted manually and recorded for each tomato plant based on 5 levels in disease index (see 2.3 Disease Index for Evaluation of Tomato Yellow Leaf Curl Disease and Figure S1).
3. Results
3.1. F8-Culture Filtrate Induced Virus Resistance in Tomato Cultivar Yu-Nu
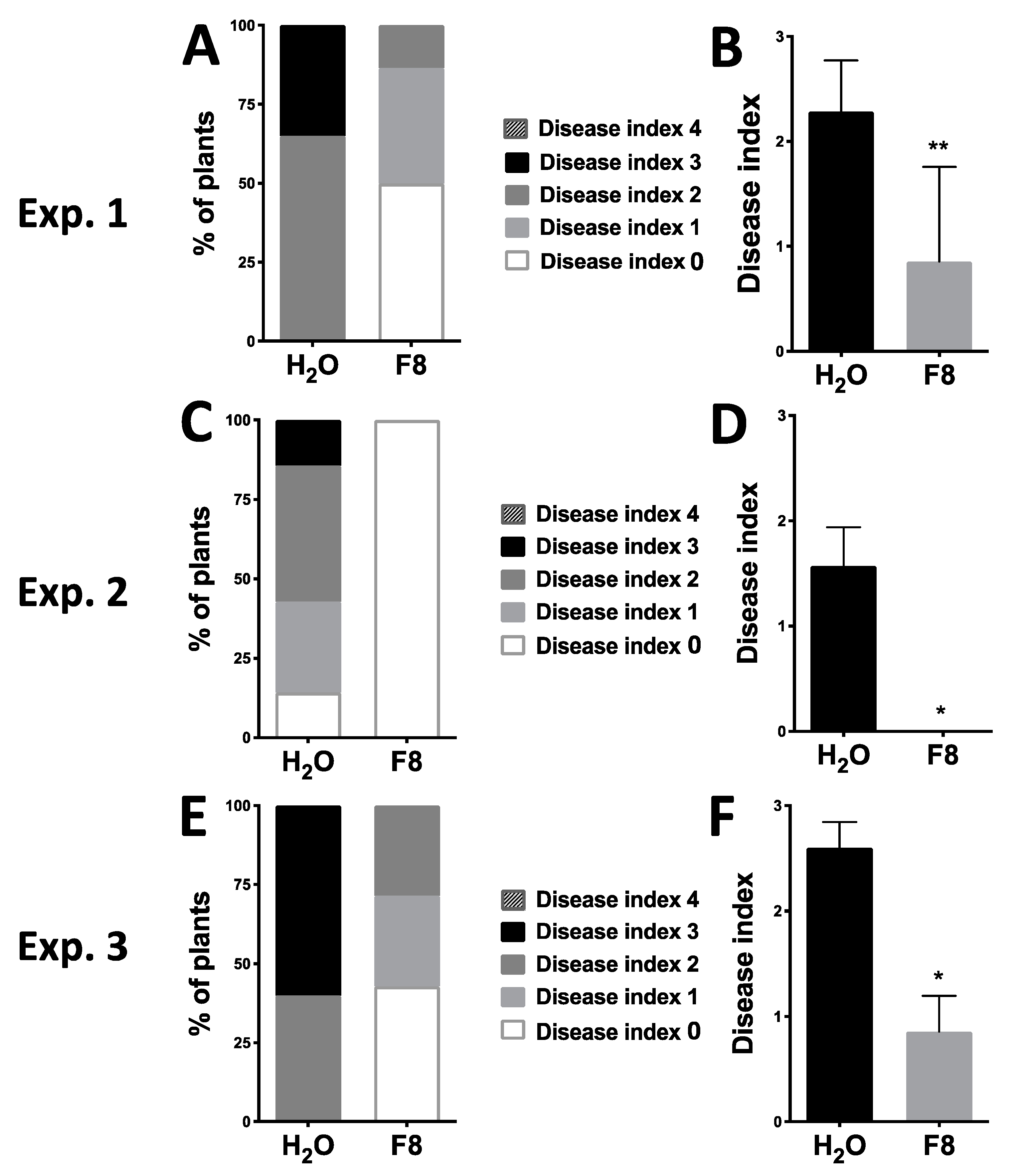
To analyze whether F8-culture filtrate can induce tomato resistance against WTBs, we selected the cherry tomato cultivar, Yu-Nu, for analysis. Yu-Nu tomato is a favorable tomato with commercial value; however, it is known for its susceptibility to several pathogens including WTBs. Initially, we conducted our experiments in a greenhouse. We used the whitefly vector, Bemisia tabaci, as a vector to transmit the TYLCTHV from infected tomato to Yu-Nu. Yu-Nu tomatoes were pretreated with H2O or F8-culture filtrate by spraying consecutively three times with a 24 h interval between each spray. All pretreated plants were inoculated with TYLCTHV by using whitefly vector at 24 h post treatment (hpt). The disease severity was scored using an established disease index (Figure S1) on all TYLCTHV-inoculated plants at 28 days post inoculation (dpi). In our 3 repeated sets of experiments (Exp. 1, 2, and 3), we analyzed 6–7 plants per treatment group (H2O or F8-culture filtrate pretreatment) in each of the experimental sets (Figure 1). Most H2O pretreated plants showed more severe symptoms than the F8-culture filtrate pretreated group (Figure 1). Healthy-looking (Disease Index 0) plants were observed more in the F8-culture filtrate pretreated group compared to the H2O pretreated plants (Figure 1A,C,E and Figure S2).
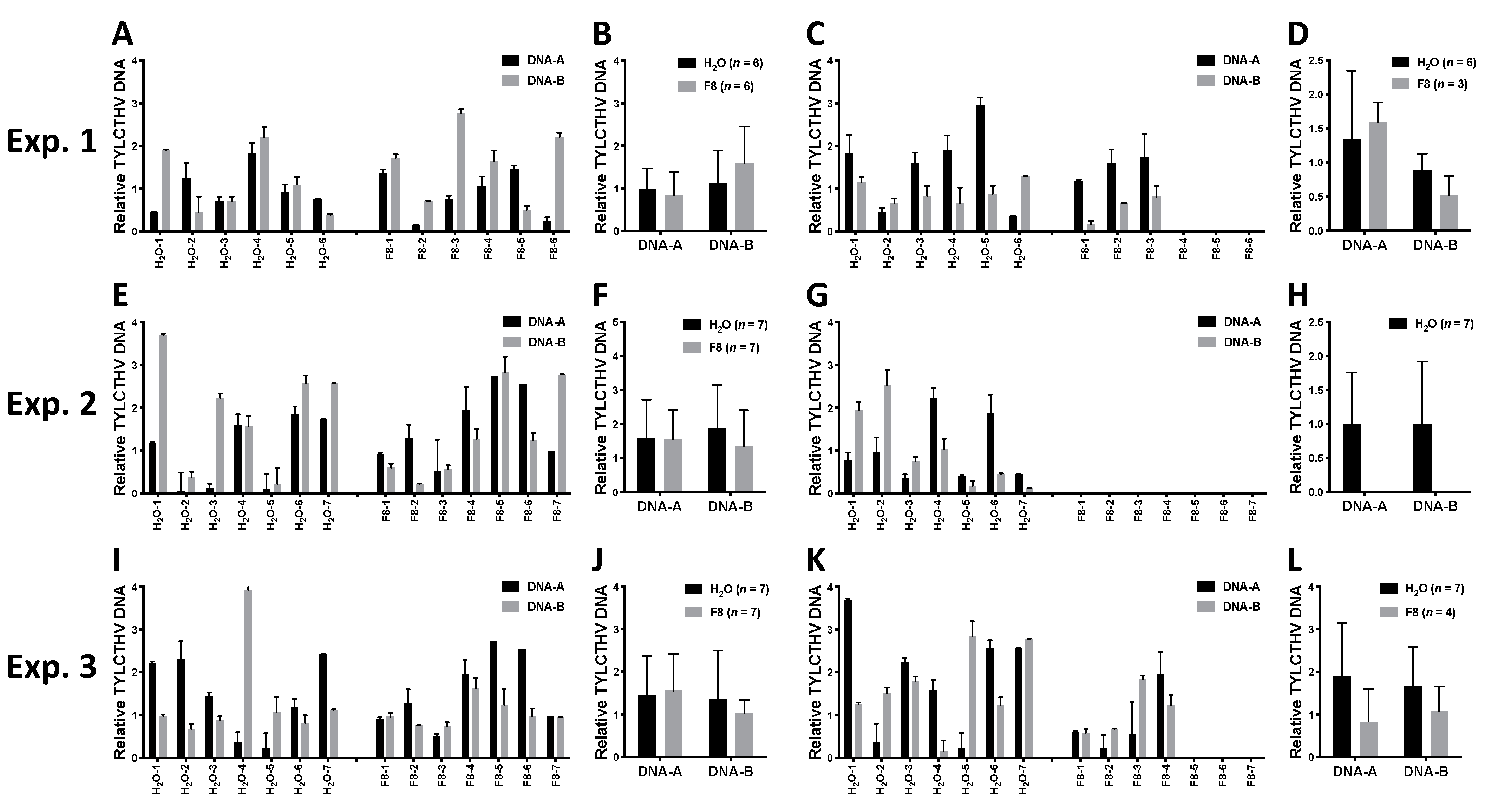
From the same 3 sets of experiments (Exp. 1, 2, and 3), the inoculated leaf and one systemic leaf from each plant (n = 6–7) were collected for evaluation of TYLCTHV accumulation in the H2O or F8-culture filtrate pretreated group. The relative accumulation of TYLCTHV genomic DNAs, DNA-A and -B, were quantified by quantitative PCR (qPCR). The results showed that TYLCTHV DNAs could be detected in inoculated leaves on plants pretreated with H2O or F8-culture filtrate (Figure 2A,B,E,F,I,J). On systemic leaves, TYLCTHV DNAs can be detected for both H2O or F8-culture filtrate pretreated plants that showed disease symptoms (Figure 1A,C,E and Figure 2C,D,G,H,K,L). However, TYLCTHV could not be detected on F8-culture filtrate pretreated healthy-looking plants (Figure 2C,G,K). Of note, even though we could observe healthy-looking plants in the H2O pretreated group in Exp. 2 (Figure 1C), we could still detect TYLCTHV on the systemic leaves of these plants (Figure 2G). The experiments were repeated for 3 times with similar results.
3.2. Mock and F8-Culture Filtrate Treated Plants Show Similar Fresh Weight
To determine whether F8-culture filtrate treatment affects the fresh weight in tomato, 3-week-old Yu-Nu were pretreated with H2O or F8-culture filtrate as mentioned above and analyzed the fresh weight of the tomato plants 5 weeks post treatment. We conducted 2 sets of experiments and analyzed 8–11 plants per treatment (H2O or F8-culture filtrate) in each of the experimental sets. We found that H2O and F8-culture filtrate-treated tomato showed similar fresh weight in our 2 replicated sets of experiments (Figure 3 and Figure S3).
3.3. F8-Culture Filtrate Mainly Induced the Expression of SA-Responsive Immune Marker Genes on Tomato Cultivar Yu-Nu
To understand the possible immune pathways involved in F8-culture filtrate-induced resistance against TYLCTHV, we detected the expression of phytohormone-responsive immune marker genes. A comparison of the F8-culture filtrate and H2O pretreated Yu-Nu tomato showed that the expression of SA-responsive immune marker gene, PR-1, increased 12.4-fold at 6 hpt, and was robustly induced by TYLCTHV to 82.8-fold at 4 dpi (Figure 4A). In addition, the expression of PR-5 increased 12.2-fold at 6 hpt, robustly increased 8.1-fold at 12 hpt, with the induction gradually decreasing to 6.1-fold at 24 hpt. Of note, after TYLCTHV infection, the PR-5 expression remained higher in F8-culture filtrate than H2O pretreated Yu-Nu tomatoes, but the induction fold was not as prominent as at 6, 12 and 24 hpt (Figure 4B).
In F8-culture filtrate-pretreated Yu-Nu tomato, we found that the expression of JA-responsive immune marker gene, OPDA reductase 3 (OPR3) [74], was increased at 24 hpt (1.5 fold) and 4 dpi (4.1 fold) (Figure 4C) compared to H2O pretreated Yu-Nu tomato, but the induction of another JA-responsive immune marker gene, STH2 [75], was not observed in H2O or F8-culture filtrate-pretreated Yu-Nu with or without TYLCTHV infection (Figure 4D).
Two ethylene (ET)-responsive immune marker genes, ethylene receptors-4 (ETR4) [76] and tomato transcription factor Pti4 [77], were also selected for analysis. The results showed that ETR4 only increased in F8-culture filtrate-pretreated Yu-Nu as compared to the H2O pretreated Yu-Nu control at 24 dpi (1.4 Fold) (Figure 4E), and the induction of Pti4 was not observed in H2O or F8-culture filtrate pretreated Yu-Nu with or without TYLCTHV infection (Figure 4F).
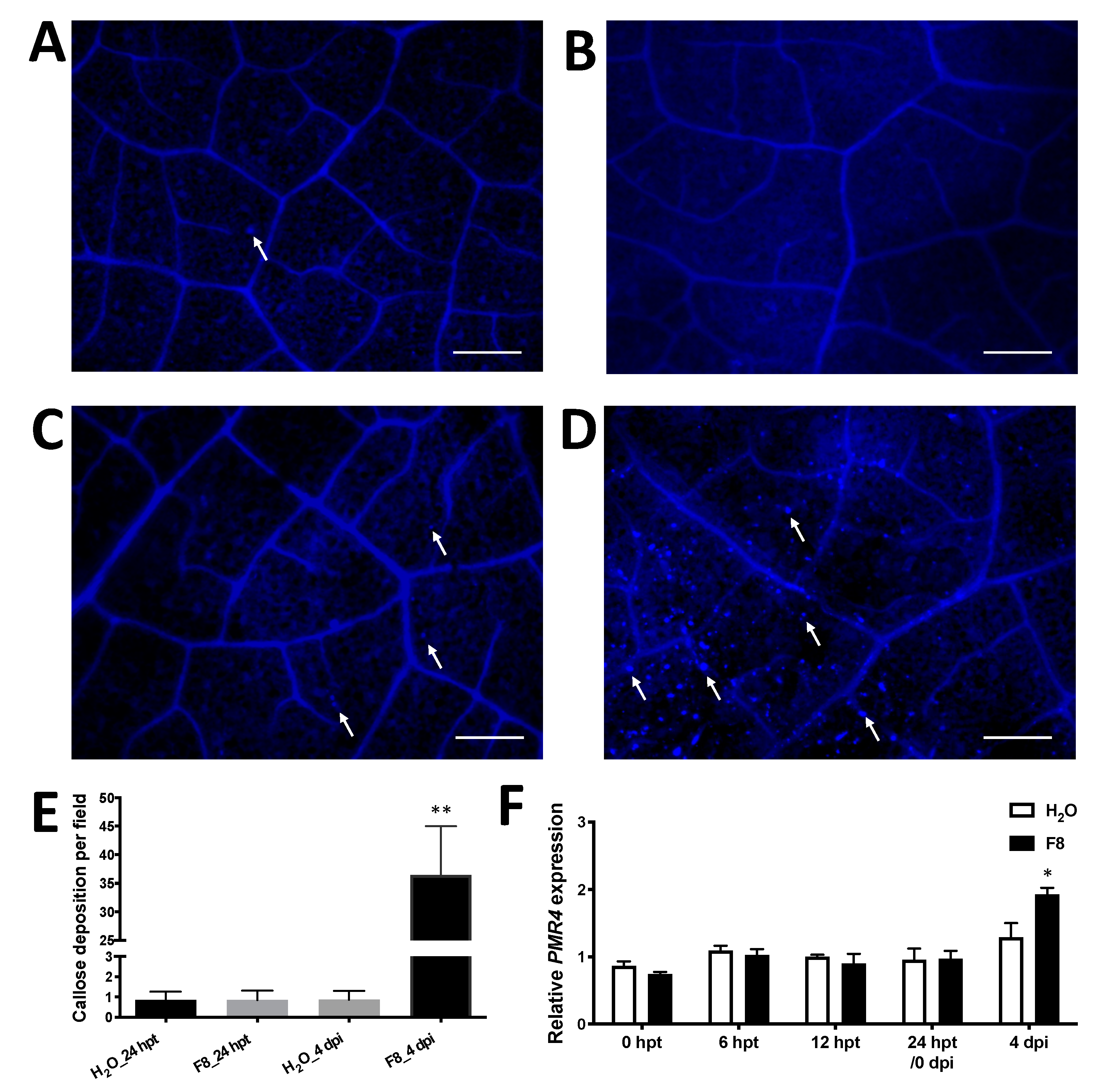
3.4. F8-Culture Filtrate Induced Callose Deposition on TYLCTHV-Inoculated Tomato Cultivar Yu-Nu
To understand how plants pretreated with F8-culture filtrate confine TYLCTHV infection (Figure 2), we analyzed whether callose deposition occurred in the tomato plants. Leaves were randomly collected from H2O or F8-culture filtrate pretreated plants at 24 hpt and stained with 0.2% aniline blue (Figure 5A,B). In addition, we also collected leaves from plants pretreated with H2O or F8-culture filtrate with TYLCTHV inoculation at 4 dpi for staining (Figure 5C,D). Fluorescence microscopy was used to examine the callose deposition under UV illumination. All images were used for quantification of callose deposition. We observed limited callose deposition on H2O or F8-culture filtrate pretreated plants at 24 hpt (Figure 5A,B,E). On TYLCTHV inoculated leaves, a limited number of callose depositions were observed on leaves pretreated with H2O (Figure 5C,E), but more profound callose depositions were observed only on leaves of plants pretreated with F8-culture filtrate (Figure 5D,E).
In addition to the staining of callose deposition, we also analyzed the expression of the pathogen-induced callose synthase gene, POWDERY MILDEW RESISTANT 4 (PMR4) [78]. Nucleic acids were extracted from collected leaves as mentioned above for callose staining. Significant induction of PMR4 was only observed on leaves of plants pretreated with F8-culture filtrate following TYLCTHV inoculation at 4 dpi (Figure 5F). The expression of PMR4 is consistent with the staining of callose deposition.
3.5. F8-Culture Filtrate Induced Virus Resistance on Tomato Cultivar Yu-Nu and Xiao-Nu in the Field
To analyze the effect of F8-culture filtrate on tomato against WTBs, we conducted two double-blind field trials on Yu-Nu and Xiao-Nu. The first field trial was conducted in Nantou County, and the data were recorded after planting in the field for 2 months. The results showed that much milder symptoms were observed on tomato cultivar Yu-Nu pretreated with F8-culture filtrate than with H2O pretreatment (Figure 6A,B).
In addition to Yu-Nu tomato, we also conducted field trials on Xiao-Nu in Alian District, Kaohsiung City. The results showed that much milder symptoms were observed on tomato cultivar Xiao-Nu pretreated with F8-culture filtrate than with H2O after planting in field for 2 months (Figure 6C,D). Collectively, our results indicated that F8-culture filtrate could induce effective resistance on different tomato cultivars against TYLCD in the field.
4. Discussion
The application of plant immunity inducers for crop protection is an environmentally sound approach, which can reduce the use of chemical pesticides to maintain healthy crops. The data presented in this study indicates that F8-culture filtrate is an effective method to induce tomato resistance against TYLCTHV both in the greenhouse and in the field (Figure 1 and Figure 2), with no obvious decrease in tomato growth (Figure 3). In addition, we also revealed the immune pathway(s) induced by F8-culture filtrate-pretreated tomato before or after TYLCTHV infection, which provides important information for understanding how plants can effectively ward-off infection by WTBs.
In designing the F8-culture filtrate, we have considered the cost, stability, and the method for application in the field. Instead of direct application of microbe(s) in the field, we chose F8-culture filtrate for practical application. This is to minimize environmental conditions that may affect the plant-microbe interaction and reduce the efficacy of induced resistance. F8-culture filtrate is made from inexpensive vegetable broth cultured with Trichosporon sp., and can be stored as a liquid at 4 °C for 2 months or as a lyophilized powder at 4 °C for at least 1 year. The powdered F8-culture filtrate can be dissolved in water before application [62]. For practical application, we only treated tomato seedlings with F8-culture filtrate before planting them in field, and no additional work was needed to induce resistance thereafter. However, profound enhanced resistance was found on F8-culture filtrate-treated tomato in the field (Figure 6).
We previously reported that F8-polysaccharide-treated N. benthamiana induced a typical immune priming expression pattern in both PR-1 and PR-2 [62]. Intriguingly, although F8-culture filtrate induced early induction of two tomato SA-mediated immune marker genes, PR-1 and PR-5, only PR-1 showed the typical immune priming expression pattern (Figure 4A). The expression of PR-5 was induced by F8-culture filtrate, but the induction fold was less after TYLCTHV inoculation than in the plants with F8-culture filtrate pretreatment (Figure 4B). This suggests that F8-culture filtrate can trigger the early SA-mediated immune pathway but only induce part of the SA-mediated immune priming. Alternatively, because we detected the PR-5 expression at 4 dpi, at that time TYLCTHV may already express effector(s) to compromise part of the immune response.
Previous reports have indicated that callose deposition has been found to confine virus infection by limiting cell-to-cell movement [79,80,81,82,83], and the SA-mediated immune pathway plays an important role in pathogen-induced callose deposition in Arabidopsis [81,84]. Consistently, our data indicated that F8-culture filtrate mainly induced the SA-mediated immune pathway and more callose deposits were found on tomato pretreated with F8-culture filtrate than with H2O (Figure 4A and Figure 5D). In addition, the gene involved in callose synthesis, PMR4, was up-regulated only in plants pretreated with F8-culture filtrate after TYLCTHV inoculation at 4 dpi (Figure 5F). Interestingly, the induction of callose depositions and PMR4 was not seen in plants pretreated with F8-culture filtrate before TYLCTHV inoculation (Figure 5B). This indicated that even though we did not detect the priming of callose depositions and PMR4 induction on F8-culture filtrate pretreated tomato before TYLCTHV inoculation, F8-culture filtrate primed the induction of tomato plants for callose depositions and PMR4 induction.
In comparison to expression of SA-mediated immune marker genes, only certain immune marker genes from the JA or ET pathways were induced in the F8-culture filtrate pretreated group as the expression of JA responsive gene, STH2, and ET responsive genes, ETR4 and Pti4, remained similar in all the treated groups. However, we found that a gene involved in synthesis of JA, OPR3, was induced at 24 hpt after F8-culture filtrate treatment and at 4 dpi after TYLCTHV inoculation on F8-culture filtrate pretreated tomato (Figure 4C). For the ethylene-mediated immune pathway, we found the gene ETR4, which functions as an ethylene receptor in tomato [85], is only induced in F8-culture filtrate pretreated tomato at 24 hpt (Figure 4E). This indicates inconsistent induction of JA- and ET-mediated immune marker genes. Notably, the induction of OPR3 and ETR4 was observed to be later than the induction of SA-mediated immune marker genes on F8-culture filtrate-treated tomato (Figure 4A,C,E). In addition, OPR3, but not ETR4, was induced after TYLCTHV inoculation (Figure 4C,E). The induction fold of OPR3 expression was higher at 4 dpi (4.1-fold) than at 24 hpt (1.5-fold). This indicates that OPR3 also showed a slight priming expression pattern.
Collectively, our data indicate that F8-culture filtrate is an effective inducer of tomato defense against TYLCTHV. In addition, we found that distinct SA immunity was induced compared to our previous report on N. benthamiana [52] This indicates that F8-culture filtrate mainly induces priming of SA immune response; however, the downstream immune responses vary among different plants. Together, our findings provide important practical information for application of F8-culture filtrate for tomato resistance against TYLCTHV and also provide insights for further enhancing SA-mediated plant immunity against viruses.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/v13081434/s1, Table S1: primer sequences, Figure S1: disease index of Yu-Nu tomato inoculated with tomato yellow leaf curl Thailand virus (TYLCTHV), Figure S2: symptom of Yu-Nu tomato inoculated with tomato yellow leaf curl Thailand virus (TYLCTHV). Figure S3: fresh weight of Yu-Nu tomato with H2O or F8-culture filtrate pretreatment.
Author Contributions
Conceptualization, H.-H.Y., Y.-S.C., and Y.T.; investigation, Y.-S.C. and Y.-H.C., methodology, H.-H.Y. and Y.-S.C.; resources, Y.-H.C. and C.-W.T.; supervision, H.-H.Y. and C.-W.T.; validation, Y.-S.C.; visualization, Y.-S.C. and Y.T.; writing—original draft preparation, H.-H.Y., Y.-S.C. and Y.T.; writing—review and editing, H.-H.Y., Y.-S.C., Y.T., and C.-W.T.; project administration, H.-H.Y. and C.-W.T.; funding acquisition, H.-H.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Innovative Translational Agricultural Research Grant Number AS-KPQ-107-ITAR-07, AS-KPQ-108-ITAR-07, AS-KPQ-109-ITAR-07 and AS-KPQ-110-ITAR-08.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting the findings of this study are available within the article and Supplementary Materials.
Acknowledgments
We would like to acknowledge the kind assistance of Wen-Ling Deng and Jen-Yu Tzeng, Department of Plant Pathology, National Chung Hsing University, in the field trial. We are also grateful to Miranda Loney for assistance with English editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brasesco, F.; Asgedom, D.; Casari, G. Strategic Analysis and Intervention Plan for Fresh and Industrial Tomato in the Agro-Commodities Procurement Zone of the Pilot Integrated Agro-Industrial Park in Central-Eastern Oromia, Ethiopia; FAO: Addis Ababa, Ethiopia, 2019. [Google Scholar]
- Prasad, A.; Sharma, N.; Hari-Gowthem, G.; Muthamilarasan, M.; Prasad, M. Tomato Yellow Leaf Curl Virus: Impact, Challenges, and Management. Trends Plant Sci. 2020, 25, 897–911. [Google Scholar] [CrossRef]
- Makkouk, K.M.; Shehab, S.; Majdalani, S.E. Tomato Yellow Leaf Curl: Incidence, Yield Losses and Transmission in Lebanon. J. Phytopathol. 1979, 96, 263–267. [Google Scholar] [CrossRef]
- Pico, M.B.; Diez, M.J.; Nuez, F. Viral diseases causing the greatest economic losses to the tomato crop. II. The Tomato yellow leaf curl virus—A review. Sci. Hortic. 1996, 67, 151–196. [Google Scholar] [CrossRef]
- Varma, A.; Malathi, V.G. Emerging geminivirus problems: A serious threat to crop production. Ann. Appl. Biol. 2003, 142, 145–164. [Google Scholar] [CrossRef]
- Yan, Z.; Wolters, A.-M.; Navas-Castillo, J.; Bai, Y. The Global Dimension of Tomato Yellow Leaf Curl Disease: Current Status and Breeding Perspectives. Microorganisms 2021, 9, 740. [Google Scholar] [CrossRef] [PubMed]
- Moriones, E.; Navas-Castillo, J. Tomato yellow leaf curl virus, an emerging virus complex causing epidemics worldwide. Virus Res. 2000, 71, 123–134. [Google Scholar] [CrossRef]
- Horowitz, A.R.; Kontsedalov, S.; Khasdan, V.; Ishaaya, I. Biotypes B and Q of Bemisia tabaci and their relevance to neonicotinoid and pyriproxyfen resistance. Arch. Insect Biochem. Physiol. 2005, 58, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, M.; Karniel, U.; Gelbart, D.; Fogel, D.; Evenor, D.; Kutsher, Y.; Makhbash, Z.; Nahon, S.; Shlomo, H.; Chen, L.; et al. A Novel Route Controlling Begomovirus Resistance by the Messenger RNA Surveillance Factor Pelota. PLoS Genet. 2015, 11, e1005538. [Google Scholar] [CrossRef] [PubMed]
- Czosnek, H.; Eybishtz, A.; Sade, D.; Gorovits, R.; Sobol, I.; Bejarano, E.; Rosas-Díaz, T.; Lozano-Durán, R. Discovering host genes involved in the infection by the Tomato Yellow Leaf Curl Virus complex and in the establishment of resistance to the virus using Tobacco Rattle Virus-based post transcriptional gene silencing. Viruses 2013, 5, 998–1022. [Google Scholar] [CrossRef] [Green Version]
- Elbaz, M.; Hanson, P.; Fgaier, S.; Laarif, A. Evaluation of tomato entries with different combinations of resistance genes to tomato yellow leaf curl disease in Tunisia. Plant Breed. 2016, 135, 525–530. [Google Scholar] [CrossRef]
- Anbinder, I.; Reuveni, M.; Azari, R.; Paran, I.; Nahon, S.; Shlomo, H.; Chen, L.; Lapidot, M.; Levin, I. Molecular dissection of Tomato leaf curl virus resistance in tomato line TY172 derived from Solanum peruvianum. Theor. Appl. Genet. 2009, 119, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Faal, P.G.; Farsi, M.; Seifi, A.; Kakhki, A.M. Virus-induced CRISPR-Cas9 system improved resistance against tomato yellow leaf curl virus. Mol. Biol. Rep. 2020, 47, 3369–3376. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, A.; Carlos, N.; Ruiz, Y.; Callard, D.; Sánchez, Y.; Ochagavía, M.E.; Seguin, J.; Malpica-López, N.; Hohn, T.; Lecca, M.R.; et al. Field Trial and Molecular Characterization of RNAi-Transgenic Tomato Plants That Exhibit Resistance to Tomato Yellow Leaf Curl Geminivirus. Mol. Plant Microbe Interact. 2016, 29, 197–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzean, Y.; Chang, H.-H.; Tu, T.-C.; Hou, B.H.; Chen, H.-M.; Chiu, Y.-S.; Chou, W.-Y.; Chang, L.; Yeh, H.-H. Engineering Plant Resistance to Tomato Yellow Leaf Curl Thailand Virus Using a Phloem-Specific Promoter Expressing Hairpin RNA. Mol. Plant Microbe Interact. 2020, 33, 87–97. [Google Scholar] [CrossRef]
- Asad, S.; Haris, W.A.A.; Bashir, A.; Zafar, Y.; Malik, K.A.; Malik, N.N.; Lichtenstein, C.P. Transgenic tobacco expressing geminiviral RNAs are resistant to the serious viral pathogen causing cotton leaf curl disease. Arch. Virol. 2003, 148, 2341–2352. [Google Scholar] [CrossRef]
- Chellappan, P.; Masona, M.V.; Vanitharani, R.; Taylor, N.J.; Fauquet, C.M. Broad Spectrum Resistance to ssDNA Viruses Associated with Transgene-Induced Gene Silencing in Cassava. Plant Mol. Biol. 2004, 56, 601–611. [Google Scholar] [CrossRef]
- Zhang, P.; Vanderschuren, H.; Futterer, J.; Gruissem, W. Resistance to cassava mosaic disease in transgenic cassava expressing antisense RNAs targeting virus replication genes. Plant Biotechnol. J. 2005, 3, 385–397. [Google Scholar] [CrossRef]
- Bonfim, K.; Faria, J.C.; Nogueira, E.O.P.L.; Mendes, É.A.; Aragão, F.J.L. RNAi-Mediated Resistance to Bean golden mosaic virus in Genetically Engineered Common Bean (Phaseolus vulgaris). Mol. Plant Microbe Interact. 2007, 20, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, S.; Mishra, A.; Praveen, S. Hairpin RNA-Mediated Strategies for Silencing of Tomato Leaf Curl Virus AC1 and AC4 Genes for Effective Resistance in Plants. Oligonucleotides 2007, 17, 251–257. [Google Scholar] [CrossRef]
- Ammara, E.U.; Mansoor, S.; Saeed, M.; Amin, I.; Briddon, R.W.; Al-Sadi, A.M. RNA interference-based resistance in transgenic tomato plants against Tomato yellow leaf curl virus-Oman (TYLCV-OM) and its associated betasatellite. Virol. J. 2015, 12, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.-Y.; Tsai, W.-S.; Ku, H.-M.; Jan, F.-J. Evaluation of DNA fragments covering the entire genome of a monopartite begomovirus for induction of viral resistance in transgenic plants via gene silencing. Transgenic Res. 2011, 21, 231–241. [Google Scholar] [CrossRef]
- Fuentes, A.; Ramos, P.L.; Fiallo, E.; Callard, D.; Sánchez, Y.; Peral, R.; Rodríguez, R.; Pujol, M.; Fiallo-Olivé, E. Intron–hairpin RNA Derived from Replication Associated Protein C1 Gene Confers Immunity to Tomato Yellow Leaf Curl Virus Infection in Transgenic Tomato Plants. Transgenic Res. 2006, 15, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sherwood, T.A.; Patte, C.P.; Hiebert, E.; Polston, J.E. Use of Tomato yellow leaf curl virus (TYLCV) Rep Gene Sequences to Engineer TYLCV Resistance in Tomato. Phytopathology 2004, 94, 490–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Feng, B.; He, P.; Shan, L. From Chaos to Harmony: Responses and Signaling upon Microbial Pattern Recognition. Annu. Rev. Phytopathol. 2017, 55, 109–137. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Klessig, D.F. DAMPs, MAMPs, and NAMPs in plant innate immunity. BMC Plant Biol. 2016, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Schwessinger, B.; Ronald, P.C. Plant Innate Immunity: Perception of Conserved Microbial Signatures. Annu. Rev. Plant Biol. 2012, 63, 451–482. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.L. Distillation Control: An Engineering Perspective; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 70–135. [Google Scholar]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Couto, D.; Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 2016, 16, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Spoel, S.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Parker, L.L. A time-resolved luminescence biosensor assay for anaplastic lymphoma kinase (ALK) activity. Chem. Commun. 2014, 51, 362–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alazem, M.; Lin, N. Roles of plant hormones in the regulation of host–virus interactions. Mol. Plant Pathol. 2015, 16, 529–540. [Google Scholar] [CrossRef]
- Asai, T.; Tena, G.; Plotnikova, J.; Willmann, M.R.; Chiu, W.-L.; Gomez-Gomez, L.; Boller, T.; Ausubel, F.M.; Sheen, J. MAP kinase signalling cascade in Arabidopsis innate immunity. Nat. Cell Biol. 2002, 415, 977–983. [Google Scholar] [CrossRef]
- Ausubel, F.M. Are innate immune signaling pathways in plants and animals conserved? Nat. Immunol. 2005, 6, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; Felix, G. A Renaissance of Elicitors: Perception of Microbe-Associated Molecular Patterns and Danger Signals by Pattern-Recognition Receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef] [PubMed]
- Pitzschke, A.; Djamei, A.; Teige, M.; Hirt, H. VIP1 response elements mediate mitogen-activated protein kinase 3-induced stress gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 18414–18419. [Google Scholar] [CrossRef] [Green Version]
- Nicaise, V. Crop immunity against viruses: Outcomes and future challenges. Front. Plant Sci. 2014, 5, 660. [Google Scholar] [CrossRef]
- Zhou, J.-M.; Chai, J. Plant pathogenic bacterial type III effectors subdue host responses. Curr. Opin. Microbiol. 2008, 11, 179–185. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, J.-M. Plant Immunity Triggered by Microbial Molecular Signatures. Mol. Plant 2010, 3, 783–793. [Google Scholar] [CrossRef] [Green Version]
- Toruño, T.Y.; Stergiopoulos, I.; Coaker, G. Plant-Pathogen Effectors: Cellular Probes Interfering with Plant Defenses in Spatial and Temporal Manners. Annu. Rev. Phytopathol. 2016, 54, 419–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, S.R.; Fisher, E.J.; Chang, J.H.; Mole, B.M.; Dangl, J.L. Subterfuge and Manipulation: Type III Effector Proteins of Phytopathogenic Bacteria. Annu. Rev. Microbiol. 2006, 60, 425–449. [Google Scholar] [CrossRef] [Green Version]
- Abramovitch, R.; Janjusevic, R.; Stebbins, E.; Martin, G.B. Type III effector AvrPtoB requires intrinsic E3 ubiquitin ligase activity to suppress plant cell death and immunity. Proc. Natl. Acad. Sci. USA 2006, 103, 2851–2856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, F.; Vernaldi, S.; Maekawa, T. Evolution and Conservation of Plant NLR Functions. Front. Immunol. 2013, 4, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuda, K.; Katagiri, F. Comparing signaling mechanisms engaged in pattern-triggered and effector-triggered immunity. Curr. Opin. Plant Biol. 2010, 13, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.; Dangl, J.L. Arabidopsis and the plant immune system. Plant J. 2010, 61, 1053–1066. [Google Scholar] [CrossRef] [Green Version]
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the Plant Immune System from Dissection to Deployment. Science 2013, 341, 746–751. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Zeng, H.; Qiu, D.; Guo, L.; Yang, X.; Shi, H.; Zhou, T.; Zhao, J. Purification and Characterization of a Novel Hypersensitive Response-Inducing Elicitor from Magnaporthe oryzae that Triggers Defense Response in Rice. PLoS ONE 2012, 7, e37654. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Zhang, C.; Zi, Q.; Qiu, D.; Liu, W.; Zeng, H. A novel elicitor identified from Magnaporthe oryzae triggers defense responses in tobacco and rice. Plant Cell Rep. 2014, 33, 1865–1879. [Google Scholar] [CrossRef]
- Wang, L.; Yang, X.; Zeng, H.; Qiu, D.; Guo, L.; Liu, Z. Extracellular expression of protein elicitor PeaT1 in Bacillus subtilis to enhance drought tolerance and growth in wheat. Sheng Wu Gong Cheng Xue Bao 2011, 27, 1355–1362. [Google Scholar] [PubMed]
- Wang, N.; Liu, M.; Guo, L.; Yang, X.; Qiu, D. A Novel Protein Elicitor (PeBA1) from Bacillus amyloliquefaciens NC6 Induces Systemic Resistance in Tobacco. Int. J. Biol. Sci. 2016, 12, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, X.; Zeng, H.; Guo, L.; Yuan, J.; Qiu, D. Fungal elicitor protein PebC1 from Botrytis cinerea improves disease resistance in Arabidopsis thaliana. Biotechnol. Lett. 2014, 36, 1069–1078. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Qiu, D.; Zeng, H.; Guo, L.; Yang, X. BcGs1, a glycoprotein from Botrytis cinerea, elicits defence response and improves disease resistance in host plants. Biochem. Biophys. Res. Commun. 2015, 457, 627–634. [Google Scholar] [CrossRef]
- Pichyangkura, R.; Chadchawan, S. Biostimulant activity of chitosan in horticulture. Sci. Hortic. 2015, 196, 49–65. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djonović, S.; Pozo, M.J.; Dangott, L.J.; Howell, C.R.; Kenerley, C.M. Sm1, a Proteinaceous Elicitor Secreted by the Biocontrol Fungus Trichoderma virens Induces Plant Defense Responses and Systemic Resistance. Mol. Plant Microbe Interact. 2006, 19, 838–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perazzolli, M.; Roatti, B.; Bozza, E.; Pertot, I. Trichoderma harzianum T39 induces resistance against downy mildew by priming for defense without costs for grapevine. Biol. Control 2011, 58, 74–82. [Google Scholar] [CrossRef]
- Seidl, V.; Marchetti, M.; Schandl, R.; Allmaier, G.; Kubicek, C.P.; Marchetti-Deschmann, M. Epl1, the major secreted protein of Hypocrea atroviridis on glucose, is a member of a strongly conserved protein family comprising plant defense response elicitors. FEBS J. 2006, 273, 4346–4359. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.-A.; Weng, S.-H.; Chen, M.-C.; Lin, J.-S.; Tsai, W.-S. Priming of Plant Resistance to Heat Stress and Tomato Yellow Leaf Curl Thailand Virus With Plant-Derived Materials. Front. Plant Sci. 2019, 10, 906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heijari, J.; Nerg, A.; Kainulainen, P.; Viiri, H.; Vuorinen, M.; Holopainen, J. Application of methyl jasmonate reduces growth but increases chemical defence and resistance against Hylobius abietis in Scots pine seedlings. Entomol. Exp. Appl. 2005, 115, 117–124. [Google Scholar] [CrossRef]
- Boughton, A.J.; Hoover, K.; Felton, G.W. Impact of chemical elicitor applications on greenhouse tomato plants and population growth of the green peach aphid, Myzus persicae. Entomol. Exp. Appl. 2006, 120, 175–188. [Google Scholar] [CrossRef]
- Chiu, Y.-S.; Chen, P.-Y.; Kuan, T.; Wang, P.-C.; Chen, Y.-J.; Yang, Y.-L.; Yeh, H.-H. A Polysaccharide Derived from a Trichosporon sp. Culture Strongly Primes Plant Resistance to Viruses. Mol. Plant Microbe Interact. 2018, 31, 1257–1270. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Huang, Y.; Xu, Z.-S.; Wang, F.; Xiong, A.-S. Salicylic acid-induced differential resistance to the Tomato yellow leaf curl virus among resistant and susceptible tomato cultivars. BMC Plant Biol. 2019, 19, 173. [Google Scholar] [CrossRef]
- Shih, S.L.; Tsai, W.S.; Lee, L.M.; Wang, J.T.; Green, S.K.; Kenyon, L. First Report of Tomato yellow leaf curl Thailand virus Associated with Pepper Leaf Curl Disease in Taiwan. Plant Dis. 2010, 94, 637. [Google Scholar] [CrossRef]
- Jan, F.-J.; Green, S.K.; Shih, S.L.; Lee, L.M.; Ito, H.; Kimbara, J.; Hosoi, K.; Tsai, W.S. First Report of Tomato yellow leaf curl Thailand virus in Taiwan. Plant Dis. 2007, 91, 1363. [Google Scholar] [CrossRef]
- Tsai, W.S.; Shih, S.L.; Kenyon, L.; Green, S.K.; Jan, F.-J. Temporal distribution and pathogenicity of the predominant tomato-infecting begomoviruses in Taiwan. Plant Pathol. 2011, 60, 787–799. [Google Scholar] [CrossRef]
- Weng, S.; Tsai, W.; Kenyon, L.; Tsai, C.-W. Different transmission efficiencies may drive displacement of tomato begomoviruses in the fields in Taiwan. Ann. Appl. Biol. 2015, 166, 321–330. [Google Scholar] [CrossRef]
- Hu, F.-Y.; Mou, D.-F.; Tsai, C.-W. Evaluation of barrier plants for the cultural control of tomato yellow leaf curl disease. J. Asia-Pac. Entomol. 2020, 23, 132–137. [Google Scholar] [CrossRef]
- Li, W.-H.; Mou, D.-F.; Hsieh, C.-K.; Weng, S.-H.; Tsai, W.-S.; Tsai, C.-W. Vector Transmission of Tomato Yellow Leaf Curl Thailand Virus by the Whitefly Bemisia tabaci: Circulative or Propagative? Insects 2021, 12, 181. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Anfoka, G.; Moshe, A.; Fridman, L.; Amrani, L.; Rotem, O.; Kolot, M.; Zeidan, M.; Czosnek, H.; Gorovits, R. Tomato yellow leaf curl virus infection mitigates the heat stress response of plants grown at high temperatures. Sci. Rep. 2016, 6, 19715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moshe, A.; Gorovits, R.; Liu, Y.; Czosnek, H. Tomato plant cell death induced by inhibition of HSP90 is alleviated by Tomato yellow leaf curl virus infection. Mol. Plant Pathol. 2016, 17, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Ning, W.; Shi, X.; Liu, B.; Pan, H.; Wei, W.; Zeng, Y.; Sun, X.; Xie, W.; Wang, S.; Wu, Q.; et al. Transmission of Tomato Yellow Leaf Curl Virus by Bemisia tabaci as Affected by Whitefly Sex and Biotype. Sci. Rep. 2015, 5, srep10744. [Google Scholar] [CrossRef]
- Sun, Y.-C.; Pan, L.-L.; Ying, F.-Z.; Li, P.; Wang, X.-W.; Liu, S.-S. Jasmonic acid-related resistance in tomato mediates interactions between whitefly and whitefly-transmitted virus. Sci. Rep. 2017, 7, 566. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Du, M.; Deng, L.; Shen, J.; Fang, M.; Chen, Q.; Lu, Y.; Wang, Q.; Li, C.; Zhai, Q. MYC2 Regulates the Termination of Jasmonate Signaling via an Autoregulatory Negative Feedback Loop. Plant Cell 2019, 31, 106–127. [Google Scholar] [CrossRef] [Green Version]
- Martín-Rodríguez, J.Á.; León-Morcillo, R.; Vierheilig, H.; Ocampo, J.A.; Ludwig-Müller, J.; García-Garrido, J.M. Ethylene-dependent/ethylene-independent ABA regulation of tomato plants colonized by arbuscular mycorrhiza fungi. New Phytol. 2011, 190, 193–205. [Google Scholar] [CrossRef]
- Li, R.; Wang, L.; Li, Y.; Zhao, R.; Zhang, Y.; Sheng, J.; Ma, P.; Shen, L. Knockout of SlNPR1 enhances tomato plants resistance against Botrytis cinerea by modulating ROS homeostasis and JA/ET signaling pathways. Physiol. Plant. 2020, 170, 569–579. [Google Scholar] [CrossRef]
- Jacobs, A.K.; Lipka, V.; Burton, R.; Panstruga, R.; Strizhov, N.; Schulze-Lefert, P.; Fincher, G.B. An Arabidopsis Callose Synthase, GSL5, Is Required for Wound and Papillary Callose Formation. Plant Cell 2003, 15, 2503–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alazem, M.; Lin, N.-S. Antiviral Roles of Abscisic Acid in Plants. Front. Plant Sci. 2017, 8, 1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.-Y.; Kim, J.-Y. Callose synthesis in higher plants. Plant Signal. Behav. 2009, 4, 489–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epel, B.L. Plant viruses spread by diffusion on ER-associated movement-protein-rafts through plasmodesmata gated by viral induced host beta-1,3-glucanases. Semin. Cell Dev. Biol. 2009, 20, 1074–1081. [Google Scholar]
- Hofmann, J.; Banora, M.; De Almeida-Engler, J.; Grundler, F.M.W. The Role of Callose Deposition Along Plasmodesmata in Nematode Feeding Sites. Mol. Plant Microbe Interact. 2010, 23, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhao, Y.; Liu, C.; Yao, G.; Wu, S.; Hou, C.; Zhang, M.; Wang, D. Callose deposition at plasmodesmata is a critical factor in restricting the cell-to-cell movement of Soybean mosaic virus. Plant Cell Rep. 2012, 31, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sager, R.; Cui, W.; Zhang, C.; Lu, H.; Lee, J.Y. Salicylic Acid Regulates Plasmodesmata Closure during Innate Immune Responses in Arabidopsis. Plant Cell 2013, 25, 2315–2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kevany, B.M.; Tieman, D.M.; Taylor, M.G.; Cin, V.D.; Klee, H.J. Ethylene receptor degradation controls the timing of ripening in tomato fruit. Plant J. 2007, 51, 458–467. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Symptoms on H2O or F8-culture filtrate pretreated Yu-Nu tomato plants inoculated with tomato yellow leaf curl Thailand virus (TYLCTHV). The level of disease index was recorded at 28 days post inoculation (dpi) in the H2O or F8-culture filtrate pretreated group (A,C,E). The average disease index of plants inoculated with TYLCTHV according to the specified pretreatment groups (B,D,F). Data represent mean ± SEM; n = 6–7 biological replicates. Student’s t-test was used for comparative analysis of the data for the F8-culture filtrate pretreated group and the H2O pretreated control group. * p < 0.05; ** p < 0.01.
Figure 1.
Symptoms on H2O or F8-culture filtrate pretreated Yu-Nu tomato plants inoculated with tomato yellow leaf curl Thailand virus (TYLCTHV). The level of disease index was recorded at 28 days post inoculation (dpi) in the H2O or F8-culture filtrate pretreated group (A,C,E). The average disease index of plants inoculated with TYLCTHV according to the specified pretreatment groups (B,D,F). Data represent mean ± SEM; n = 6–7 biological replicates. Student’s t-test was used for comparative analysis of the data for the F8-culture filtrate pretreated group and the H2O pretreated control group. * p < 0.05; ** p < 0.01.

Figure 2.
Quantification of TYLCTHV genomic DNA in inoculated tomato plants pretreated with F8-culture filtrate or H2O. Relative TYLCTHV DNA level was quantified from H2O or F8-culture filtrate pretreated plants corresponding to Exp. 1, 2, and 3 in Figure 1 by qPCR analysis. TYLCTHV DNA was detected from individual samples of TYLCTHV-inoculated leaves (A,E,I) or systemic leaves (C,G,K). Three technical replicates were performed for all samples. Data represent mean ± SEM. β-actin was used as an input control. The average of TYLCTHV DNA detected from (A,E,J) are shown in (B,F,J), respectively. The average of TYLCTHV DNA detected from (C,G,K) are shown in (D,H,L), respectively. Data represent mean ± SEM; n = 3–6 biological replicates. β-actin was used as an input control. Student’s t-test was used for comparative analysis of the data for the F8-culture filtrate pretreated group and the H2O pretreated control group.
Figure 2.
Quantification of TYLCTHV genomic DNA in inoculated tomato plants pretreated with F8-culture filtrate or H2O. Relative TYLCTHV DNA level was quantified from H2O or F8-culture filtrate pretreated plants corresponding to Exp. 1, 2, and 3 in Figure 1 by qPCR analysis. TYLCTHV DNA was detected from individual samples of TYLCTHV-inoculated leaves (A,E,I) or systemic leaves (C,G,K). Three technical replicates were performed for all samples. Data represent mean ± SEM. β-actin was used as an input control. The average of TYLCTHV DNA detected from (A,E,J) are shown in (B,F,J), respectively. The average of TYLCTHV DNA detected from (C,G,K) are shown in (D,H,L), respectively. Data represent mean ± SEM; n = 3–6 biological replicates. β-actin was used as an input control. Student’s t-test was used for comparative analysis of the data for the F8-culture filtrate pretreated group and the H2O pretreated control group.

Figure 3.
Fresh weight of Yu-Nu tomato with H2O or F8-culture filtrate pretreatment. (A) The average fresh weight was recorded for each treatment group (n = 11). (B) The phenotype of H2O or F8-culture filtrate at 5 weeks post treatment.
Figure 3.
Fresh weight of Yu-Nu tomato with H2O or F8-culture filtrate pretreatment. (A) The average fresh weight was recorded for each treatment group (n = 11). (B) The phenotype of H2O or F8-culture filtrate at 5 weeks post treatment.

Figure 4.
The expression of phytohormone-mediated immune marker genes in F8-culture filtrate or H2O pretreated Yu-Nu tomato before or after TYLCTHV inoculation. Tomato plants were pretreated with H2O or F8-culture filtrate and inoculated with TYLCTHV using whitefly vector at 24 hpt. The relative expression of SA-responsive gene PR-1 (A) and PR-5 (B), jasmonic acid (JA)-biosynthesis gene OPR3 (C), JA-responsive gene STH2 (D), and ethylene (ET)-responsive gene ETR4 (E) and Pti4 (F) were detected by qRT-PCR at 0, 12 and 24 hpt or at 4 dpi. β-actin was used as an input control. Data are mean ± SEM of 3 biological replicates. Student’s t-test was used for comparative analysis of the data for the H2O and F8-culture filtrate pretreated groups. * p < 0.05; ** p < 0.01, *** p < 0.005, **** p < 0.001.
Figure 4.
The expression of phytohormone-mediated immune marker genes in F8-culture filtrate or H2O pretreated Yu-Nu tomato before or after TYLCTHV inoculation. Tomato plants were pretreated with H2O or F8-culture filtrate and inoculated with TYLCTHV using whitefly vector at 24 hpt. The relative expression of SA-responsive gene PR-1 (A) and PR-5 (B), jasmonic acid (JA)-biosynthesis gene OPR3 (C), JA-responsive gene STH2 (D), and ethylene (ET)-responsive gene ETR4 (E) and Pti4 (F) were detected by qRT-PCR at 0, 12 and 24 hpt or at 4 dpi. β-actin was used as an input control. Data are mean ± SEM of 3 biological replicates. Student’s t-test was used for comparative analysis of the data for the H2O and F8-culture filtrate pretreated groups. * p < 0.05; ** p < 0.01, *** p < 0.005, **** p < 0.001.

Figure 5.
Staining of callose deposition in F8-culture filtrate or H2O pretreated Yu-Nu tomato before or after TYLCTHV inoculation. Tomato plants were pretreated with H2O (A,C) or F8-culture filtrate (B,D). The pretreated plants were inoculated with TYLCTHV using whitefly vector at 24 hpt. The callose deposition was stained with 0.02% aniline blue. Callose depositions were analyzed at 24 hpt without TYLCTHV infection (A,B) or with TYLCTHV infection at 4 dpi (C,D) using a Zeiss Imager Z1 fluorescence microscope. Scale bar = 200 μm. White arrow indicates callose depositions. (E) The average numbers of callose deposition per field. Two pictures were taken per leaf (TYLCTHV-inoculated) collected from each plant (n = 7 plants) from H2O or F8-culture filtrate treatment for analysis. Data shows mean number of callose deposition per field ± SEM. Student’s t-test was used to analyze the data from the F8-culture filtrate pretreated group and the H2O-pretreated control group. ** p < 0.01. (F) The relative expression of callose synthase gene PMR4 was detected by qRT-PCR at 0, 12 and 24 hpt without TYLCTHV infection or with TYLCTHV infection at 4 dpi. Data are mean ± SEM from 3 plants. β-actin was used as an input control. Student’s t-test was used for comparative analysis of the data in the H2O and F8-culture filtrate pretreated groups. * p < 0.05.
Figure 5.
Staining of callose deposition in F8-culture filtrate or H2O pretreated Yu-Nu tomato before or after TYLCTHV inoculation. Tomato plants were pretreated with H2O (A,C) or F8-culture filtrate (B,D). The pretreated plants were inoculated with TYLCTHV using whitefly vector at 24 hpt. The callose deposition was stained with 0.02% aniline blue. Callose depositions were analyzed at 24 hpt without TYLCTHV infection (A,B) or with TYLCTHV infection at 4 dpi (C,D) using a Zeiss Imager Z1 fluorescence microscope. Scale bar = 200 μm. White arrow indicates callose depositions. (E) The average numbers of callose deposition per field. Two pictures were taken per leaf (TYLCTHV-inoculated) collected from each plant (n = 7 plants) from H2O or F8-culture filtrate treatment for analysis. Data shows mean number of callose deposition per field ± SEM. Student’s t-test was used to analyze the data from the F8-culture filtrate pretreated group and the H2O-pretreated control group. ** p < 0.01. (F) The relative expression of callose synthase gene PMR4 was detected by qRT-PCR at 0, 12 and 24 hpt without TYLCTHV infection or with TYLCTHV infection at 4 dpi. Data are mean ± SEM from 3 plants. β-actin was used as an input control. Student’s t-test was used for comparative analysis of the data in the H2O and F8-culture filtrate pretreated groups. * p < 0.05.

Figure 6.
Symptoms on H2O or F8-culture filtrate treated tomato in the field in Nantou County and Kaohsiung City. Tomato seeds were sown and grown in a growth chamber for 3 weeks. Yu-Nu tomato and Xiao-Nu tomatoes were pretreated with H2O or F8 by spraying three times with a 24-h interval between each spray, then transplanted to fields in Yuchi Township, Nantou County, or Alian Dist., Kaohsiung City. The level of disease index was recorded at 2-months post transplantation (A,C). The average of the disease index (B,D). Data represent mean ± SEM. Student’s t-test was used for analysis of the data in the F8-culture filtrate pretreated group and the H2O-pretreated control group. ** p < 0.01; **** p < 0.001.
Figure 6.
Symptoms on H2O or F8-culture filtrate treated tomato in the field in Nantou County and Kaohsiung City. Tomato seeds were sown and grown in a growth chamber for 3 weeks. Yu-Nu tomato and Xiao-Nu tomatoes were pretreated with H2O or F8 by spraying three times with a 24-h interval between each spray, then transplanted to fields in Yuchi Township, Nantou County, or Alian Dist., Kaohsiung City. The level of disease index was recorded at 2-months post transplantation (A,C). The average of the disease index (B,D). Data represent mean ± SEM. Student’s t-test was used for analysis of the data in the F8-culture filtrate pretreated group and the H2O-pretreated control group. ** p < 0.01; **** p < 0.001.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chiu, Y.-S.; Tzean, Y.; Chen, Y.-H.; Tsai, C.-W.; Yeh, H.-H. Fungal F8-Culture Filtrate Induces Tomato Resistance against Tomato Yellow Leaf Curl Thailand Virus. Viruses 2021, 13, 1434. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081434
AMA Style
Chiu Y-S, Tzean Y, Chen Y-H, Tsai C-W, Yeh H-H. Fungal F8-Culture Filtrate Induces Tomato Resistance against Tomato Yellow Leaf Curl Thailand Virus. Viruses. 2021; 13(8):1434. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081434
Chicago/Turabian StyleChiu, Yi-Shu, Yuh Tzean, Yi-Hui Chen, Chi-Wei Tsai, and Hsin-Hung Yeh. 2021. "Fungal F8-Culture Filtrate Induces Tomato Resistance against Tomato Yellow Leaf Curl Thailand Virus" Viruses 13, no. 8: 1434. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081434
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.