
Tree Shrew as an Emerging Small Animal Model for Human Viral Infection: A Recent Overview
by
 ,
,
Mohammad Enamul Hoque Kayesh
1,2 ,
,
Takahiro Sanada
3,
Michinori Kohara
3 and
Kyoko Tsukiyama-Kohara
1,* 1
Transboundary Animal Diseases Centre, Joint Faculty of Veterinary Medicine, Kagoshima University, Kagoshima 890-0065, Japan
2
Department of Microbiology and Public Health, Faculty of Animal Science and Veterinary Medicine, Patuakhali Science and Technology University, Barishal 8210, Bangladesh
3
Department of Microbiology and Cell Biology, Tokyo Metropolitan Institute of Medical Science, Tokyo 156-8506, Japan
*
Author to whom correspondence should be addressed.
Viruses 2021, 13(8), 1641; https://0-doi-org.brum.beds.ac.uk/10.3390/v13081641
Submission received: 30 June 2021
/
Revised: 4 August 2021
/
Accepted: 16 August 2021
/
Published: 18 August 2021
(This article belongs to the Special Issue Basic Studies for Vaccine Development Targeting Virus Infections)
Abstract
:Viral infection is a global public health threat causing millions of deaths. A suitable small animal model is essential for viral pathogenesis and host response studies that could be used in antiviral and vaccine development. The tree shrew (Tupaia belangeri or Tupaia belangeri chinenesis), a squirrel-like non-primate small mammal in the Tupaiidae family, has been reported to be susceptible to important human viral pathogens, including hepatitis viruses (e.g., HBV, HCV), respiratory viruses (influenza viruses, SARS-CoV-2, human adenovirus B), arboviruses (Zika virus and dengue virus), and other viruses (e.g., herpes simplex virus, etc.). The pathogenesis of these viruses is not fully understood due to the lack of an economically feasible suitable small animal model mimicking natural infection of human diseases. The tree shrew model significantly contributes towards a better understanding of the infection and pathogenesis of these important human pathogens, highlighting its potential to be used as a viable viral infection model of human viruses. Therefore, in this review, we summarize updates regarding human viral infection in the tree shrew model, which highlights the potential of the tree shrew to be utilized for human viral infection and pathogenesis studies.
1. Introduction
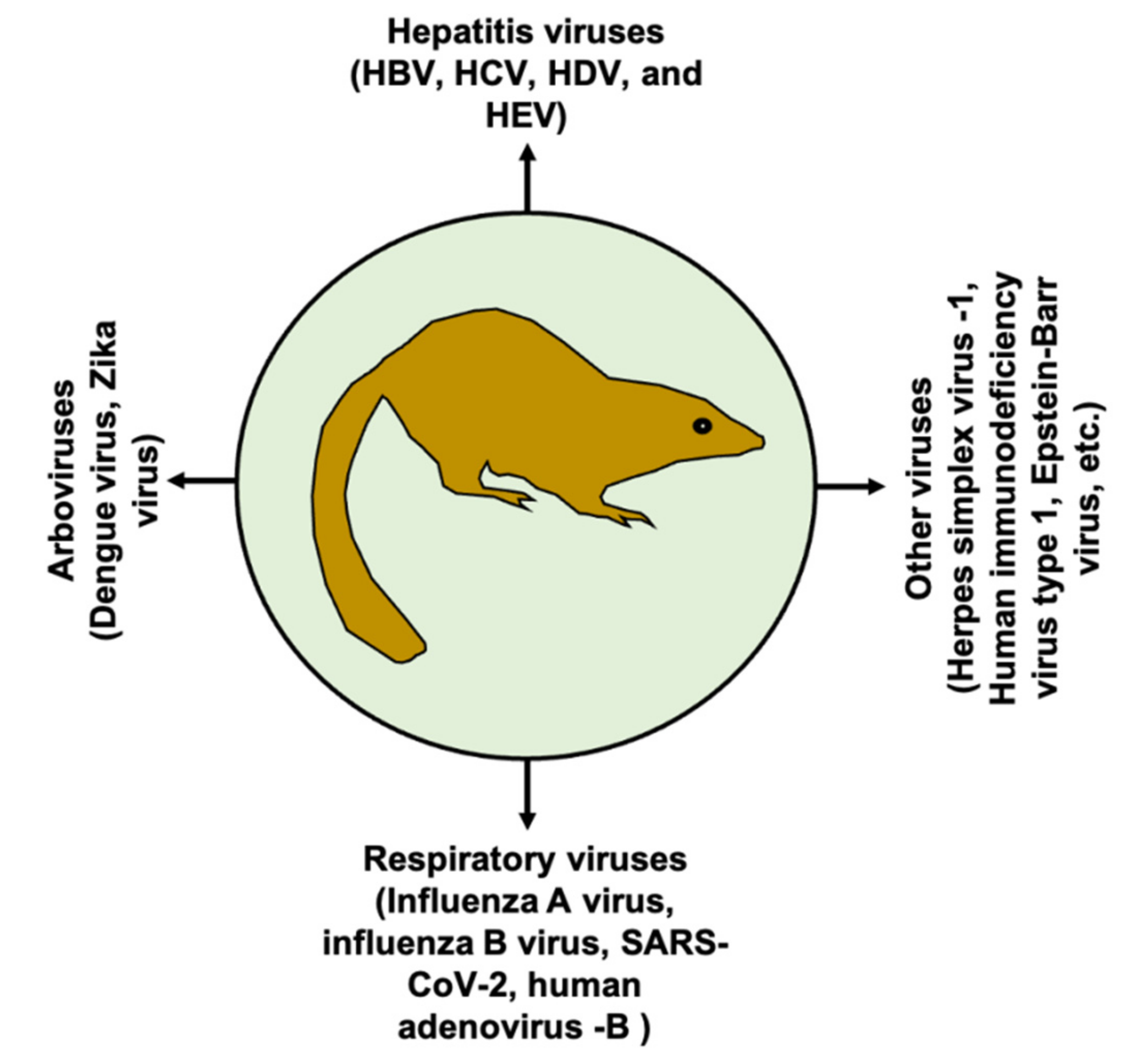
Viral infection is a major threat to public health, causing millions of deaths worldwide. Animal models mimicking human diseases are invaluable tools to study viral infection and pathogenesis, as well as for the development of prophylactic and/or therapeutic interventions. In the medical community, the term “tree shrew” or “Tupaia” refers to Tupaia belangeri (northern tree shrew) or T. belangeri chinensis (Chinese tree shrew); it belongs to the order Scandentia, which splits into two families, Ptilocercidae and Tupaiidae [1]. Tree shrews belong to the Tupaiidae family and the genus Tupaia, which includes 16 species [2,3]. Taxonomically, T. belangeri chinensis is a subspecies of T. belangeri, but genetically they show differences [2]. In addition, high genetic diversity within T. belangeri is also observed [1]. Moreover, genetic variation was also noticed within T. belangeri chinensis [4]. Although the tree shrew shows potential as animal model, genetic instability should be carefully considered during evaluation of experimental results. Tree shrews are a squirrel-like non-primate mammal with a body weight ranging between 50 and 270 g. Tree shrews are widely distributed in Southeast Asia, including South and Southwest China. At the age of six to nine months old, tree shrews can give birth on average to four babies with a gestation period of 45 days [3]. Tree shrews are genetically closer to humans than to rodents [5,6]. The tree shrew has been extensively used as an animal model for biomedical research of different conditions, including myopia [7], depression [8,9], cancer [10,11,12], chemotherapy models [13], and non-alcoholic fatty liver disease [14], to name a few. Of note, the tree shrew has also been used for viral infections, and its use in human viral infections is increasing; thus, the tree shrew emerges as a potential small animal model in advancing the knowledge of viral pathogenesis and diseases [3,15]. At the present time, tree shrews have been used as models for several important human viral pathogens (Figure 1).
The tree shrew has unique characteristics that render it a potentially advantageous animal model, including small body size, short reproductive cycle and life span, easy handling, low maintenance cost, and genetic closeness to primates. Although the tree shrew appears as a promising animal model, its extensive use is limited by the availability of tree shrew-specific reagents, individual variability, etc. [16]. As the whole genome sequence and tree shrew database are available now, these limitations should gradually decrease and the use of the tree shrew should increase. Here, we provide an update on the tree shrew viral infection model for human-infecting viruses (Table 1), including HBV, HCV, influenza virus, SARS-CoV-2, HAdV, ZIKV, and DENV infection and pathogenesis, which will enhance our understanding.
2. Tree Shrew Model for Hepatitis B and Hepatitis C Virus
2.1. Hepatitis B Virus (HBV)
HBV is a major global public health problem causing chronic hepatitis, liver cirrhosis, and/or hepatocellular carcinoma (HCC) [37]. HBV has a very restricted host range, and previously the chimpanzee (Pan troglodytes) has been used as the only natural-infection model [38]; however, its use in experimental infection has become difficult due to high expense and ethical and animal welfare issues. An alternative HBV-permissive animal model is essential, and the tree shrew has been reported to be susceptible to HBV infection. However, the infection rate is very low, and the tree shrew transiently carries the virus [18,39]. However, with the use of tree-shrew adapted HBV virus, the limitation of low infection rate in the tree shrew model should be overcome [39]. HBV induces HCC in tree shrews which is synergistically enhanced with aflatoxin B1 exposure [40]. Including our group, several groups have been working to establish a tree shrew HBV infection model. The tree shrew has been modeled for both acute and chronic HBV infection [17,19,20,41,42]. In our previous study, we observed a varied replication efficiency due to HBV genotype differences, where HBV-A2 propagation was higher than that of HBV-C; however, the overall viral titer was very low [17]. However, the effect of high genetic variation of the animal on the replication efficiency of HBV genotypes cannot be ruled out. There are eight well-known HBV genotypes (A–H) [43], which are characterized by greater than 8% difference in the nucleotide sequence of the HBV genome [44], and the characterization of all the HBV genotypes in tree shrew is yet to be completed. In our study, we observed that interferon β (IFN-β) was significantly suppressed in chronic HBV infection at 31 weeks post infection in all HBV-infected tree shrews, suggesting the impairment of the interferon response in chronic infection, whereas in acute infection the IFN-β response was variable [17]. Tree shrews show a tendency of chronic infection when infected as neonates and similar liver histopathological changes are observed as in human infections [17,19,20]. Additionally, primary tupaia hepatocytes (PTHs) are widely used for HBV infection studies, which should provide further insights into HBV pathogenesis [45,46,47]. Of note, sodium taurocholate co-transporting polypeptide (NTCP), the cellular entry receptor for HBV, was recently identified using PTHs [45]. Recently, transplantation of tupaia primary hepatocytes into chimeric mice were found to be useful and resulted in efficient HBV infection [47]. It has been reported that tumor necrosis factor α (TNF-α) can suppress HBV replication in PTHs [48]. Thus, the tree shrew model of HBV infection has been an invaluable tool for HBV study.
2.2. Hepatitis C Virus (HCV)
HCV is a major global health problem; it infects approximately 130–170 million people worldwide and causes chronic liver disease, cirrhosis, and HCC [49]. Similar to HBV, HCV also has a narrow host range, with spreading replication reported exclusively in humans and chimpanzees. Lack of small animal models is a significant obstacle in the field of HCV research. However, in vitro and in vivo tree shrew models have been very useful in the investigation of many aspects of the HCV life cycle, including HCV replication, pathogenesis and antiviral interventions [22,50,51,52,53]. However, HCV-infected tree shrews showed intermittent viral propagation and low viral load [21,22,23,51], which is suggestive of restricted replication in the tree shrew compared to humans. Additionally, liver histopathological changes observed in tree shrews are comparable to human cases [21,22]. Notably, a previous study showed the development of an acute infection that led to chronicity in the tree shrew model which was characterized by the development of liver steatosis, cirrhotic nodules and tumorigenesis [21]. In our previous study we demonstrated the HCV-specific antibody response in the tree shrew, although the titers were found to fluctuate [22], suggesting the tree shrew might be useful in preclinical vaccine efficacy testing. Moreover, the innate immune response, such as toll-like receptors and cytokines, were found to be induced in HCV infection [22], suggesting the tree shrew could serve as a potential model for an immune response and pathogenesis study.
3. Tree Shrew Model for Respiratory Viruses
3.1. Influenza Virus
The use of the tree shrew in influenza research was first introduced in 2013. As demonstrated, the tree shrew supports the replication of human influenza viruses without prior adaptation, producing mild disease symptoms [24]. The entry receptors of both human influenza virus (SAα2,6 Gal) and avian influenza virus (SAα2,3 Gal) were found to be distributed in the tree shrews’ respiratory tract [24]. In another study, it has been demonstrated that avian influenza A (H9N2) virus infection in tree shrews and ferrets showed comparable pathogenicity [25]. There was induction of a variable cytokine response in respiratory tissues at different days post-infection (dpi), including TNF-α, IFN-β, IL-4, IL-6, CXCL8 (IL-8), IL-10, IL-13, CXCL10 (IP-10), CCL5 (RANTES), CXCL9 (MIG), and CCL2 (MCP-1) [25]. In our previous study, we observed that multisite inoculation of H5N1 and H7N9 avian influenza virus with 1.0 × 106 plaque-forming units (PFU) produced severe diffuse pneumonia and focal pneumonia, respectively [26]. The severity of pneumonia was found to be correlated with proinflammatory cytokine transcript levels. Moreover, H5N1 also induced fever and body weight loss; however, no effect on body weight change by H7N9 was observed. We also observed antibody induction in tree shrews infected with H5N1 and H7N9 [26], suggesting that the tree shrew could be used for evaluating antigen-specific antibody response and for vaccine efficacy studies [54]. A recent study evaluated the infectivity and transmissibility of three unadapted different strains of influenza A virus, including pandemic H1N1 (A/Sichuan/1/2009, pdmH1N1), avian-origin H5N1 (A/Chicken/Gansu/2/2012, H5N1) and early human-origin H7N9 (A/Suzhou/SZ19/2014, H7N9), in tree shrews, and showed efficient replication and subclinical symptoms in tree shrews [27]. A strong humoral immune response was induced in virus-inoculated tree shrews, which was protective in a challenge with a homologous virus [27]. In a recent study, it has been demonstrated that intranasal inoculation of unadapted human influenza B (V0215 and Y12) with 1.0 × 106 TCID50 could infect tree shrews and produce milder nasal secretions and weight loss with no significant respiratory symptoms. However, an increase in cytokine levels of IL-6, IL-10, IP-10, TNF-α and TGF-β mRNA in tissues were observed [28]. These results indicated the suitability of tree shrews in studies of influenza viruses, particularly considering their relatively small size, evolutionary status as closer to primates and lower cost compared to ferrets. Findings obtained from different studies with influenza virus infection in tree shrews suggest that the tree shrew is a viable animal model for influenza research.
3.2. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2)
SARS-CoV-2 is the causal agent of the ongoing COVID-19 pandemic, which emerged in 2019 in Wuhan, China, spread globally and resulted millions of deaths [55,56]. SARS-CoV-2 belongs to the family Coronaviridae and the genus Betacoronavirus, and SARS-CoV-2 is the third CoV that involved spillover of an animal coronavirus to humans [57]. The pathogenesis of SARS-CoV-2 is complex and poorly understood [58]. Different animal models have been investigated for advancing the understanding of SARS-CoV-2 pathogenesis and host response, including mouse models, Syrian hamster model, ferret models, non-human primate models, and additional animal models including mink, cat, dog, pig, chicken, duck, and fruit bats [59]. Moreover, a suitable animal model is crucial for the development of targeted preventions and treatments. A recent study showed that tree shrews could be infected with SARS-CoV-2 strain 107 (1 × 107 TCID50) using multiple routes, such as oral, intranasal, and ocular routes. Viral load peaked at 3 dpi in lung tissues, and histopathology revealed hemorrhage in lung tissues of infected animals [30]. However, this study found old tree shrews (5–6-years-old) were less susceptible to SARS-CoV-2 infection compared to adult tree shrews (1-year-old), which is not compatible with cases as reported in humans, where older humans are more susceptible to SARS-CoV-2 infection than younger humans [60]. Another study reported increased body temperature in SARS-CoV-2-inoculated tree shrews, particularly in female animals; however, no clinical signs were observed [29], indicating the inconsistency of results in the study of SARS-CoV-2 infection in the tree shrew model, which could be due to its high genetic variation. However, the possibility of the tree shrew as an asymptomatic carrier or reservoir cannot be ruled out, which requires future investigation.
3.3. Human Adenoviruses (HAdVs)
HAdVs are non-enveloped, double-stranded DNA viruses with a genome of 36 kb belonging to the Adenoviridae family [61]. At the present time, seven HAdV species (A–G) and over 100 types (http://hadvwg.gmu.edu/; accessed on 30 May 2021) have been reported, based on genome sequencing, and more types remain to be identified [62]. HAdV infection often causes mild diseases; however, life-threatening respiratory disease may occur. Moreover, based on the infecting HAdV genotype, the clinical manifestation largely varies [63], and pneumonia has been reported as one of the more serious types of HAdV-induced respiratory disease in children [64]. A suitable animal model is essential for studying pathogenicity, promoting antiviral and vaccine development, and conducting preclinical efficacy testing of oncolytic adenoviral vectors. Moreover, a permissive animal model for HAdV-B infection is still lacking [31], which is crucial for a proper understanding of pathogenesis and host response. A recent study by Li et al. showed that tree shrews demonstrate susceptibility to HAdV-B infection. They showed that intranasally inoculated HAdV-55 efficiently replicates and produces severe interstitial pneumonia in tree shrews [31]. However, a pre-existing neutralizing antibody (nAb) was found in 6 tree shrews out of 18 against HAdV-3, and 1 had a preexisting nAb against HAdV-14, suggesting the occurrence of natural infection of HAdV in tree shrews. Notably, AdVs are wide spread and infect all five taxa of vertebrates. It has been reported that Tupaia adenovirus (TAV) infects the members of Tupaiidae family, and TAV has a genome of 33.5 kb [65]. However, there were no pre-existing nAbs against HAdV-55 in those tree shrews [31]. The overall findings of the study demonstrated the tree shrew as a permissive animal model for studying HAdV-B infection which may be useful for antiviral testing.
4. Tree Shrew Model for Arboviruses
4.1. Zika Virus (ZIKV)
Emerging and reemerging pathogens constitute a major public health threat globally [66]. ZIKV is a mosquito-borne flavivirus in the Flaviviridae family containing a positive-sense, single-stranded RNA genome of approximately 10.7 kb in size. ZIKV contains two major geographically different lineages, the Asian and African lineages [67]. Recently, ZIKV has spread worldwide and the WHO declared ZIKV fever as a public health emergency [68]. ZIKV has been associated with fetal microcephaly and serious neurological complications in adults [69]. A proper understanding of ZIKV pathogenesis and host-virus interaction is crucial for antiviral and vaccine development, for which a suitable small animal model is highly essential. There has been much progress in understanding of the ZIKV pathogenesis and host response using several animal models, including mice and non-human primates [70]; however, each model has its limitations. Therefore, an alternative animal model is required. Zhang et al. showed that tree shrews subcutaneously inoculated with ZIKV strain GZ01 (106 PFU/animal) produced transient viremia, cutaneous inflammation and notable dermatological manifestations of skin rashes, as commonly observed in human patients [32]. However, there was no induction of fever or other neurological and behavioral abnormalities. Notably, ZIKV infection in adult tree shrews induced an antibody response that prevented reinfection with a homologous virus [32], suggesting the tree shrew model could be used for vaccine candidate screening. During neonatal infection of 1-day-old tree shrews through the intracerebral route with 105 PFU of ZIKV, despite the detection of high levels of ZIKV RNA in the brain at 3 and 6 dpi and substantial pathological changes of neuron destruction, no signs of microcephaly in the ZIKV-infected tree shrews was observed [32]. In another study, in vitro tissue tropism of ZIKV infection in the tree shrew has been demonstrated, and the obtained findings indicated the susceptibility of the tree shrew primary cells from the kidney, lung, liver, skin and aorta to ZIKV infection [71]. Moreover, the antiviral innate immune response was induced, as observed by the induction of different cytokines, including IL-6, IL-8, TNF-α, IFN-β, CXCL9, and MX1 [71]. Although further studies are required, the above findings highlight the potential of the tree shrew for ZIKV infection and pathogenesis studies.
4.2. Dengue Virus (DENV)
DENV is the causal agent of dengue, a rapidly spreading mosquito-borne viral disease of tropical and sub-tropical areas. DENV belongs to the family Flaviviridae and the genus Flavivirus, and there are four DENV serotypes, DENV-1, DENV-2, DENV-3, and DENV-4 [72]. A suitable small animal model mimicking dengue in humans is still lacking. Previously, we showed the susceptibility of tupaia fibroblast cells to DENV infection with all four serotypes, where an upregulation of TLR8 and IL-8 mRNA expression was also observed in DENV-infected tupaia cells compared to uninfected cells [73]. Recently, the tree shrew animal model for dengue has been described by Jiang et al. as a novel model, and intravenous or subcutaneous inoculation of DENV-2 and DENV-3 in tree shrews showed some characteristics of dengue in humans [16]. As reported, there was an increased body temperature in DENV-infected tree shrews compared to uninfected animals. Moreover, proliferation of DENV and pathological changes in the brain were observed; however, viremia was very low [16]. Notably, no manifestation of severe dengue was reported. To extend the utility of the tree shrew dengue model, further research is required, including tree shrew-adapted DENV strain development, investigation of infection and pathogenesis with other serotypes, and suitability for studying the antibody-dependent enhancement (ADE) effect in the tree shrew model. Moreover, the susceptibility of tree shrews to other members of the flavivirus genus such as West Nile virus and Japanese encephalitis virus could be investigated, which should enhance the characterization of these viral infections and their pathogenesis.
5. Tree Shrew Model for Other Human Viruses
5.1. Herpes Simplex Virus (HSV)
HSV primarily infects human populations and causes a contagious infection affecting approximately 60–95% of adults globally. The HSV belongs to the family Herpesviridae, and has 2 types, HSV type 1 (HSV-1) and HSV type 2 (HSV-2) [74]. HSV-1 mainly infects the mouth, pharynx, face, eye, and central nervous system (CNS), while HSV-2 primarily infects the anogenital region [75,76]. An earlier report showed experimental infection of HSV-1 and HSV-2 (25–105 PFU/animal) in juvenile (28–45 days old) and adult (150 days old) tree shrews when inoculated intravenously, intraperitoneally, or subcutaneously. This resulted in deaths in juvenile animals and acute illness in adults [36]. Natural infection of HSV-2 in tree shrews (1/272) was reported previously, as confirmed by the presence of anti-HSV-2 IgG [77]. A recent study reported that HSV-1 infection in tree shrew trigeminal ganglia (TG) through ocular inoculation has important similar features to human infection regarding expression patterns of key HSV-1 viral genes, including LAT and ICP0 during latency, that differs significantly from mice. Both the LAT and ICP0 regions were robustly transcribed in tree shrews and human TG during latency; however, only the LAT region was transcribed in mouse TG. Additionally, spontaneous reactivation of HSV-1 infection from latency in tree shrews has been reported [33], providing an alternative tool to study HSV-1 infection and pathogenesis. A previous study reported that HSV-1 can latently infect the CNS of the tree shrew [35]. Notably, the eye structures of tree shrew are similar to humans, and a recent study demonstrated that ocular inoculation of HSV-1 causing infection in tree shrew corneas resulted in inflammation of the cornea, known as herpes simplex virus keratitis (HSK), as observed in humans [34], suggesting that the tree shrew could be a useful model for studying HSK. Overall, HSV-1-related disease manifestations as observed in humans, such as latent infections and reactivations, are also observed in tree shrews, which highlights the suitability of the tree shrew model of HSV-1 infection.
5.2. Human Immunodeficiency Virus Type 1 (HIV-1)
A recent study characterized the ability of tree shrew cells to support HIV-1 in vitro. This study showed that the exogenous expression of human CD4 and CCR5 362 molecules in tree shrew cells support HIV-1 entry and replication and showed that the tree shrew CXCR4 serves as a functional co-receptor [78]. The tree shrew cytidine deaminases of the APOBEC3 family have been reported as the host restriction factor for HIV-1 in tree shrew cells, which exerted a strong inhibitory effect on HIV-1 replication [78]. Therefore, the findings of the study discovered APOBEC3 proteins as a barrier to HIV-1 infection; this information is critical for developing gene-edited tree shrew HIV-1-infected models. Previously, induction of hypermutations in the HIV-1 genome by APOBEC3 proteins was reported [79].
6. Conclusions
The development of animal models is essential for studying the pathogenesis of viral infections, as well as for evaluating the efficacy of drugs and vaccines. The use of the tree shrew model for viral infection is increasing due to its susceptibility to several important human viral pathogens. No animal models can be a perfect alternate to the human infection model. However, animal models mimicking human diseases are crucial for properly understanding the host response and pathogenesis, and at the same time, experiments should be conducted in such a way that the results obtained using animal models can be translated to humans. As the ethical and financial concerns limit the use of chimpanzees in hepatitis virus research, the tree shrew model could be developed. However, high genetic variation within this species may hinder a wide use of this animal in biomedical research. So far, no inbred line of tree shrews has been reported, which is required to be developed to reduce the individual variations found in wild tree shrews. Moreover, there is the challenge of the overall low viral titer in tree shrews and the scarcity of the research tools that should be overcome. However, the development of inbred tree shrews is underway [4], and a recent report of transgenic tree shrews has been released [80], which should further enhance the widespread use of tree shrews in biomedical research, including viral infections.
Author Contributions
Conceptualization, M.E.H.K., T.S., M.K. and K.T.-K.; writing—original draft preparation, M.E.H.K.; writing—review and editing, M.E.H.K., T.S., M.K. and K.T.-K.; supervision, K.T.-K. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by a grant (innovative drug development network) from the Japan Agency for Medical Research and Development (AMED).
Acknowledgments
Authors would like to thank Md Abul Hashem, Bouchra Kitab, Hai-Ying Chi, and Sayeh Ezzikouri for their cordial collaboration in Tupaia experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Roberts, T.E.; Lanier, H.C.; Sargis, E.J.; Olson, L.E. Molecular phylogeny of treeshrews (Mammalia: Scandentia) and the timescale of diversification in Southeast Asia. Mol. Phylogenetics Evol. 2011, 60, 358–372. [Google Scholar] [CrossRef]
- Xu, L.; Chen, S.-Y.; Nie, W.-H.; Jiang, X.-L.; Yao, Y.-G. Evaluating the Phylogenetic Position of Chinese Tree Shrew (Tupaia belangeri chinensis) Based on Complete Mitochondrial Genome: Implication for Using Tree Shrew as an Alternative Experimental Animal to Primates in Biomedical Research. J. Genet. Genom. 2012, 39, 131–137. [Google Scholar] [CrossRef]
- Tsukiyama-Kohara, K.; Kohara, M. Tupaia belangeri as an Experimental Animal Model for Viral Infection. Exp. Anim. 2014, 63, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Xiao-Hong, L.; Yong-Gang, Y. Characterization of 12 polymorphic microsatellite markers in the Chinese tree shrew (Tupaia belangeri chinensis). Zool. Res. 2013, 34, 62. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Huang, Z.-Y.; Cao, C.-C.; Chen, C.-S.; Chen, Y.-X.; Fan, D.-D.; He, J.; Hou, H.; Hu, L.; Hu, X.-T.; et al. Genome of the Chinese tree shrew. Nat. Commun. 2013, 4, 1426. [Google Scholar] [CrossRef] [Green Version]
- Sanada, T.; Tsukiyama-Kohara, K.; Shin, I.T.; Yamamoto, N.; Kayesh, M.E.H.; Yamane, D.; Takano, J.-I.; Shiogama, Y.; Yasutomi, Y.; Ikeo, K.; et al. Construction of complete Tupaia belangeri transcriptome database by whole-genome and comprehensive RNA sequencing. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Norton, T.T. Experimental Myopia in Tree Shrews. Novartis Found. Symp. 2007, 155, 178–209. [Google Scholar] [CrossRef]
- Hai-Ying, C.; Nagano, K.; Ezzikouri, S.; Yamaguchi, C.; Kayesh, M.E.H.; Rebbani, K.; Kitab, B.; Nakano, H.; Kouji, H.; Kohara, M.; et al. Establishment of an intermittent cold stress model using Tupaia belangeri and evaluation of compound C737 targeting neuron-restrictive silencer factor. Exp. Anim. 2016, 65, 285–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Shen, F.; Li, C.; Li, Y.; Wang, X. Depression-like behaviors in tree shrews and comparison of the effects of treatment with fluoxetine and carbetocin. Pharmacol. Biochem. Behav. 2016, 145, 1–8. [Google Scholar] [CrossRef]
- Hai-Ying, C.; Tanaka, Y.; Hifumi, T.; Shoji, K.; Kayesh, M.E.H.; Hashem, A.; Kitab, B.; Sanada, T.; Fujiyuki, T.; Yoneda, M.; et al. Pathological and genetic aspects of spontaneous mammary gland tumor in Tupaia belangeri (tree shrew). PLoS ONE 2020, 15, e0233232. [Google Scholar] [CrossRef] [PubMed]
- Ge, G.-Z.; Xia, H.-J.; He, B.-L.; Zhang, H.-L.; Liu, W.-J.; Shao, M.; Wang, C.-Y.; Xiao, J.; Ge, F.; Li, F.-B.; et al. Generation and characterization of a breast carcinoma model by PyMT overexpression in mammary epithelial cells of tree shrew, an animal close to primates in evolution. Int. J. Cancer 2015, 138, 642–651. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.; Peng, H.; Zhong, L.; Wu, P.; He, J.; Deng, Z.; Huang, Y. The Tree Shrew as a Model for Cancer Research. Front. Oncol. 2021, 11, 653236. [Google Scholar] [CrossRef]
- Li, Y.; Su, J.-J.; Qin, L.-L.; Egner, P.A.; Wang, J.-S.; Groopman, J.D.; Kensler, T.W.; Roebuck, B.D. Reduction of aflatoxin B(1) adduct biomarkers by oltipraz in the tree shrew (Tupaia belangeri chinensis). Cancer Lett. 2000, 154, 79–83. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Z.; Li, Y.; Liao, S.; Wu, X.; Chang, Q.; Liang, B. Cholesterol induces lipoprotein lipase expression in a tree shrew (Tupaia belangeri chinensis) model of non-alcoholic fatty liver disease. Sci. Rep. 2015, 5, 15970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayesh, M.E.H.; Hashem, A.; Kitab, B.; Tsukiyama-Kohara, K. Pathogenesis and Immune Response Caused by Vector-Borne and Other Viral Infections in a Tupaia Model. Microorganisms 2019, 7, 686. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Lu, C.; Sun, Q. Tree Shrew as a New Animal Model for the Study of Dengue Virus. Front. Immunol. 2021, 12, 621164. [Google Scholar] [CrossRef]
- Kayesh, M.E.H.; Ezzikouri, S.; Chi, H.; Sanada, T.; Yamamoto, N.; Kitab, B.; Haraguchi, T.; Matsuyama, R.; Nkogue, C.N.; Hatai, H.; et al. Interferon-β response is impaired by hepatitis B virus infection in Tupaia belangeri. Virus Res. 2017, 237, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Walter, E.; Keist, R.; Niederöst, B.; Pult, I.; Blum, H.E. Hepatitis B virus infection of tupaia hepatocytesin vitroandin vivo. Hepatology 1996, 24, 1–5. [Google Scholar] [CrossRef]
- Yang, C.; Ruan, P.; Ou, C.; Su, J.; Cao, J.; Luo, C.; Tang, Y.; Wang, Q.; Qin, H.; Sun, W.; et al. Chronic hepatitis B virus infection and occurrence of hepatocellular carcinoma in tree shrews (Tupaia belangeri chinensis). Virol. J. 2015, 12, 10–26. [Google Scholar] [CrossRef] [Green Version]
- Ruan, P.; Yang, C.; Su, J.; Cao, J.; Ou, C.; Luo, C.; Tang, Y.; Wang, Q.; Yang, F.; Shi, J.; et al. Histopathological changes in the liver of tree shrew (Tupaia belangeri chinensis) persistently infected with hepatitis B virus. Virol. J. 2013, 10, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amako, Y.; Tsukiyama-Kohara, K.; Katsume, A.; Hirata, Y.; Sekiguchi, S.; Tobita, Y.; Hayashi, Y.; Hishima, T.; Funata, N.; Yonekawa, H.; et al. Pathogenesis of Hepatitis C Virus Infection in Tupaia belangeri. J. Virol. 2010, 84, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Kayesh, M.E.H.; Ezzikouri, S.; Sanada, T.; Michinori, K.; Hayashi, Y.; Rebbani, K.; Kitab, B.; Matsuu, A.; Miyoshi, N.; Hishima, T.; et al. Oxidative Stress and Immune Responses During Hepatitis C Virus Infection in Tupaia belangeri. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Xu, X.; Chen, H.; Cao, X.; Ben, K. Efficient infection of tree shrew (Tupaia belangeri) with hepatitis C virus grown in cell culture or from patient plasma. J. Gen. Virol. 2007, 88, 2504–2512. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-F.; Zhao, J.; Zhu, Y.-T.; Wang, Y.-T.; Liu, R.; Zhao, S.-S.; Li, R.-F.; Yang, C.-G.; Li, J.-Q.; Zhong, N.-S. The tree shrew provides a useful alternative model for the study of influenza H1N1 virus. Virol. J. 2013, 10, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Yuan, B.; Xia, X.; Zhang, S.; Du, Q.; Yang, C.; Li, N.; Zhao, J.; Zhang, Y.; Zhang, R.; et al. Tree shrew as a new animal model to study the pathogenesis of avian influenza (H9N2) virus infection. Emerg. Microbes Infect. 2018, 7, 166. [Google Scholar] [CrossRef] [Green Version]
- Sanada, T.; Yasui, F.; Honda, T.; Kayesh, M.E.H.; Takano, J.-I.; Shiogama, Y.; Yasutomi, Y.; Tsukiyama-Kohara, K.; Kohara, M. Avian H5N1 influenza virus infection causes severe pneumonia in the Northern tree shrew (Tupaia belangeri). Virology 2019, 529, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, X.; Yang, J.; Wang, Z.; Jia, Y.; Han, L.; Wang, L.; Zhu, Q. Comparative Pathogenicity and Transmissibility of Pandemic H1N1, Avian H5N1, and Human H7N9 Influenza Viruses in Tree Shrews. Front. Microbiol. 2019, 10, 2955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, B.; Yang, C.; Xia, X.; Zanin, M.; Wong, S.-S.; Yang, F.; Chang, J.; Mai, Z.; Zhao, J.; Zhang, Y.; et al. The tree shrew is a promising model for the study of influenza B virus infection. Virol. J. 2019, 16, 77. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wang, J.; Kuang, D.; Xu, J.; Yang, M.; Ma, C.; Zhao, S.; Li, J.; Long, H.; Ding, K.; et al. Susceptibility of tree shrew to SARS-CoV-2 infection. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Xu, L.; Yu, D.-D.; Ma, Y.-H.; Yao, Y.-L.; Luo, R.-H.; Feng, X.-L.; Cai, H.-R.; Han, J.-B.; Wang, X.-H.; Li, M.-H.; et al. COVID-19-like symptoms observed in Chinese tree shrews infected with SARS-CoV-2. Zool. Res. 2020, 41, 517–526. [Google Scholar] [CrossRef]
- Li, X.; Zhou, Z.; Liu, W.; Fan, Y.; Luo, Y.; Li, K.; Zheng, Z.; Tian, X.; Zhou, R. Chinese tree shrew: A permissive model for in vitro and in vivo replication of human adenovirus species B. Emerg. Microbes Infect. 2021, 10, 424–438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.-N.; Zhang, L.; Deng, Y.-Q.; Feng, Y.; Ma, F.; Wang, Q.; Ye, Q.; Han, Y.; Sun, X.; Zhang, F.-C.; et al. Zika Virus Infection in Tupaia belangeri Causes Dermatological Manifestations and Confers Protection against Secondary Infection. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, E.; Ye, Y.; Zhang, K.; Yang, J.; Gong, D.; Zhang, J.; Hong, R.; Zhang, H.; Li, L.; Chen, G.; et al. Longitudinal transcriptomic characterization of viral genes in HSV-1 infected tree shrew trigeminal ganglia. Virol. J. 2020, 17, 1–13. [Google Scholar] [CrossRef]
- Li, L.; Li, Y.; Li, X.; Xia, Y.; Wang, E.; Gong, D.; Chen, G.; Yang, L.; Zhang, K.; Zhao, Z.; et al. HSV-1 infection and pathogenesis in the tree shrew eye following corneal inoculation. J. Neurovirol. 2020, 26, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, Z.; Li, X.; Wang, E.; Lang, F.; Xia, Y.; Fraser, N.W.; Gao, F.; Zhou, J. Reactivation of HSV-1 following explant of tree shrew brain. J. Neurovirol. 2016, 22, 293–306. [Google Scholar] [CrossRef] [Green Version]
- Darai, G.; Schwaier, A.; Komitowski, D.; Munk, K. Experimental Infection of Tupaia belangeri (Tree Shrews) with Herpes Simplex Virus Types 1 and 2. J. Infect. Dis. 1978, 137, 221–226. [Google Scholar] [CrossRef]
- Lee, W.M. Hepatitis B Virus Infection. N. Engl. J. Med. 1997, 337, 1733–1745. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Gao, Z.; He, W.; Ma, Y.; Feng, X.; Cai, T.; Lu, F.; Liu, L.; Li, W. microRNA expression in hepatitis B virus infected primary treeshrew hepatocytes and the independence of intracellular miR-122 level for de novo HBV infection in culture. Virology 2014, 448, 247–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, R.Q.; Su, J.J.; Huang, D.R.; Gan, Y.C.; Yang, C.; Huang, G.H. Human hepatitis B virus and hepatocellular carcinoma I. Experimental infection of tree shrews with hepatitis B virus. J. Cancer Res. Clin. Oncol. 1996, 122, 283–288. [Google Scholar] [CrossRef]
- Yan, R.Q.; Su, J.J.; Huang, D.R.; Gan, Y.C.; Yang, C.; Huang, G.H. Human hepatitis B virus and hepatocellular carcinoma II. Experimental induction of hepatocellular carcinoma in tree shrews exposed to hepatitis B virus and aflatoxin B1. J. Cancer Res. Clin. Oncol. 1996, 122, 289–295. [Google Scholar] [CrossRef]
- Li, J.; Shi, T.-D.; Han, J.-F.; Zeng, X.-G.; Fan, C.-L.; Han, C.; Liu, H.-L.; Wu, Y.-Z. A systematic study of Tupaia as a model for human acute hepatitis B infection. J. Veter. Med. Sci. 2021, 83, 21–0026. [Google Scholar] [CrossRef]
- Wang, S.-S.; Su, J.-J.; Zhang, H.-Y.; Li, Y.; Zhang, T.; Wang, Y.-C.; Huang, G.-Y.; Qin, L.-L.; Ge, X.-M.; Yu, Y.; et al. [Human hepatitis B virus infection of tree shrews and Macaca assamensis in vivo]. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi 2003, 17, 21–24. [Google Scholar] [PubMed]
- Mizuochi, T.; Okada, Y.; Umemori, K.; Mizusawa, S.; Yamaguchi, K. Evaluation of 10 commercial diagnostic kits for in vitro expressed hepatitis B virus (HBV) surface antigens encoded by HBV of genotypes A to H. J. Virol. Methods 2006, 136, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Tsuda, F.; Sakugawa, H.; Sastrosoewignjo, R.I.; Imai, M.; Miyakawa, Y.; Mayumi, M. Typing Hepatitis B Virus by Homology in Nucleotide Sequence: Comparison of Surface Antigen Subtypes. J. Gen. Virol. 1988, 69, 2575–2583. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 1. [Google Scholar] [CrossRef]
- Kock, J.; Nassal, M.; MacNelly, S.; Baumert, T.F.; Blum, H.E.; von Weizsacker, F. Efficient Infection of Primary Tupaia Hepatocytes with Purified Human and Woolly Monkey Hepatitis B Virus. J. Virol. 2001, 75, 5084–5089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanada, T.; Tsukiyama-Kohara, K.; Yamamoto, N.; Ezzikouri, S.; Benjelloun, S.; Murakami, S.; Tanaka, Y.; Tateno, C.; Kohara, M. Property of hepatitis B virus replication in Tupaia belangeri hepatocytes. Biochem. Biophys. Res. Commun. 2016, 469, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Köck, J.; Lu, Y.; Yang, D.; Lu, M.; Zhao, X. Suppression of hepatitis B virus replication in Tupaia hepatocytes by tumor necrosis factor alpha of Tupaia belangeri. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 361–368. [Google Scholar] [CrossRef]
- Lavanchy, D. The global burden of hepatitis C. Liver Int. 2009, 29, 74–81. [Google Scholar] [CrossRef]
- Zhao, X.; Tang, Z.-Y.; Klumpp, B.; Wolff-Vorbeck, G.; Barth, H.; Levy, S.; Von Weizsäcker, F.; Blum, H.E.; Baumert, T.F. Primary hepatocytes of Tupaia belangeri as a potential model for hepatitis C virus infection. J. Clin. Investig. 2002, 109, 221–232. [Google Scholar] [CrossRef]
- Feng, Y.-M.; Lu, C.; Han, Y.; Liu, L.; Sun, X.; Dai, J.; Xia, X. Tree shrew, a potential animal model for hepatitis C, supports the infection and replication of HCV in vitro and in vivo. J. Gen. Virol. 2017, 98, 2069–2078. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Zhu, Y.; Xia, X.; Liu, Y.; Feng, Y.; Hua, X.; Chen, Z.; Ding, H.; Gao, L.; Wang, Y.; et al. Tupaia CD81, SR-BI, Claudin-1, and Occludin Support Hepatitis C Virus Infection. J. Virol. 2010, 85, 2793–2802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.-C.; Riezu-Boj, J.-I.; Lasarte, J.J.; Guillén, J.; Su, J.-H.; Civeira, M.-P.; Prieto, J. Transmission of Hepatitis C Virus Infection to Tree Shrews. Virology 1998, 244, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Honda, T.; Gomi, S.; Yamane, D.; Yasui, F.; Yamamoto, T.; Munakata, T.; Itoh, Y.; Ogasawara, K.; Sanada, T.; Yamaji, K.; et al. Development and Characterization of a Highly Sensitive NanoLuciferase-Based Immunoprecipitation System for the Detection of Anti-Influenza Virus HA Antibodies. mSphere 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Coronaviridae Study Group of the International Committee on Taxonomy of, Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Harrison, A.G.; Lin, T.; Wang, P. Mechanisms of SARS-CoV-2 Transmission and Pathogenesis. Trends Immunol. 2020, 41, 1100–1115. [Google Scholar] [CrossRef]
- Muñoz-Fontela, C.; Dowling, W.E.; Funnell, S.G.P.; Gsell, P.-S.; Riveros-Balta, A.X.; Albrecht, R.A.; Andersen, H.; Baric, R.S.; Carroll, M.W.; Cavaleri, M.; et al. Animal models for COVID-19. Nature 2020, 586, 509–515. [Google Scholar] [CrossRef]
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; Junglen, S.; et al. Changes to virus taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2019). Arch. Virol. 2019, 164, 2417–2429. [Google Scholar] [CrossRef] [Green Version]
- Seto, D.; Chodosh, J.; Brister, J.R.; Jones, M.S. Members of the Adenovirus Research Community Using the Whole-Genome Sequence To Characterize and Name Human Adenoviruses. J. Virol. 2011, 85, 5701–5702. [Google Scholar] [CrossRef] [Green Version]
- Mellon, G.; Henry, B.; Aoun, O.; Boutolleau, D.; Laparra, A.; Mayaux, J.; Sanson, M.; Caumes, E. Adenovirus related lymphohistiocytic hemophagocytosis: Case report and literature review. J. Clin. Virol. 2016, 78, 53–56. [Google Scholar] [CrossRef]
- Zou, L.; Yi, L.; Yu, J.; Song, Y.; Liang, L.; Guo, Q.; Zhuang, X.; Zhang, Y.; Kang, M.; Wu, J. Adenovirus infection in children hospitalized with pneumonia in Guangzhou, China. Influ. Other Respir. Viruses 2021, 15, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Bahr, U.; Schöndorf, E.; Handermann, M.; Darai, G. Molecular anatomy of Tupaia (tree shrew) adenovirus genome; evolution of viral genes and viral phylogeny. Virus Genes 2003, 27, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.F. From “A”IV to “Z”IKV: Attacks from Emerging and Re-emerging Pathogens. Cell 2018, 172, 1157–1159. [Google Scholar] [CrossRef] [Green Version]
- Faye, O.; Freire, C.C.M.; Iamarino, A.; Faye, O.; De Oliveira, J.V.C.; Diallo, M.; Zanotto, P.M.A.; Sall, A.A. Molecular Evolution of Zika Virus during Its Emergence in the 20th Century. PLoS Negl. Trop. Dis. 2014, 8, e2636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imperato, P.J. The Convergence of a Virus, Mosquitoes, and Human Travel in Globalizing the Zika Epidemic. J. Community Health 2016, 41, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L.; Trevisan, M.; Sinigaglia, A.; Lavezzo, E.; Palù, G. Zika virus: From pathogenesis to disease control. FEMS Microbiol. Lett. 2016, 363, fnw202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, T.E.; Diamond, M.S. Animal Models of Zika Virus Infection, Pathogenesis, and Immunity. J. Virol. 2017, 91, e00009-17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Shen, Z.-L.; Feng, Y.; Li, D.-Q.; Zhang, N.-N.; Deng, Y.-Q.; Qi, X.-P.; Sun, X.-M.; Dai, J.-J.; Yang, C.-G.; et al. Infectivity of Zika virus on primary cells support tree shrew as animal model. Emerg. Microbes Infect. 2019, 8, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Simmons, C.P.; Farrar, J.; Chau, N.V.V.; Wills, B. Dengue. N. Engl. J. Med. 2012, 366, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Kayesh, M.E.H.; Kitab, B.; Sanada, T.; Hayasaka, D.; Morita, K.; Kohara, M.; Tsukiyama-Kohara, K. Susceptibility and initial immune response of Tupaia belangeri cells to dengue virus infection. Infect. Genet. Evol. 2017, 51, 203–210. [Google Scholar] [CrossRef]
- Boehmer, P.; Nimonkar, A. Herpes Virus Replication. IUBMB Life 2003, 55, 13–22. [Google Scholar] [CrossRef]
- World Health Organization. Herpes Simplex Virus; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- Chayavichitsilp, P.; Buckwalter, J.; Krakowski, A.C.; Friedlander, S.F. Herpes Simplex. Pediatr. Rev. 2009, 30, 119–130. [Google Scholar] [CrossRef]
- Han, J.-B.; Zhang, G.-H.; Duan, Y.; Ma, J.-P.; Zhang, X.-H.; Luo, R.-H.; Lü, L.-B.; Zheng, Y.-T. Sero-epidemiology of six viruses natural infection in Tupaia belangeri chinensis. Zool. Res. 2011, 32, 11–16. [Google Scholar] [CrossRef]
- Luo, M.-T.; Mu, D.; Yang, X.; Luo, R.-H.; Zheng, H.-Y.; Chen, M.; Guo, Y.-Q.; Zheng, Y.-T. Tree Shrew Cells Transduced with Human CD4 and CCR5 Support Early Steps of HIV-1 Replication, but Viral Infectivity Is Restricted by APOBEC3. J. Virol. 2021, 95. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.-T.; Fan, Y.; Mu, D.; Yao, Y.-G.; Zheng, Y.-T. Molecular cloning and characterization of APOBEC3 family in tree shrew. Gene 2018, 646, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-H.; Yan, L.-Z.; Ban, W.-Z.; Tu, Q.; Wu, Y.; Wang, L.; Bi, R.; Ji, S.; Ma, Y.-H.; Nie, W.-H.; et al. Long-term propagation of tree shrew spermatogonial stem cells in culture and successful generation of transgenic offspring. Cell Res. 2017, 27, 241–252. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Tree shrew as a model of human viral infections. The tree shrew has been modeled for several important human viral pathogens as indicated in the figure.
Figure 1.
Tree shrew as a model of human viral infections. The tree shrew has been modeled for several important human viral pathogens as indicated in the figure.


{kind=link}
Table 1.
Human viral infections in the tree shrew model.
| Virus Species | Serotype/Subtype/Genotypes/Strain | Inoculum Dose | Inoculation Route | Age of Animal | Viral Load or Titer (Plasma/Serum/Nasal Wash/Tissue) | Cytokine and Antibody Response | Clinical Signs/Strengths | Limitations | References |
|---|---|---|---|---|---|---|---|---|---|
| HBV | HBV genotype A2, C | 1 × 106–1 × 107 GEs/animal | SC, IP | Newborn or adults (1 Y) | 10–>103 copies/mL serum | IFN-β response impaired in chronic infection, and variable in acute infection; TNF-α expression changes | Immunocompetent animal model; viremia develops; development of both acute and chronic infection; occurrence of histopathological changes in the liver; detection of intrahepatic HBV DNA | Individual variations of viral replication; intermittent viremia; low viral titer | [17,18,19,20] |
| HCV | HCV (genotypes 1a, 1b, 4a, 2a, 2c, 3b, 6) | 0.03× 106–12.32×106; 6 × 105 –1 × 107 GEs/animal | IP, IV | Adults (6 M; 1 Y) | 4–~104 or 105 copies/mL serum | Intrahepatic IFN- β and IL-6 expression changes; anti-NS3 and anti-core Abs produced | Viremia develops; chronic hepatitis; progression of fibrosis; detection of intrahepatic HCV RNA | Individual variations of viral replication; intermittent viremia; low viral titer | [21,22,23] |
| Influenza A virus | H1N1 | 105 TCID50 | IN | 3-M-old | 102.94–104.24 TCID50/mL | Seroconversion | Infects tree shrews without prior adaptation; replicates in respiratory tracts; develops acute bronchopneumonia and interstitial pneumonia | No loss of appetite, congested eyes and otologic manifestations | [24] |
| Avian influenza A | H9N2 | 106 TCID50 | IN | Adults | 101.5–>105 TCID50/mL | Cytokines induced in respiratory tissues; Ab induction | Virus replication in respiratory tissues as observed on histology | Asymptomatic infection; no virus shedding | [25] |
| Avian influenza A | H5N1 | 1.0 × 106 PFU | Multisite inoculation | Adult (6M to 4Y-old) | 2.8 × 103–5.2 × 107 vRNA copies/mg of lung tissue | IFN-γ, IL-6, TNF-α induced; Ab (IgG) induced at 7 dpi | Replicates in lung tissues; develops severe diffuse pneumonia with fever and weight loss | No sneezing; effect of multisite inoculation on infectivity and pathogenesis yet to be confirmed | [26] |
| Avian influenza A | H7N9 (A/Anhui/1/2013) | 1.0 × 106 PFU | Multisite inoculation | Adult (6M to 4Y-old) | 6.5 × 101–3.4 × 104 vRNA copies/mg of lung tissue | IFN-γ induced; Ab (IgG) induced at 7 dpi | Focal pneumonia | No sneezing | [26] |
| Avian influenza A | H5N1, H7N9, H1N1, | 1.0 × 106 EID50 | IN | 1 to 4 M-old | - | Neutralizing Ab induced | Vaccine efficacy testing | [27] | |
| Influenza B virus | Strains of Yamagata or Victoria lineage | 1.0 × 106 TCID50 | IN | Adult | - | Induction of IL-6, IL-10, IP-10, TNF-α and TGF-β mRNA; seroconversion | Weight loss, milder nasal secretions | No significant respiratory symptoms | [28] |
| SARS-CoV-2 | - | 106 PFU | IN | Young (6 M to 12 M), adults (2Y to 4 Y) and old (5 Y to 7 Y) | 105.92 vRNA copies/mL (nasal wash) | - | Increased body temperature; mild pulmonary abnormalities found in histopathology | No clinical signs develop | [29] |
| SARS-CoV-2 | SARS-CoV-2 strain 107 | 1 × 107 TCID50 | Oral, intranasal, ocular | Old (5–6 Y); Adult (1 Y) | 1.36 × 103 to 9.32 × 105 copies/g lung tissue | - | vRNA detected in lung tissues and hemorrhage in lung tissues | No viremia; no data on induction of fever | [30] |
| Human adenovirus- B (HAdV-B) | HAdV-3, HAdV-7, HAdV-14, HAdV-55 | 5 × 105 TCID50 | IN | 6–8 M old | - | Increased expression of IL-8, IL-10, IL-17A, and IFN-γ; Ab produced | HAdV efficiently replicates and produced high viral load with severe interstitial pneumonia | Presence of pre-existing nAbs against HAdV-3 and HAdV-14 | [31] |
| Dengue virus (DENV) | DENV-2DENV-3 | 1.5 × 103 PFU | SC, IV | Adult | <2 × 104copies/mL serum | nAb produced | Increased body temperature | Very low viremia, lack of severe dengue manifestations | [16] |
| Zika virus (ZIKV) | ZIKV strain GZ01 | 106 PFU | SC | 5M | 104.40–106.02 RNA copies/mL (1 dpi) | Strong cytokine response induced; nAb produced | Skin rash; suitable for antivirals and vaccine testing | No fever or other neurological and behavioral abnormalities; transient viremia | [32] |
| Herpes simplex virus type 1 (HSV-1) | HSV-1 strain 17+ | 1 × 106 PFU | Ocular | 6M | - | - | Spontaneous reactivation occurs | All HSV-1 genes are not expressed | [33] |
| HSV-1 | HSV-1 McKrae virus | 1 × 106 PFU | Ocular | 6M | - | - | Herpes simplex virus keratitis develops; HSV-1 transcripts, ICP0, ICP4, and LAT detected | - | [34,35] |
| HSV-1, HSV-2 | - | 25 –105 PFU | SC, IV, IP | Juvenile (1–1.5 M) and adult (5M) | - | - | Higher pathogenicity in juveniles and caused death; herpetic hepatitis; encephalitis and fibrosis in the spleen | - | [36] |
TCID50, tissue culture dose 50; PFU, plaque-forming units; EID50, embryo infectious dose 50; GE, genome equivalents; M, month; Y, year; IN, intranasally; vRNA, viral RNA; dpi, days post-infection; nAb, neutralizing antibody.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kayesh, M.E.H.; Sanada, T.; Kohara, M.; Tsukiyama-Kohara, K. Tree Shrew as an Emerging Small Animal Model for Human Viral Infection: A Recent Overview. Viruses 2021, 13, 1641. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081641
AMA Style
Kayesh MEH, Sanada T, Kohara M, Tsukiyama-Kohara K. Tree Shrew as an Emerging Small Animal Model for Human Viral Infection: A Recent Overview. Viruses. 2021; 13(8):1641. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081641
Chicago/Turabian StyleKayesh, Mohammad Enamul Hoque, Takahiro Sanada, Michinori Kohara, and Kyoko Tsukiyama-Kohara. 2021. "Tree Shrew as an Emerging Small Animal Model for Human Viral Infection: A Recent Overview" Viruses 13, no. 8: 1641. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081641
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.