
Influenza Virus Infections in Cats

 , ,
, ,  , , , ,
, , , ,  ,
, 
{kind=link}
Abstract
:1. Introduction
2. Aetiology
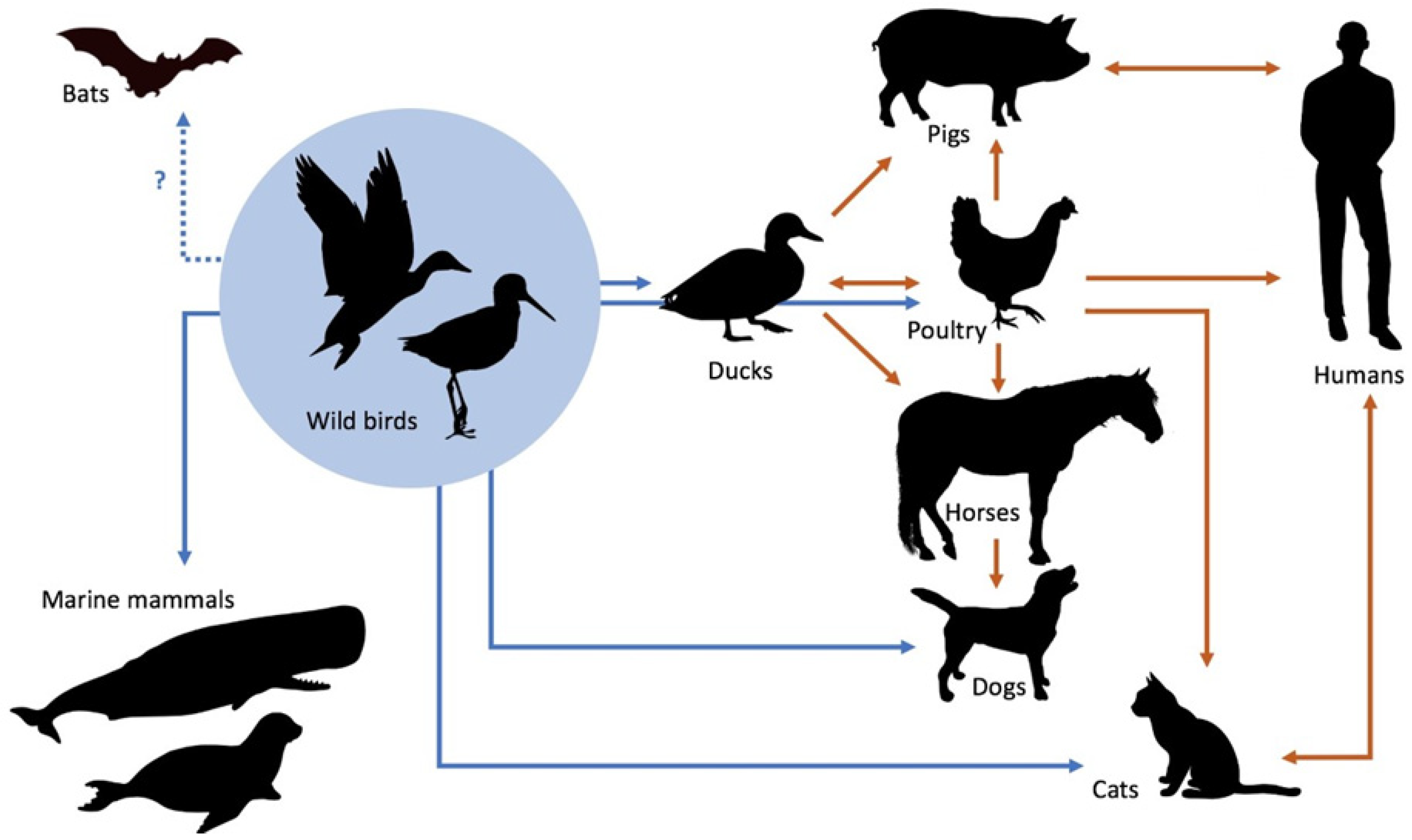
3. Circulation of IAVs
4. Pathogenesis
5. Epidemiology of IAV Infections in Cats
5.1. Low Pathogenic IAVs
5.2. Highly Pathogenic H5N1 IAV
6. Clinical Signs
7. Pathological Lesions
8. Diagnosis
9. Control
10. Human Risks
Funding
Acknowledgments
Conflicts of Interest
References
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Primers 2018, 28, 3. [Google Scholar] [CrossRef] [PubMed]
- Thiry, E.; Zicola, A.; Addie, D.; Egberink, H.; Hartmann, K.; Lutz, H.; Poulet, H.; Horzinek, M. Highly pathogenic avian influenza H5N1 virus in cats and other carnivores. Vet. Microbiol. 2007, 122, 25–31. [Google Scholar] [CrossRef]
- Yang, W.; Schountz, T.; Ma, W. Bat influenza viruses: Current status and perspective. Viruses 2021, 13, 547. [Google Scholar] [CrossRef] [PubMed]
- Wille, M.; Holmes, E.C. The ecology and evolution of influenza viruses. Cold Spring Harb. Perspect. Med. 2020, 10, a038489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Trovão, N.S.; Wang, G.; Zhao, W.; He, P.; Zhou, H.; Mo, Y.; Wei, Z.; Ouyang, K.; Huang, W.; et al. Emergence and evolution of novel reassortant influenza A viruses in canines in southern China. mBio 2018, 9, e00909-18. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Schuurman, N.; Tieke, M.; Quist, B.; Zwinkels, S.; van Kuppeveld, F.; de Haan, C.A.M.; Egberink, H. Serological screening of influenza A virus antibodies in cats and dogs indicates frequent infection with different subtypes. J. Clin. Microbiol. 2020, 58, e01689-20. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Li, G.; Wang, R.; Shi, W.; Li, K.; Wang, S.; Lai, A.; Su, S. Host-range shift of H3N8 canine influenza virus: A phylodynamic analysis of its origin and adaptation from equine to canine host. Vet. Res. 2019, 50, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irwin, C.K.; Yoon, K.J.; Wang, C.; Hoff, S.J.; Zimmerman, J.J.; Denagamage, T.; O’Connor, A.M. Using the systematic review methodology to evaluate factors that influence the persistence of influenza virus in environmental matrices. Appl. Environ. Microbiol. 2011, 77, 1049–1060. [Google Scholar] [CrossRef] [Green Version]
- Keeler, S.P.; Berghaus, R.D.; Stallknecht, D.E. Persistence of low pathogenic avian influenza viruses in filtered surface water from waterfowl habitats in Georgia, USA. J. Wildl. Dis. 2012, 48, 999–1009. [Google Scholar] [CrossRef] [Green Version]
- Sonnberg, S.; Webby, R.J.; Webster, R.G. Natural history of highly pathogenic avian influenza H5N1. Virus Res. 2013, 178, 63–77. [Google Scholar] [CrossRef] [Green Version]
- Crawford, P.C.; Dubovi, E.J.; Castleman, W.L.; Stephenson, I.; Gibbs, E.P.J.; Chen, L.; Smith, C.; Hill, R.C.; Ferro, P.; Pompey, J.; et al. Transmission of equine influenza virus to dogs. Science 2005, 310, 482–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, P.C.; Dubovi, E.J.; Castleman, W.L.; Stephenson, I.; Gibbs, E.P.; Chen, L.; Smith, C.; Hill, R.C.; Ferro, P.; Pompey, J.; et al. Infection of dogs with equine influenza virus: Evidence for transmission from horses during the Australian outbreak. Aust. Vet. J. 2011, 89, 27–28. [Google Scholar] [CrossRef]
- Song, D.; Kang, B.; Lee, C.; Jung, K.; Ha, G.; Kang, D.; Park, S.; Park, B.; Oh, J. Transmission of avian influenza virus (H3N2) to dogs. Emerg. Infect. Dis. 2008, 14, 741–746. [Google Scholar] [CrossRef]
- Newbury, S.P.; Cigel, F.; Killian, M.L.; Leutenegger, C.M.; Seguin, M.A.; Crossley, B.; Brennen, R.; Suarez, D.L.; Torchetti, M.; Toohey-Kurth, K. First detection of avian lineage H7N2 in Felis catus. Genome Announc. 2017, 5, e00457-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, R.G. Are equine 1 influenza viruses still present in horses? Equine Vet. J. 1993, 25, 537–538. [Google Scholar] [CrossRef] [PubMed]
- Wasik, B.R.; Voorhees, I.E.H.; Parrish, C.R. Canine and feline influenza. Cold Spring Harb. Perspect. Med. 2021, 11, a038562. [Google Scholar] [CrossRef] [Green Version]
- Song, D.S.; An, D.J.; Moon, H.J.; Yeom, M.J.; Jeong, H.Y.; Jeong, W.S.; Park, S.J.; Kim, H.K.; Han, S.Y.; Oh, J.S.; et al. Interspecies transmission of the canine influenza H3N2 virus to domestic cats in South Korea, 2010. J. Gen. Virol. 2011, 92, 2350–2355. [Google Scholar] [CrossRef]
- Fiorentini, L.; Taddei, R.; Moreno, A.; Gelmetti, D.; Barbieri, I.; De Marco, M.A.; Tosi, G.; Cordioli, P.; Massi, P. Influenza A pandemic (H1N1) 2009 virus outbreak in a cat colony in Italy. Zoonoses Public Health 2011, 58, 573–581. [Google Scholar] [CrossRef]
- Jeoung, H.Y.; Lim, S.I.; Shin, B.H.; Lim, J.-A.; Song, J.-Y.; Song, D.-S.; Kang, B.-K.; Moon, H.-J.; An, D.-J. A novel canine influenza H3N2 virus isolated from cats in an animal shelter. Vet. Microbiol. 2013, 165, 281–286. [Google Scholar] [CrossRef]
- Paniker, C.K.; Nair, C.M. Infection with A2 Hong Kong influenza virus in domestic cats. Bull. World Health Organ. 1970, 43, 859–862. [Google Scholar]
- Romváry, J.; Rózsa, J.; Farkas, E. Infection of dogs and cats with the Hong Kong influenza A (H3N2) virus during an epidemic period in Hungary. Acta Vet. Acad. Sci. Hung. 1975, 25, 255–259. [Google Scholar]
- Hinshaw, V.S.; Webster, R.G.; Easterday, B.C.; Bean, W.J., Jr. Replication of avian influenza A viruses in mammals. Infect. Immun. 1981, 34, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Kuiken, T.; Rimmelzwaan, G.; van Riel, D.; Van Amerongen, G.; Baars, M.; Fouchier, R.; Osterhaus, A. Avian H5N1 influenza in cats. Science 2004, 306, 241. [Google Scholar] [CrossRef] [PubMed]
- Rimmelzwaan, G.F.; van Riel, D.; Baars, M.; Bestebroer, T.M.; van Amerongen, G.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Influenza A virus (H5N1) infection in cats causes systemic disease with potential novel routes of virus spread within and between hosts. Am. J. Pathol. 2006, 168, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Van Riel, D.; Munster, V.J.; de Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.; Osterhaus, A.D.M.E.; Kuiken, T. H5N1 virus attachment to lower respiratory tract. Science 2006, 312, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Songserm, T.; Amonsin, A.; Jam-on, R.; Sae-Heng, N.; Meemak, N.; Pariyothorn, N.; Payungporn, S.; Theamboonlers, A.; Poovorawan, Y. Avian influenza H5N1 in naturally infected domestic cat. Emerg. Infect. Dis. 2006, 12, 681–683. [Google Scholar] [CrossRef] [PubMed]
- Yingst, S.L.; Saad, M.D.; Felt, S.A. Qinghai-like H5N1 from domestic cats, northern Iraq. Emerg. Infect. Dis. 2006, 12, 1295–1297. [Google Scholar] [CrossRef]
- Keawcharoen, J.; Oraveerakul, K.; Kuiken, T.; Fouchier, R.A.M.; Amonsin, A.; Payungporn, S.; Noppornpanth, S.; Wattanodorn, S.; Theambooniers, A.; Tantilertcharoen, R.; et al. Avian influenza H5N1 in tigers and leopards. Emerg. Infect. Dis. 2004, 10, 2189–2191. [Google Scholar] [CrossRef]
- Reperant, L.A.; van de Bildt, M.W.G.; van Amerongen, G.; Leijten, L.M.E.; Watson, S.; Palser, A.; Kellam, P.; Eissens, A.C.; Frijlink, H.W.; Osterhaus, A.D.M.E.; et al. Marked endotheliotropism of highly pathogenic avian influenza virus H5N1 following intestinal inoculation in cats. J. Virol. 2012, 86, 1158–1165. [Google Scholar] [CrossRef] [Green Version]
- Daly, J.M.; Blunden, A.S.; Macrae, S.; Miller, J.; Bowman, S.J.; Kolodziejek, J.; Nowotny, N.; Smith, K.C. Transmission of equine influenza virus to English foxhounds. Emerg. Infect. Dis. 2008, 14, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.C.; Bromfield, C.R.; Crawford, P.C.; Dodds, W.J.; Gibbs, E.P.; Hernandez, J.A. Serological evidence of H3N8 canine influenza-like virus circulation in USA dogs prior to 2004. Vet. J. 2012, 191, 312–316. [Google Scholar] [CrossRef]
- Payungporn, S.; Crawford, P.C.; Kouo, T.S.; Chen, L.M.; Pompey, J.; Castleman, W.L.; Dubovi, E.J.; Katz, J.M.; Donis, R.O. Influenza A virus (H3N8) in dogs with respiratory disease, Florida. Emerg. Infect. Dis. 2008, 14, 902–908. [Google Scholar] [CrossRef] [PubMed]
- Rivailler, P.; Perry, I.A.; Jang, Y.; Davis, C.T.; Chen, L.M.; Dubovi, E.J.; Donis, R.O. Evolution of canine and equine influenza (H3N8) viruses co-circulating between 2005 and 2008. Virology 2010, 408, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.; Wang, L.; Fu, X.; He, S.; Hong, M.; Zhou, P.; Lai, A.; Gray, G.; Li, S. Equine influenza A(H3N8) virus infection in cats. Emerg. Infect. Dis. 2014, 20, 2096–2099. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Hughes, J.; Murcia, P.R. Origins and evolutionary dynamics of H3N2 canine influenza virus. J. Virol. 2015, 89, 5406–5418. [Google Scholar] [CrossRef] [Green Version]
- Voorhees, I.E.H.; Glaser, A.L.; Toohey-Kurth, K.L.; Newbury, S.; Dalziel, B.D.; Dubovi, E.J.; Poulsen, K.; Leutenegger, C.; Willgert, K.J.; Brisbane-Cohen, L.; et al. Spread of canine influenza A (H3N2) virus, United States. Emerg. Infect. Dis. 2017, 23, 1950–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Song, D.; Moon, H.; Yeom, M.; Park, S.; Hong, M.; Na, W.; Webby, R.J.; Webster, R.G.; Park, B.; et al. Inter- and intraspecies transmission of canine influenza virus (H3N2) in dogs, cats, and ferrets. Influenza Other Respir. Viruses 2013, 7, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Blachere, F.M.; Lindsley, W.G.; Weber, A.M.; Beezhold, D.H.; Thewlis, R.E.; Mead, K.R.; Noti, J.D. Detection of an avian lineage influenza A(H7N2) virus in air and surface samples at a New York City feline quarantine facility. Influenza Other Respir. Viruses 2018, 12, 613–622. [Google Scholar] [CrossRef]
- Lee, C.T.; Slavinski, S.; Schiff, C.; Merlino, M.; Daskalakis, D.; Liu, D.; Rakeman, J.L.; Misener, M.; Thompson, C.; Leung, Y.L.; et al. Outbreak of Influenza A(H7N2) Among Cats in an Animal Shelter with Cat-to-Human Transmission-New York City, 2016. Clin. Infect. Dis. 2017, 65, 1927–1929. [Google Scholar] [CrossRef]
- Hatta, M.; Zhong, G.; Gao, Y.; Nakajima, N.; Fan, S.; Chiba, S.; Deering, K.M.; Ito, M.; Imai, M.; Kiso, M.; et al. Characterization of a feline influenza A(H7N2) virus. Emerg. Infect. Dis. 2018, 24, 75–86. [Google Scholar] [CrossRef]
- Belser, J.A.; Pulit-Penaloza, J.A.; Sun, X.; Brock, N.; Pappas, C.; Creager, H.; Zeng, H.; Tumpey, T.M.; Maines, T.R. A novel A(H7N2) influenza virus isolated from a veterinarian caring for cats in a New York City animal shelter causes mild disease and transmits poorly in the ferret model. J. Virol. 2017, 91, e00672-17. [Google Scholar] [CrossRef] [Green Version]
- Pigott, A.M.; Haak, C.E.; Breshears, M.A.; Linklater, A.K. Acute bronchointerstitial pneumonia in two indoor cats exposed to the H1N1 influenza virus. J. Vet. Emerg. Crit. Care 2014, 24, 715–723. [Google Scholar] [CrossRef]
- Löhr, C.V.; DeBess, E.E.; Baker, R.J.; Hiett, S.L.; Hoffman, K.A.; Murdoch, V.J.; Fischer, K.A.; Mulrooney, D.M.; Selman, R.L.; Hammill-Black, W.M. Pathology and viral antigen distribution of lethal pneumonia in domestic cats due to pandemic (H1N1) 2009 influenza A virus. Vet. Pathol. 2010, 47, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Sponseller, B.A.; Strait, E.; Jergens, A.; Trujillo, J.; Harmon, K.; Koster, L.; Jenkins-Moore, M.; Killian, M.; Swenson, S.; Bender, H.; et al. Influenza A pandemic (H1N1) 2009 virus infection in domestic cat. Emerg. Infect. Dis. 2010, 16, 534–537. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Daniels, J.B.; Zhang, Y.; Rodriguez-Palacios, A.; Hayes-Ozello, K.; Mathes, L.; Lee, C.-W. Pandemic and seasonal human influenza virus infections in domestic cats: Prevalence, association with respiratory disease, and seasonality patterns. J. Clin. Microbiol. 2011, 49, 4101–4105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campagnolo, E.R.; Rankin, J.T.; Daverio, S.A.; Hunt, E.A.; Lute, J.R.; Tewari, D.; Acland, H.M.; Ostrowski, S.R.; Moll, M.E.; Urdaneta, V.V.; et al. Fatal pandemic (H1N1) 2009 influenza A virus infection in a Pennsylvania domestic cat. Zoonoses Public Health 2011, 58, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Tangwangvivat, R.; Chanvatik, S.; Charoenkul, K.; Chaiyawong, S.; Janethanakit, T.; Tuanudom, R.; Prakairungnamthip, D.; Boonyapisitsopa, S.; Bunpapong, N.; Amonsin, A. Evidence of pandemic H1N1 influenza exposure in dogs and cats, Thailand: A serological survey. Zoonoses Public Health 2019, 66, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.R.; Liu, C.G.; Yin, X.; Zhou, D.H.; Wei, P.; Chang, H.Y. Serological report of pandemic (H1N1) 2009 infection among cats in northeastern China in 2012-02 and 2013-03. Virol. J. 2014, 11, 49. [Google Scholar] [CrossRef] [Green Version]
- Van den Brand, J.M.; Stittelaar, K.J.; van Amerongen, G.; Van De Bildt, M.W.; Leijten, L.M.; Kuiken, T.; Osterhaus, A.D. Experimental pandemic (H1N1) 2009 virus infection of cats. Emerg. Infect. Dis. 2010, 16, 1745–1747. [Google Scholar] [CrossRef]
- Cao, X.; Yang, F.; Wu, H.; Xu, L. Genetic characterization of novel reassortant H5N6-subtype influenza viruses isolated from cats in eastern China. Arch. Virol. 2017, 162, 3501–3505. [Google Scholar] [CrossRef] [PubMed]
- Borland, S.; Gracieux, P.; Jones, M.; Mallet, F.; Yugueros-Marcos, J. Influenza A virus infection in cats and dogs: A literature review in the light of the “One Health” concept. Front. Public Health 2020, 8. [Google Scholar] [CrossRef]
- Webster, R.G.; Guan, Y.; Poon, L.; Krauss, S.; Webby, R.; Govorkovai, E.; Peiris, M. The spread of the H5N1 bird flu epidemic in Asia in 2004. Arch. Virol. Suppl. 2005, 19, 17–29. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cumulative Number of Confirmed Human Cases for Avian Influenza A(H5N1). Available online: https://www.who.int/influenza/human_animal_interface/2020_10_07_tableH5N1.pdf?ua=1) (accessed on 16 June 2021).
- Thanawongnuwech, R.; Amonsin, A.; Tantilertcharoen, R.; Damrongwatanapokin, S.; Theamboonlers, A.; Payungporn, S.; Nanthapornphiphat, K.; Ratanamungklanon, S.; Tunak, E.; Songserm, T.; et al. Probable tiger-to-tiger transmission of avian influenza H5N1. Emerg. Infect. Dis. 2005, 11, 699–701. [Google Scholar] [CrossRef]
- Leschnik, M.; Weikel, J.; Möstl, K.; Revilla-Fernández, S.; Wodak, E.; Bagó, Z.; Vanek, E.; Benetka, V.; Hess, M.; Thalhammer, J.G. Subclinical infection with avian influenza A (H5N1) virus in cats. Emerg. Infect. Dis. 2007, 13, 243–247. [Google Scholar] [CrossRef]
- Marschall, J.; Schulz, B.; Harder, T.C.; Vahlenkamp, T.W.; Huebner, J.; Huisinga, E.; Hartmann, K. Prevalence of influenza A H5N1 virus in cats from areas with occurrence of highly pathogenic avian influenza in birds. J. Feline Med. Surg. 2008, 10, 355–358. [Google Scholar] [CrossRef]
- Zhao, F.R.; Zhou, D.H.; Zhang, Y.G.; Shao, J.-J.; Lin, T.; Li, Y.-F.; Wei, P.; Chang, H.-Y. Detection prevalence of H5N1 avian influenza virus among stray cats in eastern China. J. Med. Virol. 2015, 87, 436–1440. [Google Scholar] [CrossRef]
- Vahlenkamp, T.W.; Harder, T.C.; Giese, M.; Lin, F.; Teifke, J.P.; Klopfleisch, R.; Hoffmann, R.; Tarpey, I.; Beer, M.; Mettenleiter, T.C. Protection of cats against lethal influenza H5N1 challenge infection. J. Gen. Virol. 2008, 89, 968–974. [Google Scholar] [CrossRef]
- ProMED-mail. PRO/AH/EDR> Avian Influenza (82): Europe (Germany, Estonia) Wild Bird, HPAI H5N1, OIE. Available online: https://promedmail.org/promed-post/?id=8446706 (accessed on 16 June 2021).
- Knight, C.G.; Davies, J.L.; Joseph, T.; Ondrich, S.; Rosa, B.V. Pandemic H1N1 influenza virus infection in a Canadian cat. Can. Vet. J. 2016, 57, 497–500. [Google Scholar]
- Statement of the Standing Committee on the Food Chain and Animal Health. Available online: https://ec.europa.eu/commission/presscorner/detail/en/MEMO_06_104 (accessed on 13 June 2021).
- Velineni, S.; Hainer, N.; Conlee, D.; Hutchinson, K. Vaccination with an inactivated canine influenza H3N2 virus vaccine is safe and elicits an immune response in cats. J. Feline Med. Surg. 2020, 22, 199–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harder, T.C.; Vahlenkamp, T.W. Influenza virus infections in dogs and cats. Vet. Immunol. Immunopathol. 2010, 134, 54–60. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frymus, T.; Belák, S.; Egberink, H.; Hofmann-Lehmann, R.; Marsilio, F.; Addie, D.D.; Boucraut-Baralon, C.; Hartmann, K.; Lloret, A.; Lutz, H.; et al. Influenza Virus Infections in Cats. Viruses 2021, 13, 1435. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081435
Frymus T, Belák S, Egberink H, Hofmann-Lehmann R, Marsilio F, Addie DD, Boucraut-Baralon C, Hartmann K, Lloret A, Lutz H, et al. Influenza Virus Infections in Cats. Viruses. 2021; 13(8):1435. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081435
Chicago/Turabian StyleFrymus, Tadeusz, Sándor Belák, Herman Egberink, Regina Hofmann-Lehmann, Fulvio Marsilio, Diane D. Addie, Corine Boucraut-Baralon, Katrin Hartmann, Albert Lloret, Hans Lutz, and et al. 2021. "Influenza Virus Infections in Cats" Viruses 13, no. 8: 1435. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081435