
Virucidal Efficacy of Blue LED and Far-UVC Light Disinfection against Feline Infectious Peritonitis Virus as a Model for SARS-CoV-2



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Preparation of Virus Stock
2.3. Virus Titration
2.4. Virus Contamination and Recovery Matrices
2.5. Light Sources
2.6. Temperature and Humidity Readings
2.7. Light Exposure Trials
2.7.1. Effects of Light Exposure on Viability of Virus in Suspension
2.7.2. Effects of Light Treatment on Viability of Virus on Different Surface Matrices
2.8. Data and Statistical Analysis
3. Results
3.1. Inactivation of FIPV after Exposure to UVC LED and UVC Mercury Lights
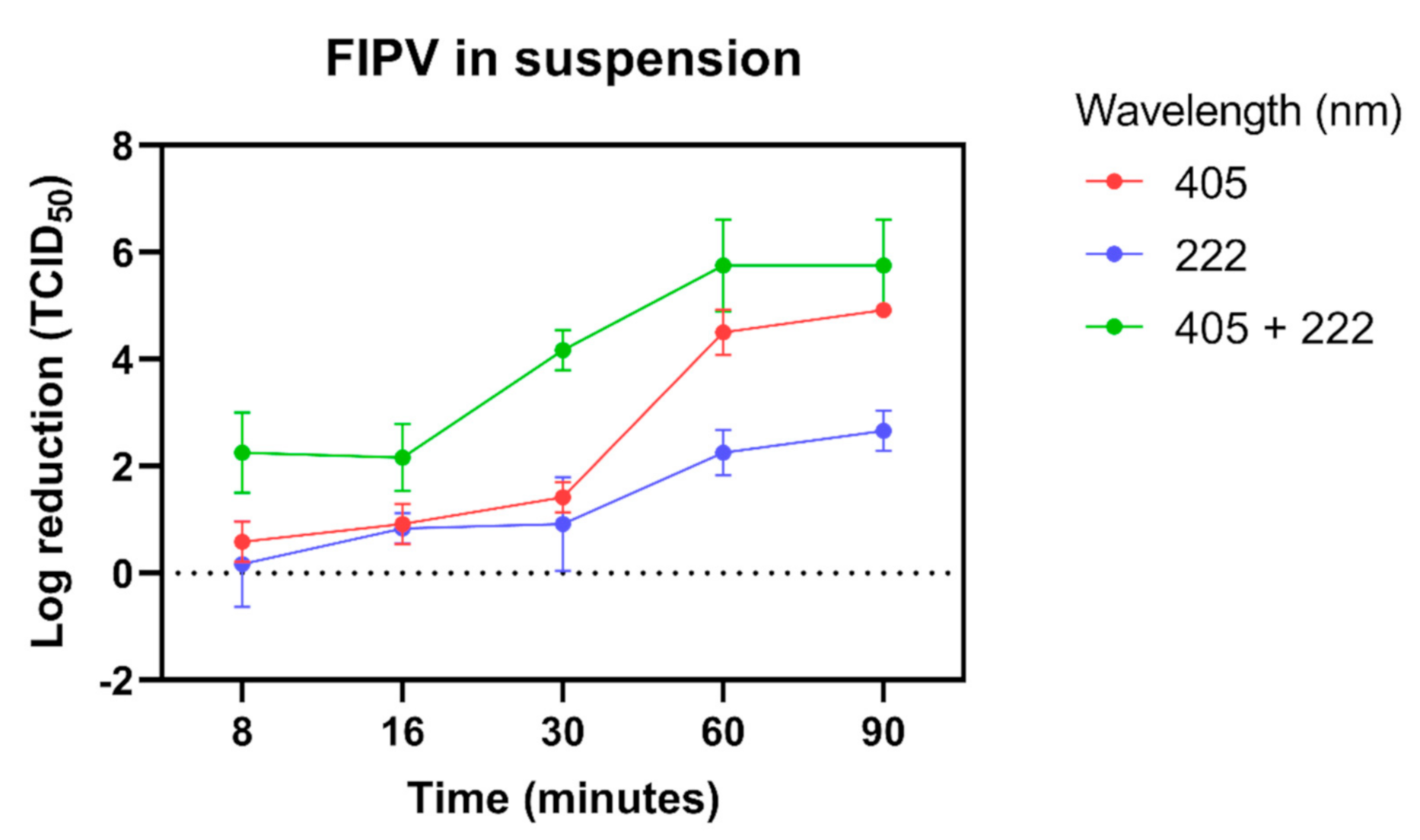
3.2. Inactivation of FIPV in Liquid Suspension after Exposure to Light at 405 nm and 222 nm
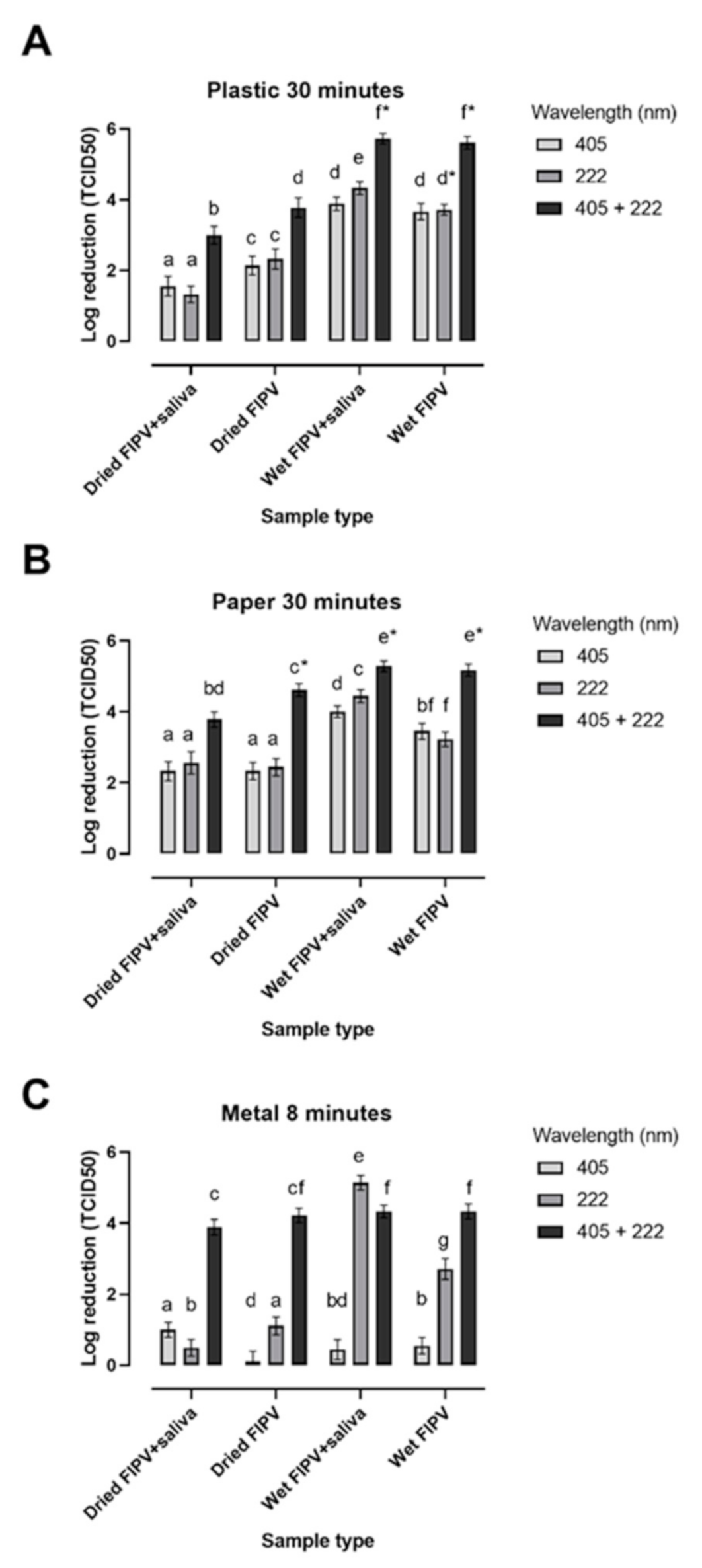
3.3. Inactivation of FIPV on Different Matrices after Exposure to Lights at 405 nm and 222 nm
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Origins of SARS-CoV-2. Available online: https://apps.who.int/iris/bitstream/handle/10665/332197/WHO-2019-nCoV-FAQ-Virus_origin-2020.1-eng.pdf (accessed on 16 June 2021).
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- World Health Organization. Worldometer COVID-19 Coronavirus Pandemic. Available online: https://www.worldometers.info/coronavirus/ (accessed on 28 May 2021).
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar]
- Wang, Y.; Grunewald, M.; Perlman, S. Coronaviruses: An Updated Overview of Their Replication and Pathogenesis. In Coronaviruses. Methods in Molecular Biology; Maier, H., Bickerton, E., Eds.; Springer: New York, NY, USA, 2020; Volume 2203. [Google Scholar]
- Pedersen, N.C. A review of feline infectious peritonitis virus infection: 1963–2008. J. Feline Med. Surg. 2009, 11, 225–258. [Google Scholar] [CrossRef]
- Pedersen, N.C. An update on feline infectious peritonitis: Virology and immunopathogenesis. Vet. J. 2014, 201, 123–132. [Google Scholar] [CrossRef] [Green Version]
- CDC. Coronaviruses. Available online: https://www.cdc.gov/coronavirus/types.html (accessed on 17 May 2021).
- World Health Organization. Coronavirus. Available online: https://www.who.int/health-topics/coronavirus#tab=tab_1 (accessed on 17 May 2021).
- Drosten, C.; Günther, S.; Preiser, W.; van der Werf, S.; Brodt, H.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A.M.; et al. Identification of a Novel Coronavirus in Patients with Severe Acute Respiratory Syndrome. N. Engl. J. Med. 2003, 348, 1967–1976. [Google Scholar] [CrossRef]
- Ksiazek, T.G.; Erdman, D.; Goldsmith, C.S.; Zaki, S.R.; Peret, T.; Emery, S.; Tong, S.; Urbani, C.; Comer, J.A.; Lim, W.; et al. A Novel Coronavirus Associated with Severe Acute Respiratory Syndrome. N. Engl. J. Med. 2003, 348, 1953–1966. [Google Scholar] [CrossRef]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Sreedharan, J.; Nair, S.C.; Muttappallymyalil, J.; Gopakumar, A.; Eapen, N.T.; Satish, K.P.; Manda, V. Case fatality rates of COVID-19 across the globe: Are the current draconian measures justified? J. Public Health 2021. [Google Scholar] [CrossRef]
- World Health Organization. WHO MERS-CoV Global Summary and Risk Assessment; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- World Health Organization. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/interactive-timeline# (accessed on 17 May 2021).
- Lin, X.D.; Wang, W.; Hao, Z.; Wang, Z.; Guo, W.; Guan, X.; Wang, M.; Wang, H.; Zhou, R.; Li, M.; et al. Extensive diversity of coronaviruses in bats from China. Virology 2017, 507, 1–10. [Google Scholar] [CrossRef]
- Guan, Y.; Zheng, B.J.; He, Y.Q.; Liu, X.L.; Zhuang, Z.X.; Cheung, C.L.; Luo, S.W.; Li, P.H.; Zhang, L.J.; Guan, Y.J.; et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drosten, C.; Kellam, P.; Memish, Z.A. Evidence for camel-to-human transmission of MERS coronavirus. N. Engl. J. Med. 2014, 371, 1359–1360. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.; Wang, X.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.; Zhu, Y.; Li, B.; Huang, C.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassell, J.M.; Begon, M.; Ward, M.J.; Fèvre, E.M. Urbanization and Disease Emergence: Dynamics at the Wildlife–Livestock–Human Interface. Trends Ecol. Evol. 2017, 32, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.F.; Yuan, S.; Kok, K.; To, K.K.; Chu, H.; Yang, J.; Xing, F.; Liu, J.; Yip, C.C.; Poon, R.W.; et al. A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: A study of a family cluster. Lancet 2020, 395, 514–523. [Google Scholar] [CrossRef] [Green Version]
- Bahl, P.; Doolan, C.; de Silva, C.; Chughtai, A.A.; Bourouiba, L.; MacIntyre, C.R. Airborne or droplet precautions for health workers treating COVID-19? J. Infect. Dis. 2020, jiaa189. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Xu, Y.; Gao, R.; Lu, R.; Han, K.; Wu, G.; Tan, W. Detection of SARS-CoV-2 in Different Types of Clinical Specimens. J. Am. Med Assoc. 2020, 323, 1843–1844. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Wang, Z.; Zhang, S.; Li, X.; Li, L.; Li, C.; Cui, Y.; Fu, R.; Dong, Y.; Chi, X.; et al. Aerosol and Surface Distribution of Severe Acute Respiratory Syndrome Coronavirus 2 in Hospital Wards, Wuhan, China, 2020. Emerg. Infect. Dis. 2020, 26, 1586. [Google Scholar] [CrossRef]
- Pastorino, B.; Touret, F.; Gilles, M.; de Lamballerie, X.; Charrel, R.N. Prolonged Infectivity of SARS-CoV-2 in Fomites. Emerg. Infect. Dis. 2020, 26, 2256. [Google Scholar] [CrossRef] [PubMed]
- Van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef]
- Food Standards Agency (UK). Qualitative Risk Assessment: What Is the Risk of Food or Food Contact Materials Being a Source or Transmission Route of SARS-CoV-2 for UK Consumers? Available online: https://www.food.gov.uk/research/research-projects/qualitative-risk-assessment-on-the-risk-of-food-or-food-contact-materials-as-a-transmission-route-for-sars-cov-2 (accessed on 28 May 2021).
- French Agency for Food, Environmental and Occupational Health (ANSES). Opinion on an Urgent Request to Assess Certain Risks Associated with COVID-19. Available online: https://www.anses.fr/en/system/files/SABA2020SA0037-1.pdf (accessed on 28 May 2021).
- Xiling, G.; Yin, C.; Ling, W.; Xiaosong, W.; Jingjing, F.; Fang, L.; Xiaoyan, Z.; Yiyue, G.; Ying, C.; Lunbiao, C.; et al. In vitro inactivation of SARS-CoV-2 by commonly used disinfection products and methods. Sci. Rep. 2021, 11, 2418. [Google Scholar] [CrossRef]
- Hadi, J.; Dunowska, M.; Wu, S.; Brightwell, G. Control Measures for SARS-CoV-2: A Review on Light-Based Inactivation of Single-Stranded RNA Viruses. Pathogens 2020, 9, 737. [Google Scholar] [CrossRef] [PubMed]
- Soehnge, H.; Ouhtit, A.; Ananthaswamy, O.N. Mechanisms of induction of skin cancer by UV radiation. Front. Biosci. 1997, 2, 538–551. [Google Scholar]
- Cutler, T.D.; Zimmerman, J.J. Ultraviolet irradiation and the mechanisms underlying its inactivation of infectious agents. Anim. Health Res. Rev. 2011, 12, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Zaffina, S.; Camisa, V.; Lembo, M.; Vinci, M.R.; Tucci, M.G.; Borra, M.; Napolitano, A.; Cannatà, V. Accidental Exposure to UV Radiation Produced by Germicidal Lamp: Case Report and Risk Assessment. Photochem. Photobiol. 2012, 88, 1001–1004. [Google Scholar] [CrossRef]
- Narita, K.; Asano, K.; Naito, K.; Ohashi, H.; Sasaki, M.; Morimoto, Y.; Igarashi, T.; Nakane, A. Ultraviolet C light with wavelength of 222 nm inactivates a wide spectrum of microbial pathogens. J. Hosp. Infect. 2020, 105, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Tomb, R.M.; Maclean, M.; Coia, J.E.; Graham, E.; McDonald, M.; Atreya, C.D.; MacGregor, S.J.; Anderson, J.G. New Proof-of-concept in viral inactivation: Virucidal Efficacy of 405 nm light against feline Calicivirus as a model for Norovirus Decontamination. Food Environ. Virol. 2017, 9, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Araud, E.; Fuzawa, M.; Shisler, J.L.; Li, J.; Nguyen, T.H. UV inactivation of rotavirus and tulane virus targets different components of the virions. Appl. Environ. Microbiol. 2020, 86, e02436-19. [Google Scholar] [CrossRef]
- Narita, K.; Asano, K.; Morimoto, Y.; Igarashi, T.; Nakane, A. Chronic irradiation with 222-nm UVC light induces neither DNA damage nor epidermal lesions in mouse skin, even at high doses. PLoS ONE 2018, 13, e0201259. [Google Scholar] [CrossRef]
- Wainwright, M. Photoinactivation of viruses. Photochem. Photobiol. Sci. 2004, 3, 406–411. [Google Scholar] [CrossRef]
- Freshney, R. Culture of Animal Cells: A Manual of Basic Technique, 5th ed.; Wiley: Hoboken, NJ, USA, 2005. [Google Scholar]
- Hsiung, G.D.; Fong, C.K.Y.; Landry, M.L. Hsiung’s Diagnostic Virology, 4th ed.; Yale University Press: New Haven, CT, USA, 1994. [Google Scholar]
- Atkinson, G.F. The Spearman-Karber Method of Estimating 50% Endpoints; Cornell University: Ithaca, NY, USA, 1961. [Google Scholar]
- University Hospital Heidelberg, TCID50 Calculator. Available online: https://www.klinikum.uni-heidelberg.de/zentrum-fuer-infektiologie/molecular-virology/welcome/downloads (accessed on 23 July 2021).
- Kaufer, A.M.; Theis, T.; Lau, K.A.; Gray, J.L.; Rawlinson, W.D. Laboratory biosafety measures involving SARS-CoV-2 and the classification as a Risk Group 3 biological agent. Pathology 2020, 52, 790–795. [Google Scholar] [CrossRef] [PubMed]
- FDA. Q5A Viral Safety Evaluation of Biotechnology Products Derived from Cell Lines of Human or Animal Origin. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/q5a-viral-safety-evaluation-biotechnology-products-derived-cell-lines-human-or-animal-origin (accessed on 6 June 2021).
- Anonymous. Virus Taxonomy: The Classification and Nomenclature of Viruses The Online (10th) Report of the ICTV; International Committee on Taxonomy of Viruses (ICTV): London, UK, 2017. [Google Scholar]
- Saknimit, M.; Inatsuki, I.; Sugiyama, Y.; Yagami, K. Virucidal efficacy of physico-chemical treatments against coronaviruses and parvoviruses of laboratory animals. Jikken Dobutsu. Exp. Anim. 1988, 37, 341–345. [Google Scholar]
- Theerawatanasirikul, S.; Kuo, C.J.; Phecharat, N.; Chootip, J.; Lekcharoensuk, C.; Lekcharoensuk, P. Structural-based virtual screening and in vitro assays for small molecules inhibiting the feline coronavirus 3CL protease as a surrogate platform for coronaviruses. Antivir. Res. 2020, 182, 104927. [Google Scholar] [CrossRef] [PubMed]
- Camero, M.; Lanave, G.; Catella, C.; Lucente, M.S.; Decaro, N.; Martella, V.; Buonavoglia, C. Evaluation of virucidal activity of fabrics using feline coronavirus. J. Virol. Methods 2021, 295, 114214. [Google Scholar] [CrossRef] [PubMed]
- Cimolai, N. Environmental and decontamination issues for human coronaviruses and their potential surrogates. J. Med. Virol. 2020, 92, 2498–2510. [Google Scholar] [CrossRef] [PubMed]
- Darnell, M.E.; Subbarao, K.; Feinstone, S.M.; Taylor, D.R. Inactivation of the coronavirus that induces severe acute respiratory syndrome, SARS-CoV. J. Virol. Methods 2004, 121, 85–91. [Google Scholar] [CrossRef]
- Darnell, M.E.; Taylor, D.R. Evaluation of inactivation methods for severe acute respiratory syndrome coronavirus in noncellular blood products. Transfusion 2006, 46, 1770–1777. [Google Scholar] [CrossRef] [Green Version]
- Duan, S.M.; Zhao, X.S.; Wen, R.F.; Huang, J.J.; Pi, G.H.; Zhang, S.X.; Han, J.; Bi, S.L.; Ruan, L.; Dong, X.P.; et al. Stability of SARS coronavirus in human specimens and environment and its sensitivity to heating and UV irradiation. Biomed. Environ. Sci. 2003, 16, 246–255. [Google Scholar]
- Mukhopadhyay, S.; Ukuku, D.O.; Juneja, V.; Fan, X. Effects of UV-C treatment on inactivation of Salmonella enterica and Escherichia coli O157:H7 on grape tomato surface and stem scars, microbial loads, and quality. Food Control 2014, 44, 110–117. [Google Scholar] [CrossRef]
- Kim, Y.H.; Jeong, S.G.; Back, K.H.; Park, K.H.; Chung, M.S.; Kang, D.H. Effect of various conditions on inactivation of Escherichia coli O157:H7, Salmonella Typhimurium, and Listeria monocytogenes in fresh-cut lettuce using ultraviolet radiation. Int. J. Food Microbiol. 2013, 166, 349–355. [Google Scholar] [CrossRef]
- Casini, B.; Tuvo, B.; Cristina, M.L.; Spagnolo, A.M.; Totaro, M.; Baggiani, A.; Privitera, G.P. Evaluation of an Ultraviolet C (UVC) Light-Emitting Device for Disinfection of High Touch Surfaces in Hospital Critical Areas. Int. J. Environ. Res. Public Health 2019, 16, 3572. [Google Scholar] [CrossRef] [Green Version]
- Bolton, J.R.; Mayor-Smith, I.; Linden, K.G. Rethinking the concepts of fluence (UV dose) and fluence rate: The importance of photon-based units—A systemic review. Photochem. Photobiol. 2015, 91, 1252–1262. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Shao, S.; Li, J.; Kumar, S.S.; Sokoloff, J.B.; Hong, J. Droplet evaporation residue indicating SARS-COV-2 survivability on surfaces. Phys. Fluids 2021, 33, 013309. [Google Scholar] [CrossRef] [PubMed]
- Marcén, M.; Ruiz, V.; Serrano, J.; Condón, S.; Mañas, P. Oxidative stress in E. coli cells upon exposure to heat treatments. Int. J. Food Microbiol. 2017, 16, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Ratnesar-Shumate, S.; Williams, G.; Green, B.; Krause, M.; Holland, B.; Wood, S.; Bohannon, J.; Boydston, J.; Freeburger, D.; Hooper, I.; et al. Simulated Sunlight Rapidly Inactivates SARS-CoV-2 on Surfaces. J. Infect. Dis. 2020, 222, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Goldfarb, A.R.; Saidel, L.J.; Mosovich, E. Ultraviolet absorption spectra of proteins. J. Biol. Chem. 1951, 193, 397–404. [Google Scholar] [CrossRef]
- Coohill, T.P. Virus-cell interactions as probes for vacuum-ultraviolet radiation damage and repair. Photochem. Photobiol. 1986, 44, 359–363. [Google Scholar] [CrossRef]
- Ash, C.; Dubec, M.; Donne, K.; Bashford, T. Effect of wavelength and beam width on penetration in light-tissue interaction using computational methods. Lasers Med. Sci. 2017, 32, 1909–1918. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Gupta, A.; Murray, C.K.; Vrahas, M.S.; Tegos, G.P.; Hamblin, M.R. Blue light for infectious diseases: Propionibacterium acnes, Helicobacter pylori, and beyond? Drug Resist. Updates 2012, 15, 223–236. [Google Scholar] [CrossRef] [Green Version]
- Bumah, V.V.; Aboualizadeh, E.; Masson-Meyers, D.S.; Eells, J.T.; Enwemeka, C.S.; Hirschmugl, C.J. Spectrally resolved infrared microscopy and chemometric tools to reveal the interaction between blue light (470 nm) and methicillin-resistant Staphylococcus aureus. J. Photochem. Photobiol. 2017, 167, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Sizunf, J.; Yu, M.W.N.; Talbot, P.J. Survival of human coronaviruses 229E and OC43 in suspension and after drying onsurfaces: A possible source ofhospital-acquired infections. J. Hosp. Infect. 2000, 46, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Gélinas, P.; Goulet, J. Neutralization of the activity of eight disinfectants by organic matter. J. Appl. Bacteriol. 1983, 54, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.L.; Boehm, A.B.; Davies-Colley, R.J.; Dodd, M.C.; Kohn, T.; Linden, K.G.; Liu, Y.; Maraccini, P.A.; McNeill, K.; Mitch, W.A.; et al. Sunlight-mediated inactivation of health-relevant microorganisms in water: A review of mechanisms and modeling approaches. Environ. Sci. Process. Impacts 2018, 20, 1089–1122. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Matrix/Virus | Saliva | Time (min) | Dose (J/cm2) | Control Virus (TCID50/mL ± SD) | Treated Virus (TCID50/mL ± SD) | Log Reduction ± SD |
|---|---|---|---|---|---|---|
| 265 nm + 405 nm | ||||||
| Media | − | 1 | 1.350 | 5.13 ± 0.17 | 0.30 ± 0.00 | 4.83 ± 0.17 1 |
| Metal | ||||||
| Wet | + | 1 | 1.350 | 5.58 ± 0.20 | 0.30 ± 0.00 | 5.28 ± 0.20 1 |
| Wet | − | 1 | 1.350 | 5.80 ± 0.21 | 0.91 ± 0.11 | 4.89 ± 0.24 |
| Dried | + | 1 | 1.350 | 5.25 ± 0.18 | 0.30 ± 0.00 | 4.94 ± 0.18 1 |
| Dried | − | 1 | 1.350 | 5.91 ± 0.18 | 1.25 ± 0.18 | 4.67 ± 0.25 |
| Paper | ||||||
| Wet | + | 1 | 1.350 | 5.58 ± 0.20 | 0.30 ± 0.00 | 5.28 ± 0.20 1 |
| Wet | − | 1 | 1.350 | 6.13 ± 0.17 | 0.80 ± 0.00 | 5.33 ± 0.17 2 |
| Dried | + | 1 | 1.350 | 5.13 ± 0.17 | 0.30 ± 0.00 | 4.83 ± 0.17 1 |
| Dried | − | 1 | 1.350 | 5.02 ± 0.15 | 0.30 ± 0.00 | 4.72 ± 0.15 1 |
| Plastic | ||||||
| Wet | + | 1 | 1.350 | 5.58 ± 0.15 | 0.30 ± 0.00 | 5.28 ± 0.15 1 |
| Wet | − | 1 | 1.350 | 5.25 ± 0.18 | 0.30 ± 0.00 | 4.94 ± 0.18 1 |
| Dried | + | 1 | 1.350 | 3.91 ± 0.11 | 0.30 ± 0.00 | 3.61 ± 0.11 1 |
| Dried | − | 1 | 1.350 | 4.80 ± 0.16 | 0.80 ± 0.00 | 4.00 ± 0.16 2 |
| 265 nm | ||||||
| Media | − | 1 | 0.101 | 5.25 ± 0.18 | 0.30 ± 0.00 | 4.94 ± 0.18 1 |
| Metal | ||||||
| Wet | + | 1 | 0.101 | 5.69 ± 0.11 | 0.30 ± 0.00 | 5.39 ± 0.11 1 |
| Wet | − | 1 | 0.101 | 4.91 ± 0.18 | 0.30 ± 0.00 | 4.61 ± 0.18 1 |
| Dried | + | 1 | 0.101 | 3.25 ± 0.20 | 0.30 ± 0.00 | 2.94 ± 0.20 1 |
| Dried | − | 1 | 0.101 | 3.91 ± 0.11 | 0.30 ± 0.00 | 3.61 ± 0.11 1 |
| Paper | ||||||
| Wet | + | 1 | 0.101 | 5.69 ± 0.18 | 0.30 ± 0.00 | 5.39 ± 0.18 1 |
| Wet | − | 1 | 0.101 | 4.58 ± 0.20 | 0.30 ± 0.00 | 4.28 ± 0.20 1 |
| Dried | + | 1 | 0.101 | 4.80 ± 0.21 | 0.91 ± 0.11 | 3.89 ± 0.24 2 |
| Dried | − | 1 | 0.101 | 5.13 ± 0.21 | 0.30 ± 0.00 | 4.83 ± 0.21 1 |
| Plastic | ||||||
| Wet | + | 1 | 0.101 | 5.91 ± 0.11 | 0.30 ± 0.00 | 5.61 ± 0.11 1 |
| Wet | − | 1 | 0.101 | 4.69 ± 0.18 | 0.30 ± 0.00 | 4.39 ± 0.18 1 |
| Dried | + | 1 | 0.101 | 4.02 ± 0.15 | 0.80 ± 0.00 | 3.22 ± 0.15 2 |
| Dried | − | 1 | 0.101 | 4.91 ± 0.18 | 1.58 ± 0.24 | 3.33 ± 0.17 2 |
| 254 nm | ||||||
| Media | − | 1 | 0.117 | 7.13 ± 0.17 | 0.91 ± 0.11 | 6.22 ± 0.20 |
| Metal | ||||||
| Wet | + | 1 | 0.117 | 5.80 ± 0.21 | 0.30 ± 0.00 | 5.50 ± 0.21 1 |
| Wet | − | 1 | 0.117 | 5.69 ± 0.11 | 0.30 ± 0.00 | 5.39 ± 0.11 1 |
| Dried | + | 1 | 0.117 | 4.02 ± 0.15 | 0.30 ± 0.00 | 3.72 ± 0.15 1 |
| Dried | − | 1 | 0.117 | 4.80 ± 0.00 | 0.30 ± 0.00 | 4.50 ± 0.00 1 |
| Paper | ||||||
| Wet | + | 1 | 0.117 | 5.47 ± 0.21 | 0.30 ± 0.00 | 5.17 ± 0.21 1 |
| Wet | − | 1 | 0.117 | 5.69 ± 0.11 | 0.30 ± 0.00 | 5.39 ± 0.11 1 |
| Dried | + | 1 | 0.117 | 5.02 ± 0.15 | 0.80 ± 0.00 | 4.22 ± 0.15 2 |
| Dried | − | 1 | 0.117 | 4.47 ± 0.17 | 0.30 ± 0.00 | 4.17 ± 0.17 1 |
| Plastic | ||||||
| Wet | + | 1 | 0.117 | 5.25 ± 0.18 | 0.30 ± 0.00 | 4.94 ± 0.18 1 |
| Wet | − | 1 | 0.117 | 5.47 ± 0.17 | 0.30 ± 0.00 | 5.17 ± 0.17 1 |
| Dried | + | 1 | 0.117 | 4.36 ± 0.18 | 0.30 ± 0.00 | 4.05 ± 0.18 1 |
| Dried | − | 1 | 0.117 | 4.13 ± 0.17 | 0.30 ± 0.00 | 3.83 ± 0.17 1 |
| Matrix/Virus | Saliva | Time (min) | Dose (J/cm2) | Control Virus (TCID50/mL ± SD) | Treated Virus (TCID50/mL ± SD) | Log Reduction ± SD |
|---|---|---|---|---|---|---|
| 405 nm + 222 nm | ||||||
| Metal | ||||||
| Wet | + | 4 | 4.11 | 5.02 ± 0.20 | 0.80 ± 0.00 | 4.22 ± 0.20 2 |
| Wet | − | 8 | 8.23 | 5.25 ± 0.18 | 0.91 ± 0.11 | 4.33 ± 0.21 |
| Dried | + | 8 | 8.23 | 4.69 ± 0.22 | 0.80 ± 0.00 | 3.89 ± 0.22 |
| Dried | − | 8 | 8.23 | 5.13 ± 0.17 | 0.91 ± 0.11 | 4.22 ± 0.20 |
| Paper | ||||||
| Wet | + | 8 | 8.23 | 5.80 ± 0.16 | 0.80 ± 0.00 | 5.00 ± 0.16 |
| Wet | − | 16 | 16.46 | 5.69 ± 0.18 | 0.30 ± 0.00 | 5.39 ± 0.22 1 |
| Dried | + | 30 | 30.85 | 4.80 ± 0.16 | 1.02 ± 0.15 | 3.78 ± 0.22 |
| Dried | − | 16 | 16.46 | 4.91 ± 0.22 | 0.30 ± 0.00 | 4.61 ± 0.22 1 |
| Plastic | ||||||
| Wet | + | 16 | 16.46 | 5.91 ± 0.11 | 0.30 ± 0.00 | 5.61 ± 0.11 1 |
| Wet | − | 16 | 16.46 | 5.80 ± 0.16 | 0.30 ± 0.00 | 5.50 ± 0.16 1 |
| Dried | + | 30 | 30.85 | 4.58 ± 0.15 | 1.58 ± 0.20 | 3.00 ± 0.25 |
| Dried | − | 30 | 30.85 | 5.25 ± 0.18 | 1.47 ± 0.21 | 3.78 ± 0.28 |
| 405 nm | ||||||
| Metal | ||||||
| Wet | + | 60 | 60.55 | 5.25 ± 0.18 | 0.30 ± 0.00 | 4.94 ± 0.18 1 |
| Wet | − | 60 | 60.55 | 5.47 ± 0.17 | 0.30 ± 0.00 | 5.17 ± 0.17 1 |
| Dried | + | 90 | 90.83 | 4.91 ± 0.22 | 1.02 ± 0.15 | 3.89 ± 0.27 |
| Dried | − | 60 | 60.55 | 4.47 ± 0.21 | 0.91 ± 0.11 | 3.56 ± 0.24 |
| Paper | ||||||
| Wet | + | 30 | 30.28 | 4.91 ± 0.11 | 0.91 ± 0.11 | 4.00 ± 0.16 |
| Wet | − | 60 | 60.55 | 4.69 ± 0.17 | 0.30 ± 0.00 | 4.39 ± 0.17 1 |
| Dried | + | 30 | 30.28 | 4.91 ± 0.18 | 2.58 ± 0.20 | 2.33 ± 0.27 |
| Dried | − | 30 | 30.28 | 4.91 ± 0.11 | 2.58 ± 0.22 | 2.33 ± 0.25 |
| Plastic | ||||||
| Wet | + | 60 | 60.55 | 4.91 ± 0.11 | 0.80 ± 0.00 | 4.11 ± 0.11 |
| Wet | − | 60 | 60.55 | 4.80 ± 0.00 | 0.30 ± 0.00 | 4.50 ± 0.00 1 |
| Dried | + | 90 | 90.83 | 4.47 ± 0.17 | 1.02 ± 0.15 | 3.45 ± 0.29 |
| Dried | − | 90 | 90.83 | 5.25 ± 0.20 | 2.99 ± 0.19 | 2.26 ± 0.28 |
| 222 nm | ||||||
| Metal | ||||||
| Wet | + | 8 | 0.15 | 5.58 ± 0.20 | 0.80 ± 0.00 | 4.78 ± 0.20 |
| Wet | − | 16 | 0.31 | 5.69 ± 0.18 | 1.13 ± 0.17 | 4.56 ± 0.25 |
| Dried | + | 30 | 0.58 | 4.91 ± 0.18 | 3.25 ± 0.18 | 1.67 ± 0.25 |
| Dried | − | 90 | 1.74 | 5.02 ± 0.15 | 3.80 ± 0.24 | 1.22 ± 0.28 |
| Paper | ||||||
| Wet | + | 16 | 0.31 | 5.58 ± 0.15 | 0.69 ± 0.19 | 4.89 ± 0.24 |
| Wet | − | 60 | 1.16 | 4.36 ± 0.18 | 0.30 ± 0.00 | 4.05 ± 0.18 1 |
| Dried | + | 90 | 1.74 | 4.36 ± 0.18 | 1.02 ± 0.15 | 3.33 ± 0.23 |
| Dried | − | 60 | 1.16 | 3.69 ± 0.18 | 0.80 ± 0.00 | 2.89 ± 0.18 |
| Plastic | ||||||
| Wet | + | 16 | 0.31 | 5.25 ± 0.18 | 0.91 ± 0.11 | 4.33 ± 0.21 2 |
| Wet | − | 60 | 1.16 | 4.25 ± 0.18 | 0.30 ± 0.00 | 3.95 ± 0.18 1 |
| Dried | + | 90 | 1.74 | 4.69 ± 0.18 | 1.80 ± 0.21 | 2.89 ± 0.28 |
| Dried | − | 90 | 1.74 | 5.80 ± 0.16 | 2.58 ± 0.29 | 3.22 ± 0.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gardner, A.; Ghosh, S.; Dunowska, M.; Brightwell, G. Virucidal Efficacy of Blue LED and Far-UVC Light Disinfection against Feline Infectious Peritonitis Virus as a Model for SARS-CoV-2. Viruses 2021, 13, 1436. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081436
Gardner A, Ghosh S, Dunowska M, Brightwell G. Virucidal Efficacy of Blue LED and Far-UVC Light Disinfection against Feline Infectious Peritonitis Virus as a Model for SARS-CoV-2. Viruses. 2021; 13(8):1436. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081436
Chicago/Turabian StyleGardner, Amanda, Sayani Ghosh, Magdalena Dunowska, and Gale Brightwell. 2021. "Virucidal Efficacy of Blue LED and Far-UVC Light Disinfection against Feline Infectious Peritonitis Virus as a Model for SARS-CoV-2" Viruses 13, no. 8: 1436. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081436