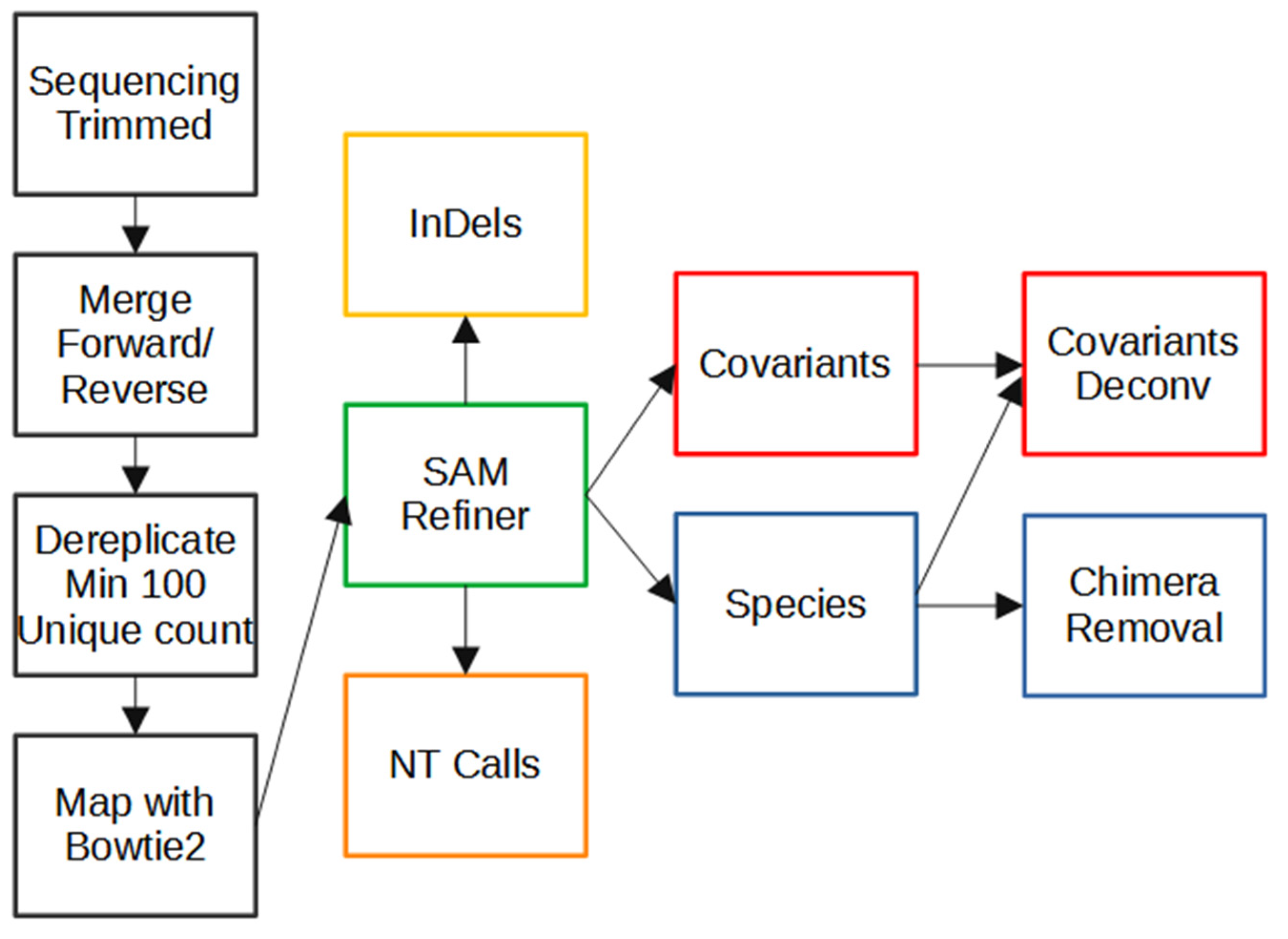
3.2. SAM Refiner: SAM Processing
Our program, SAM Refiner, is currently a command line-based python script and is available at
https://github.com/degregory/SAM_Refiner accessed on 23 June 2021 along with updated documentation. In order to run SAM Refiner, a python compiler or interpreter is needed (
https://docs.python.org/3/tutorial/interpreter.html accessed on 23 June 2021). Though only tested in a Linux environment, it should function with other common operating systems.
Figure 2 shows the command line usage for SAM Refiner. Standard SAM formatted files are the starting point for our program. These files are generated by many mapping programs, including Bowtie2 [
17] and BWA [
18]. The default functions of SAM Refiner follow. Files with the extension .sam (case insensitive) in the working directory will be identified and processed. To process SAM files, SAM Refiner must be provided a FASTA formatted file for a reference sequence using the command line argument ‘–r reference.fasta’, where the FASTA file contains the same sequence ID and sequence used to map the sequencing reads in the SAM formatted file. If the IDs of the given reference and the reference of mapped sequences in the SAM file do not match, those sequences will be ignored. If the SAM formatted files were generated from dereplicated or collapsed sequences that contain the unique read count in sequence ids where the count is at the end of the id and denoted with a ‘=’ or ‘−’, SAM Refiner will recognize the counts, i.e., ‘Seq1:1;counts = 20′ will be recognized as a sequence with 20 occurrences.
For each SAM file, SAM Refiner initially outputs 4 tab separated value (TSV) files that can be read by any standard spreadsheet software. For a SAM file with the name Sample.sam, the outputs are named Sample_unique_seqs.tsv, Sample_nt_calls.tsv, Sample_indels.tsv and Sample_covars.tsv. Example outputs of each are provided in
Supplementary Files 1, 2, 3, and 4, respectively (
https://github.com/degregory/SR_manuscript/tree/master/Supplementals accessed on 23 June 2021). All reports are based on the FASTA reference relative to the SAM formatted file, so any errors made by the mapping or incongruence between the FASTA reference and the mapping reference will result in propagated errors. The reports also include the coded amino acids and their position in the coded peptide as if the reference is an in-frame coding sequence. If multiple nucleotides in a single codon differ from the reference, they will be reported together as a MNP with the associated amino acid change. Within the files, all of the sample-specific outputs start with the name of the sample taken from the SAM file name followed in parenthesis by the count of reads mapped.
The Sample_unique_seqs.tsv file (
Supplementary 1) lists the unique sequence reads mapped in the SAM file using a variant notation to list the variations from the reference along with occurrence count and abundance. For example, using the previously mentioned SARS-CoV-2 spike ORF as the reference sequence, a sequence read that matches the reference except for having a T at position 1501 instead of the reference A would be reported simply as ‘1501A(N501Y)’. The abundance reported uses decimal notation, so 0.2 represents 20% abundance. Unique sequences that have an abundance below 0.001 are not reported.
The Sample_nt_calls.tsv file (
Supplementary 2) has a line for each nt position covered in at least 0.1% of the reads. Based on the reference sequence, each line first reports the nt position, the reference nt, the encoded amino acid position, and the amino acid residue encoded by the reference sequence. The line then reports the number of calls for each base and for deletions at that position, followed by the most abundant (primary) call and its counts and abundance. If the primary nt is different from the reference sequence, the amino acids encoded by the primary nt sequence and by the reference sequence with only that nt changed are reported. Further, if the second (secondary) and third (tertiary) most abundant nts are above 0.1% of the total read counts, those nts, their counts, abundances, and associated amino acid changes are also reported.
The Sample_indels.tsv (
Supplementary 3) file lists each insertion or deletion found in the mapping along with its occurrence count and abundance. Reported insertions have the format of ‘position-insertNT(s)’, so an insertion between nt positions 54 and 55 of the sequence ‘GCA’ will be reported as ‘55-insertGCA’. Reported deletions have the format ‘start Position-end positionDel’, so a deletion of the nts at positions 61 through 64 would be reported as ‘61-64Del’. Amino acid changes are reported if the indel maintains the reading frame. If there are no indels in the reads, no indel report will be generated.
Finally, the Sample_covars.tsv (
Supplementary 4) file lists all observed single polymorphisms and polymorphisms combinations relative to the reference sequence. The number and abundance of sequence reads containing each covariant (covar) are reported regardless of whether any of those reads have other variations or not. As an example of this processing, the sequence ‘1212G(G404G) 1501T(N501Y) 1709A(A570D)’ with 100 counts would have the covariants of ‘1212G(G404G)’, ‘1501T(N501Y)’, ‘1709A(A570D)’, ‘1212G(G404G) 1501T(N501Y)’, ‘1212G(G404G) 1709A(A570D)’, ‘1501T(N501Y) 1709A(A570D)’ and ‘1212G(G404G) 1501T(N501Y) 1709A(A570D)’, and contribute 100 counts to each. Because unique sequences that fall below the 0.1% reporting cutoff can still contribute to covariants, there may be polymorphisms in the reported covariants that are not seen in the unique sequence output. Any sequences with more than 40 polymorphisms from the reference are ignored. While all sequences with 40 or fewer polymorphisms are analyzed, only combinations of 8 or fewer polymorphisms are reported.
Once the above outputs are generated from each SAM file found, SAM Refiner will collect information from each sample and report them in a single file for the covars and unique_seqs reports (Collected_Covariances.tsv and Collected_Unique_Seqs.tsv). These collections have a threshold of 1% occurrence for reporting.
Many options are available as command line arguments that can change parameters of SAM processing of SAM Refiner (
Figure 2). There are no strictly required command line arguments, though the ‘-r’ argument is required for the SAM processing. Omitting the reference sequence will cause SAM Refiner to skip SAM processing and only perform the collections and chimera removal (see below), which require pre-existing outputs. The other input option is the ‘-S’ argument, which provides SAM Refiner with SAM files to process instead of searching the working directory. The use of dereplicated/collapsed counts in the SAM files can be disabled by using ‘--use_counts 0’. There are also options available for the outputs. All outputs can be separately suppressed with the arguments ‘--seq 0’, ‘--nt_call 0’, ‘--indel 0’, ‘--covar 0’ and ‘--collect 0’. The collections file names can be prepended with a string specified by the argument ‘--colID’. To change the reporting threshold for the sample and collected outputs, arguments ‘--min_abundance1’ and ‘--min_abundance2’ are used, respectively. For ‘--min_abundance1’, despite its name, the value can be used to either set a minimal abundance threshold or a minimal count threshold. Values of 1 or greater will set a count threshold, while those less than 1 will set an abundance threshold. Only an abundance threshold is available for ‘--min_abundance2’. All amino acid information in the reports can be suppressed with the argument ‘--AAreport 0’, which is recommended if the reference does not primarily provide an in-frame coding sequence. Users can also have all nt changes processed independently, even if they are in the same codon, with ‘--AAcodonasMNP 0’. Using ‘--ntabund’ will change the required mapped coverage threshold for reporting a position in the nt_calls output. Finally, ‘--max_dist’ and ‘--max_covar’ allow changes to covar processing and reporting. Sequences with more variations than the amount specified by ‘--max_dist’ are not included in the covar analysis. The maximum number of polymorphisms reported in a combination can be set with ‘--max_covar’. As an example, if ‘--max_covar 2’ were used for Sup. 4, then ‘1216-1216Del 1501T(N501Y) 1709A(A570D)’, ‘1212G(G404G) 1501T(N501Y) 1709A(A570D)’ and ‘1217-1217Del 1501T(N501Y) 1709A(A570D)’ would not be reported.
Using the SAM files generated from the sequencing data of the Fenton sewershed, we ran SAM Refiner with the same reference as was used for Bowtie2 mapping, the SARS-CoV-2 (NCBI Reference Sequence: NC_045512.2) spike ORF sequence. The resulting outputs can be accessed at
https://github.com/degregory/SR_manuscript/tree/master/Fenton_Data accessed on 23 June 2021. These outputs allow us to see the variant lineages present at different dates in this sewer shed. However, as can be seen in
Supplementary 1, many of the sequences reported appear to be chimeric sequences arising from template jumping. While these outputs can still be used for further analysis, removing chimeric sequences makes such analysis easier, so SAM Refiner also has methods to remove such chimeric sequences.
3.3. SAM Refiner: Chimera Removal
PCR amplification can introduce sequence errors that obscure the original template sequences. Of most concern are the introduction of false SNPs and chimeric reads. Most PCR-introduced SNPs can be removed from analysis by the use of an abundance threshold such as is the default for SAM Refiner, or as was used in our pre-processing dereplication step. There are also numerous other programs that can be used to attempt to remove such errors. Chimeric sequences are generally more difficult to remove. Many programs exist for this task; however, we were unable to find any that provided satisfying results for our amplicon sequencing. We developed two algorithms for SAM Refiner in order to remove chimeric errors arising from PCR template jumping from the SAM processing outputs. They are redundant in their function but crosschecking between the two different methods allows for increased confidence in the results.
The algorithms to remove chimeric sequences rely on the unique sequence and covariant files generated by SAM processing. The first algorithm, chimera removed (chim rm), goes through the individual unique sequences, starting with the lowest abundance, to determine if the sequences are chimeric.
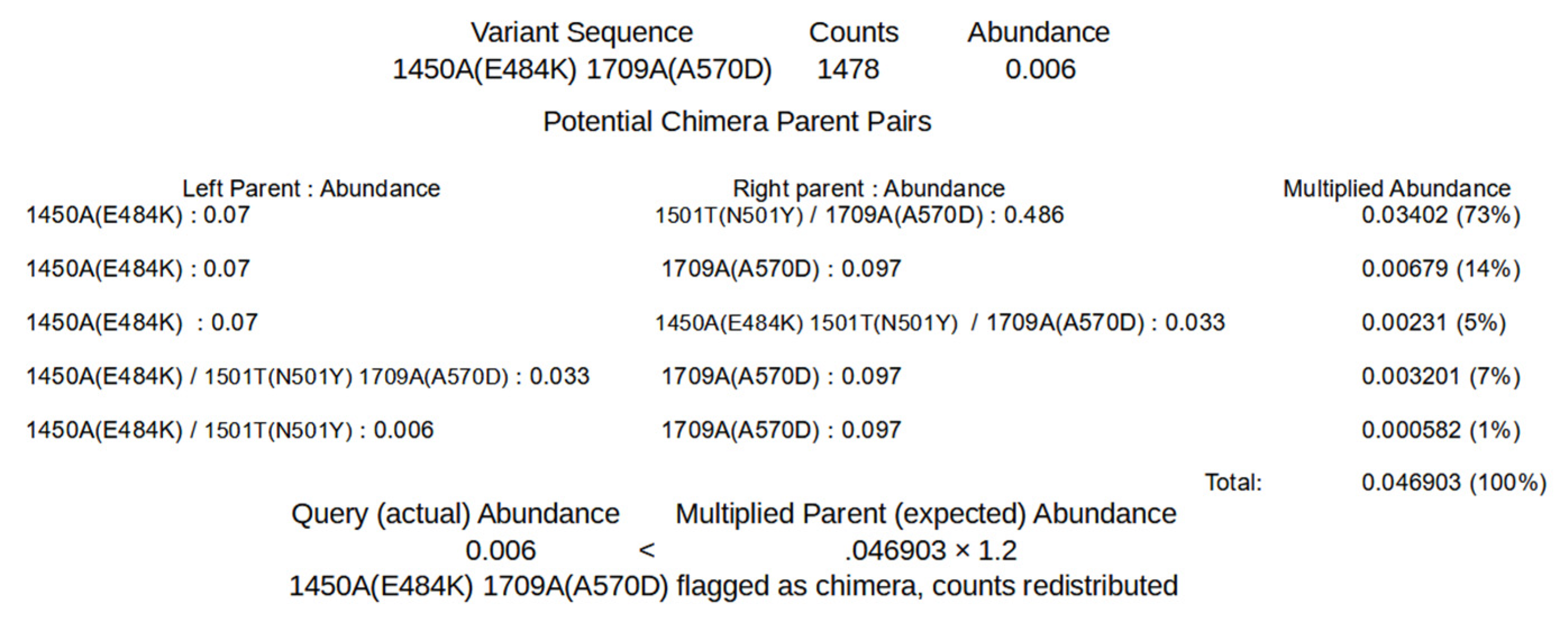
Figure 3 shows a simplified example of how the determination is made on the lowest abundant sequence of an example unique sequence output (
Supplementary 5). For this step, the sequence being considered as a potential chimera is broken up into all possible dimeric halves. Each pair is then compared to all the other sequences to detect potential parents. A sequence is flagged as a potential parent if its abundance is greater than or equal to the abundance of the potential chimera multiplied by 1.8 (foldab) and there is at least one other sequence that would be a matched parent to the complimentary dimeric half. When a pair of dimeric halves have potential parents, the abundances of parent pairs are multiplied. The products from each potential parent pairings are summed as an expected abundance value and compared to the observed abundance of the potential chimera. If the abundance of the potential chimera is less than that of the expected value multiplied by 1.2 (alpha), that sequence is flagged as a chimera and removed. The counts attributed to the flagged chimeric sequence are then redistributed to the parent sequences based on the relative expected contribution to recombination. Once this process has been performed for all the sequences, it is repeated until no more sequences are flagged as chimeric or 100 chimera removal cycles have completed. The results of this algorithm that have a recalculated abundance of 0.001 or greater are output in a new file (
Supplementary 6 Example_a1.2f1.8rd1_chim_rm.tsv). The added string represents values of the parameters used for the processing (alpha, foldab and redist; see below for more information on the parameters).
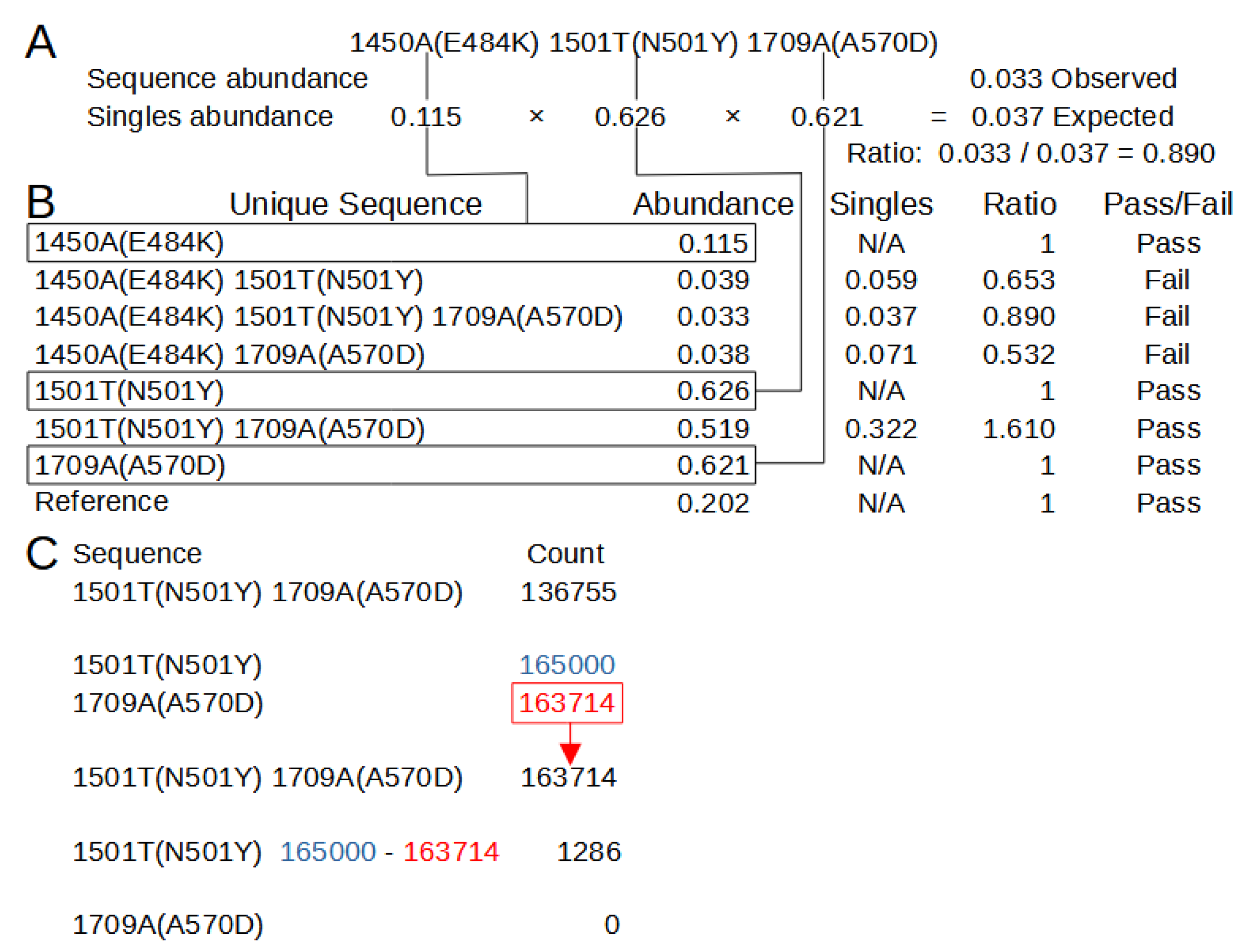
The second algorithm, covariant deconvolution (covar deconv), is a two-step process.
Figure 4 shows these processes using the example outputs found in
Supplementary 5 and 7. The first step determines if a sequence is likely to be a true or chimeric sequence by obtaining the ratio of the frequency of a given covariant sequence relative to an expected abundance of that covariant sequence assuming random recombination of its individual polymorphisms. The expected abundance is obtained by multiplying the abundances of each individual polymorphism that is present in that covariant sequence. For instance, in a sample where ‘1501T(N501Y)’ has an abundance of 0.32 and ‘1709A(A570D)’ has an abundance of 0.35, the expected abundance of the covariant ‘1501T(N501Y) 1709A(A570D)’ would be 0.112 [0.32 × 0.35]. If the ratio of the observed abundance to the expected abundance is equal to or greater than 1 (beta), that covariant passes the check and is sent to the second step. Any sequence that has an abundance of 0.3 or greater is automatically passed. If such a sequence has an observed/expected ratio less than 1, it will be assigned a ratio of 1. The second step processes the passed sequences in order of greatest observed/expected ratio to least. If multiple sequences have the same ratio, they are processed in order of greatest to least distance from the reference. Sequences that automatically pass the first step are processed after the other sequences in order of least abundant to greatest. Sequences are assigned a new occurrence count based on their constituent individual polymorphisms. For the sequence being processed, the count for the least abundant individual polymorphism is assigned to the sequence and constituent polymorphisms making up the sequence have their count reduced by the amount of the least abundant polymorphism. This reduction means the individual polymorphism that had the least counts is assigned 0 counts, so any sequence not yet processed in which that polymorphism is present is functionally removed. This process is repeated until all sequences have been reassessed or removed. The final results with an abundance of 0.001 or greater are reported in a new file (
Supplementary 8 Example_covar_deconv.tsv).
As before, the results from individual samples are collected and reported for entries above 1% occurrence. A number of command line arguments will also influence the chimera removal algorithms. Both chimera removal algorithms run by default, but either or both steps can be disabled (‘--chim_rm 0’ and ‘--covar_deconv 0’). The collections are again disabled with ‘--collect 0’. An additional output of the covariants that passed the first step of the second algorithm can be generated with ‘--pass_out 1’ (
Supplementary 9). The outputs are constrained as before by a minimum abundance with command line arguments ‘--min_abundance1’ and ‘--min_abundance2’. Collection file names are also prepended with ‘--colID’. The only input parameter that can be changed by command line argument is the abundance of sequences or covariants that will be considered in the algorithms. By default, only entries from the inputs that have a 0.001 abundance or greater are processed. This threshold can be changed with ‘--chim_in_abund’.
Four parameters can be altered for the first algorithm. The abundance ratio that is used as a threshold for selecting potential parents of a potential chimera can be set with ‘--foldab’. Larger values will generally reduce the pool of sequences that will be considered as potential parents, thus potentially reducing the total expected abundance obtained from parent pairs and the number of sequences flagged as chimeric. In the simplest theoretical model of PCR chimera generation, two parents generate one chimera. The parents have at least twice the abundance of the chimera as they would exist and have been amplified prior to the chimera, but the reality of chimera generation can be much more complex as many sequences may generate identical chimeras multiple times. If a sample has little chimera generation, a ‘--foldab’ value close to 2, such as the default of 1.8, should be sufficient to remove chimeras without also removing non-chimeric sequences in error. However, the more chimera generation observed, the more the ‘--foldab’ value needs to be reduced to accurately remove all chimeric sequences. Though it would be rare, this value can even be set to 0 so as not to exclude any sequence from being considered a potential parent. Lower values, however, will also increase the likelihood of a sequence being flagged as a chimera in error. Users may need to empirically determine the best value for their samples.
The multiplier for the parental summed abundance for determining if a sequence is a chimera can be set with ‘--alpha’. Larger values will generally result in a greater number of sequences flagged as chimeric. As with ‘--foldab’, the optimal value for ‘--alpha’ will depend on the extent of chimera generation in the samples being processed, with a value near 1 for minimal chimera generation (such as the default 1.2) and 2 or even higher for rampant chimera generation. Once again, the later would also increase the likelihood of sequences being flagged as chimeric in error.
Redistribution of the counts from the chimera to the parent sequences can be disabled with ‘--redist 0’. Redistribution is meant to give an estimate of the counts and abundances that would have been observed without chimera generation which users may wish to forgo. The maximum number of chimera removal cycles can be changed by ‘--max_cycles’, (i.e., ‘--max_cycles 2’ will only allow two iterations of the chimera removal). Multiple removal cycles allow chimeras to be found based on new counts and abundances resulting from previous cycles, increasing the likelihood chimeras are removed from a sample.
The second algorithm has two parameters that can be changed. The ratio threshold at which a covariant will be passed to the second step can be altered with ‘--beta’. The abundance at which a covariant will automatically be passed can be changed with ‘--autopass’.
The chimera removal methods of SAM Refiner were also used on the Fenton sewershed sequencing data. Due to the relatively high amount of chimeric sequences in our samples, we used the command line arguments ‘--foldab = 0.6 –alpha = 2.2’. The outputs generated for the Fenton sewershed from 2-2-21 to 4-13-21 can be accessed at
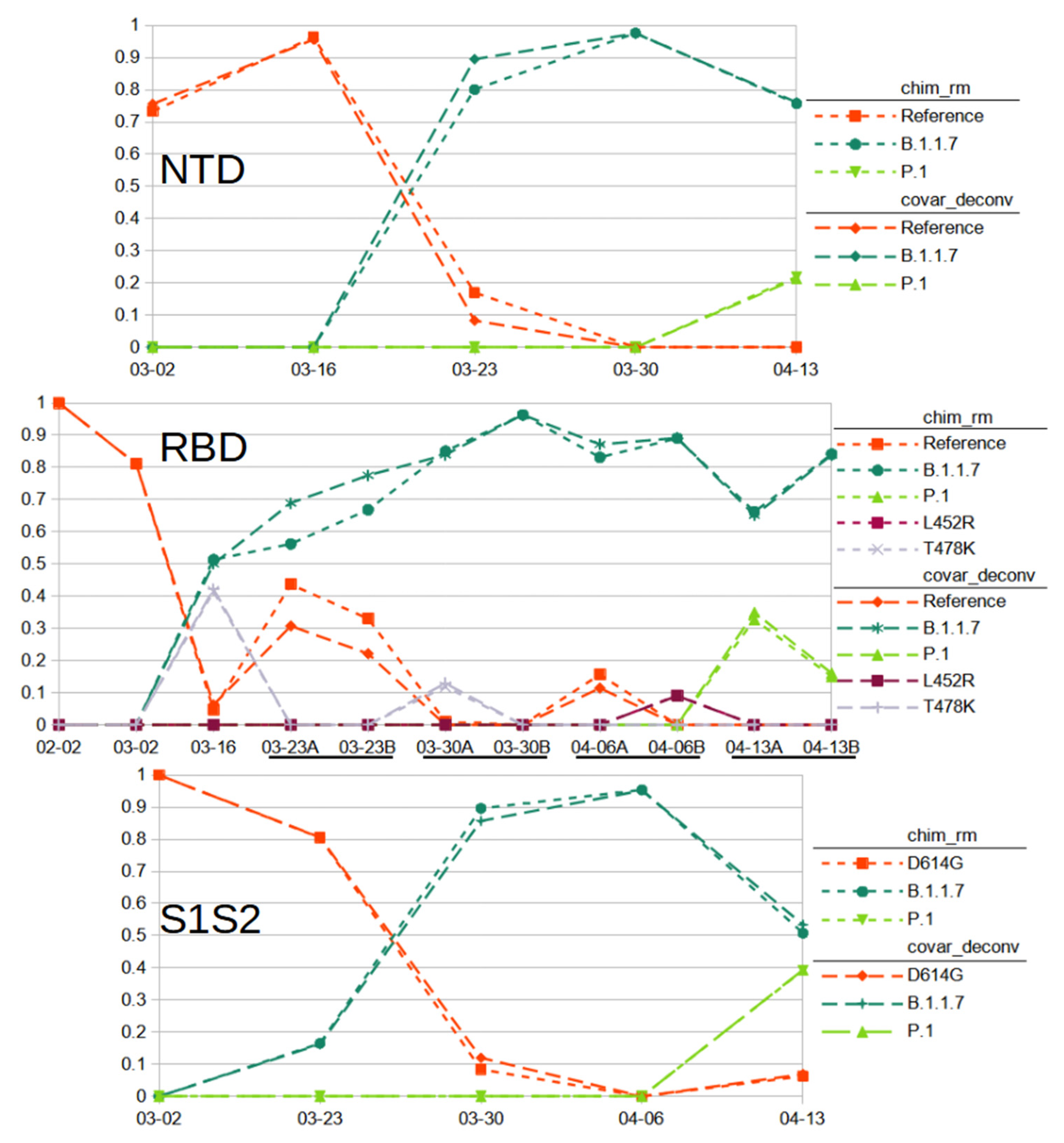
https://github.com/degregory/SR_manuscript/tree/master/Fenton_Data accessed on 23 June 2021. The two different chimera removal methods showed good concordance, validating each as being a viable detection method. Duplicate RT-PCR preparation and sequencing of the same wastewater sample also generally provided similar results, though less consistently (
Figure 5. Compare A and B RBD amplicon preparations). These differences were more pronounced with covariants with relatively low abundance, such as is seen with 3-30 RBD samples, where one detects T478K and the other does not (
Figure 5). These differences illustrate the stochastic nature of RT-PCR amplification.
We used the chimera removed and covariant deconvolution outputs to assign sequences to known variant lineages or the reference (
Supplementary 10, 11 and 12) based on polymorphisms present. Polymorphisms were considered for lineage assignment if they appeared in multiple sequencing runs or were known to be present in circulating populations reported to GSIAD (
https://www.gisaid.org/, accessed on 20 February 2021). Polymorphisms that could not be validated were not taken into account for lineage assignment. Based on these assignments, we were able to observe the changes to virus populations in the sewershed over time (
Figure 5). We classified the sequences found from the NTD amplicon as matching reference sequence, lineage B.1.1.7 (Alpha) with ‘203-208Del 429-431Del’ or lineage P.1 (Gamma) with ‘412T(D138Y) 570T(R190S)’ (
Supplementary 10). Sequences from the RBD amplicon matched reference sequence, lineages B.1.1.7 with ‘1501T(N501Y) 1709A(A570D)’, P.1 with ‘1250C(K417T) 1450A(E484K) 1501T(N501Y)’, or had the single variations of T478K or L452R (
Supplementary 11). T478K and L452R each have lineage associations. However, no other polymorphisms are associated with these in the RBD amplicons, nor were any polymorphisms present in the other amplicons that would indicate the presence of any associated lineages. While these SNPs could be the result of PCR error, it is more likely the associated lineages exist in the sewershed, but, due to stochastic effects, the other associated polymorphisms in the other amplicons were not detected. They could have also arisen in a reference background. As we cannot assign them to a known lineage with any certainty, we assigned them to their own category. Sequences from the S1S2 amplicon matched lineage B.1.1.7 with ‘1841G(D614G) 2042A(P681H) 2147T(T716I)’, lineage P.1 with ‘1841G(D614G) 1963T(H655Y) 2063T(A688V)’ or the B.1 lineage with only the now ubiquitous D614G variation (
Supplementary 12). The 03-23 S1S2 sample had a sequence ‘1841G(D614G) 2037G(N679K) 2063T(A688V)’. While A688V is associated with P.1, it does not appear in that context here. As that is the only sample where those covariant sequences were observed and the polymorphisms are not frequently reported in GISAID (outside of P.1 for A688V), we did not feel we could validate this sequence as a novel lineage and instead tentatively assigned it to the reference category. From these results, we can conclude that the SARS-CoV-2 population of this sewershed changed in March 2021 from almost exclusively the D614G B.1 lineage to mainly the B.1.1.7 lineage, with the introduction of P.1 early in April 2021. This general method is now being used to track SARS-CoV-2 variants in many Missouri sewersheds (
https://storymaps.arcgis.com/stories/f7f5492486114da6b5d6fdc07f81aacf accessed on 23 June 2021).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}