
Large-Scale International Validation of an Indirect ELISA Based on Recombinant Nucleocapsid Protein of Rift Valley Fever Virus for the Detection of IgG Antibody in Domestic Ruminants

 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Origin of Serum Specimens
2.2. VNT
2.3. rNP I- ELISA
2.4. Selection of Cut-Off Values
2.5. Determination of Diagnostic Accuracy and Other Statistical Analysis
2.6. Serum Inactivation
2.7. rNP I-ELISA Robustness
2.8. IgG-Sandwich RVFV ELISA
3. Results
3.1. VNT
3.2. Selection and Optimization of Cut-Off Values
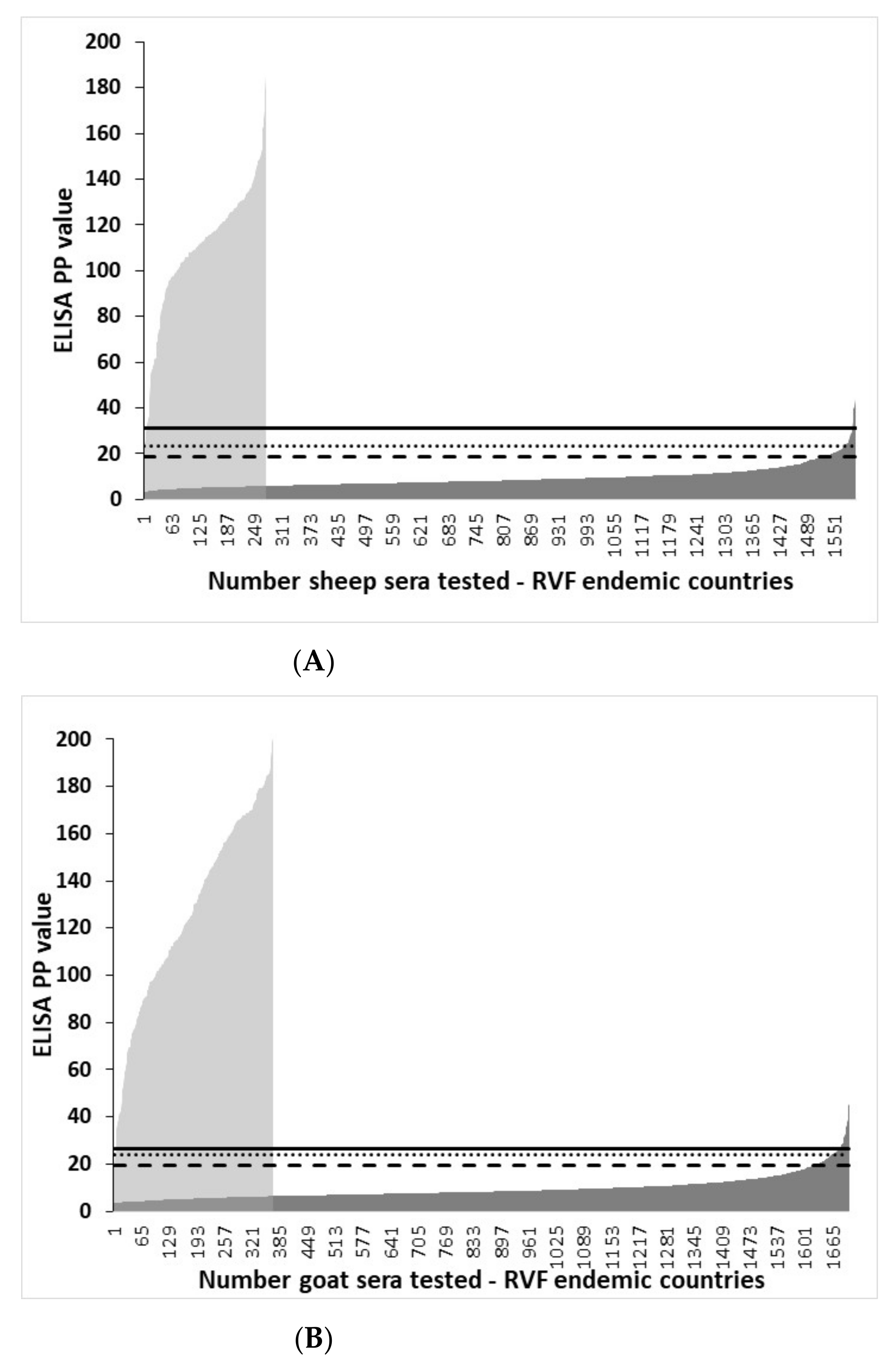
3.3. Distribution of rNP I-ELISA PP Values in Ruminant Sera from RVF-Endemic Countries and the Selection of Cut-Offs
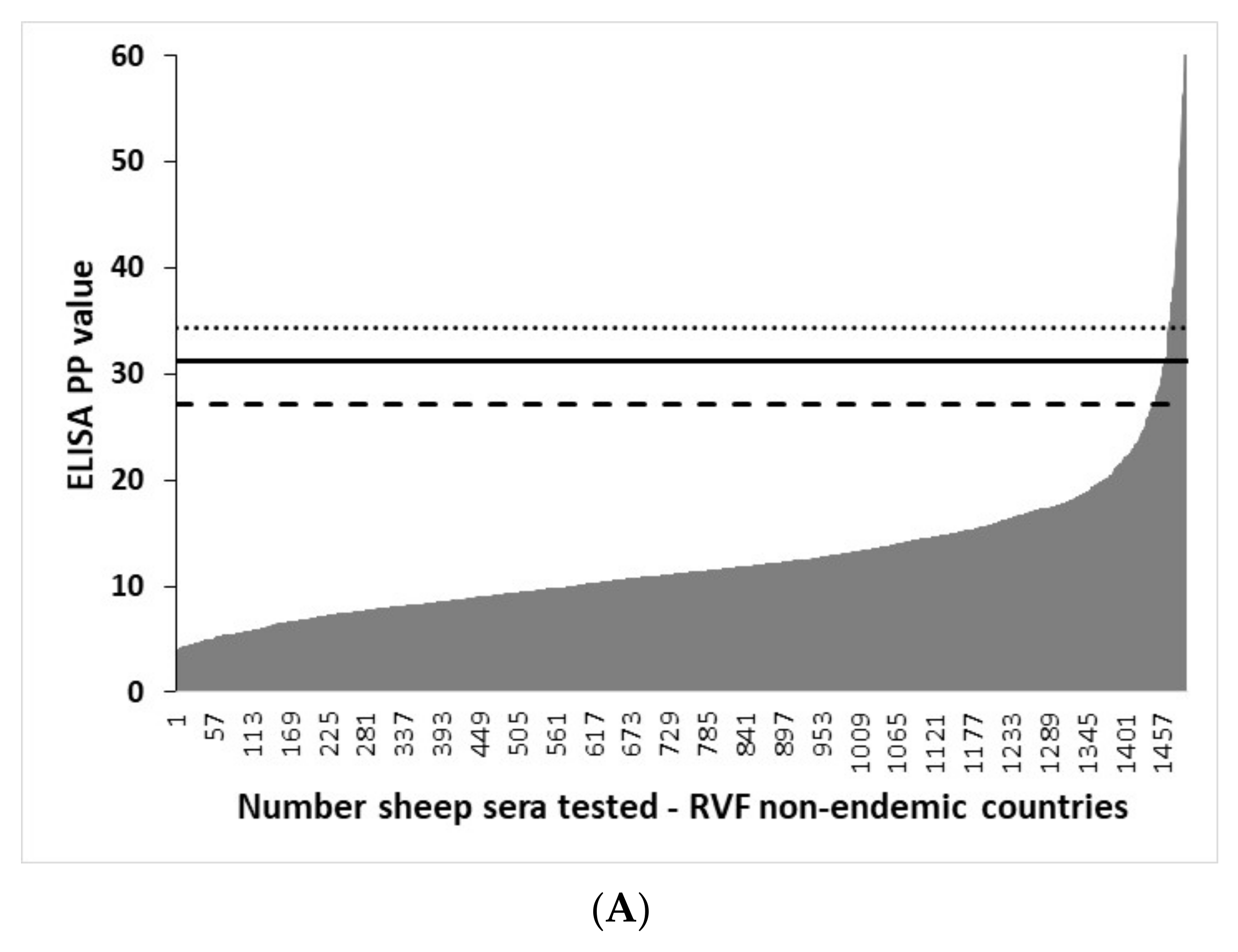
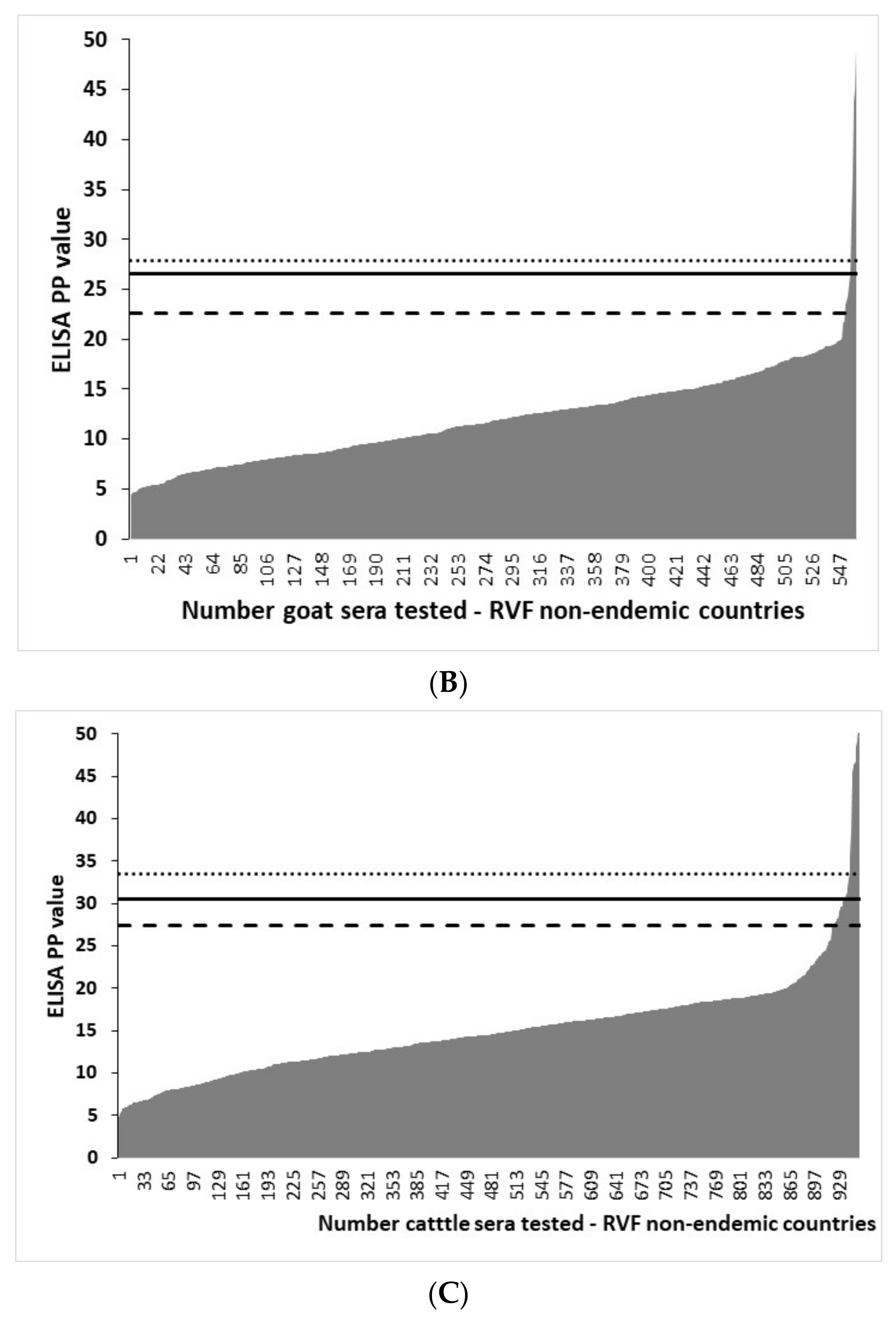
3.4. Distribution of rNP I-ELISA PP Values in Ruminant Sera from RVF-Free Countries and the Selection of Cut-Offs
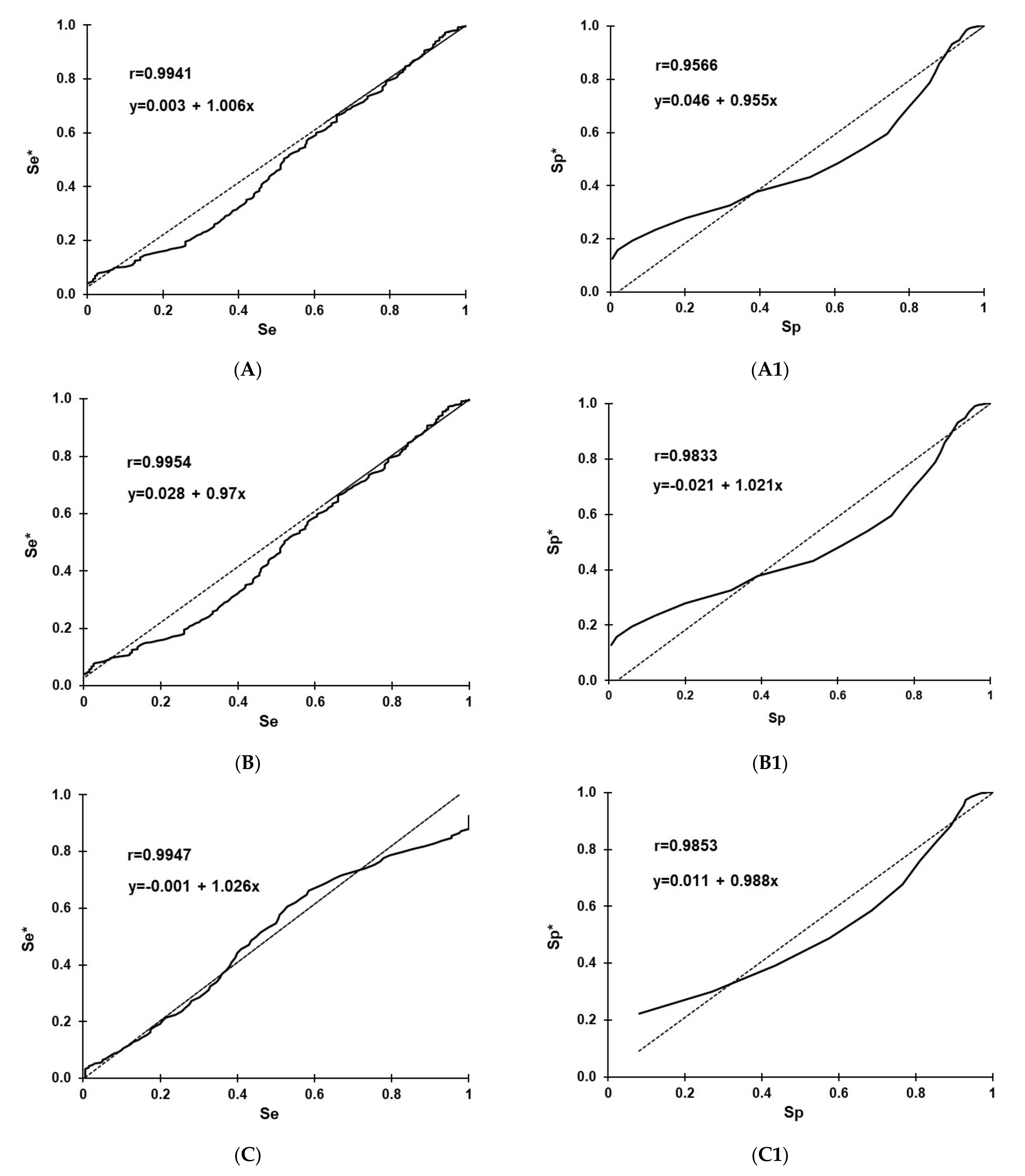
3.5. D-Se and D-Sp
3.6. The Effect of Inactivation
3.7. The Effect of Temperature Incubation
3.8. IgG-Sandwich RVFV ELISA
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bird, B.H.; Ksiazek, T.G.; Nichol, S.T.; Maclachlan, N.J. Rift Valley fever virus. J. Am. Vet. Med. Assoc. 2009, 234, 883–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevalier, V.; Pepin, M.; Plee, L.; Lancelot, R. Rift Valley fever—A threat for Europe? Eurosurveillance 2010, 15, 19506. [Google Scholar] [CrossRef] [PubMed]
- Paweska, J.T.; Jansen van Vuren, P. Rift Valley fever virus: A virus with potential for global emergence. In The Role of Animals in Emerging Viral Diseases; Johnson, N., Ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2013; pp. 169–200. ISBN 978-0-12-405191-1. [Google Scholar]
- McMillen, C.M.; Hartman, A.L. Rift Valley fever in animals and humans: Current perspectives. Antivir. Res. 2018, 156, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Mehand, M.S.; Al-Shorbaji, F.; Millett, P.; Murgue, B. The WHO R&D Blueprint: 2018 review of emerging infectious diseases requiring urgent research and development efforts. Antivir. Res. 2018, 159, 63–67. [Google Scholar] [PubMed]
- Swanepoel, R.; Coetzer, J.A.W. Rift Valley fever. In Infectious Diseases of Livestock with Special Reference to Southern Africa; Coetzer, J.A.W., Thomson, G.R., Tustin, R.C., Eds.; Oxford University Press: Cape Town, South Africa, 2004; pp. 688–717. [Google Scholar]
- Paweska, J.T. Rift Valley fever. In Clinical Case Study of Emerging Infectious Diseases; Ergonul, O., Can, F., Akova, A., Madoff, L., Eds.; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2014; pp. 73–93. ISBN 978-0-12-416975-3. [Google Scholar]
- Swanepoel, R.; Paweska, J.T. Rift Valley fever. In Oxford Textbook of Zoonoses: Biology, Clinical Practise, and Public Health Control, 2nd ed.; Palmer, S.R., Soulsby, L., Torgerson, P.R., Brown, D.W.G., Eds.; Oxford University Press: Oxford, UK, 2011; pp. 421–431. [Google Scholar]
- Paweska, J.T. Rift Valley fever. In New Developments in Major Vector-Borne Diseases—Part II: Important Diseases for Veterinarians; Zientara, S., Verwoerd, D., Pastoret, P.P., Eds.; International Office of Epizootics: Paris, France, 2015; Volume 34, pp. 375–389. [Google Scholar]
- McMillen, C.M.; Hartman, A.L. Rift Valley fever: A threat to pregnant women hiding in plain sight? J. Virol. 2021, 95, e01394-19. [Google Scholar] [CrossRef] [PubMed]
- Sidwell, R.W.; Smee, D.F. Viruses of the Bunya- and Togaviridae families; potential as bioterrorism agnets and means of control. Antivir. Res. 2003, 57, 101–111. [Google Scholar] [CrossRef]
- Lim, D.V.; Simpson, J.M.; Kearns, E.A.; Kramer, M.F. Current and developing technologies for monitoring agents of bioterrorism and biowarfare. Clin. Microbiol. 2005, 18, 583–607. [Google Scholar] [CrossRef] [Green Version]
- Lomly, S.; Horton, D.L.; Hernandez-Triana, L.L.M.; Johson, N.; Fooks, A.R.; Hewson, R. Rift Valley fever virus: Strategies for maintenance, survival and vertical transmission in mosquitoes. J. Gen. Virol. 2017, 98, 875–887. [Google Scholar] [CrossRef] [Green Version]
- Pepin, M.; Bouloy, M.; Bird, B.H.; Kemp, A.; Paweska, J. Rift Valley fever virus (Bunyaviridae: Phlebovirus): An update on pathogenesis, molecular epidemiology, vectors, diagnostics and prevention. Vet. Res. 2010, 41, 61. [Google Scholar] [CrossRef] [Green Version]
- Brett, N.A.; Thomas, J.; Weyer, J.; Cengimbo, A.; Essoya, L.D.; Jacobs, C.; Ntuli, S.; Modise, M.; Mathonsi, M.; Mashishi, M.S.; et al. Epidemiological investigations into outbreaks of Rift Valley fever in humans, South Africa, 2008–2201. Emerg. Infect. Dis. 2013, 19, 1918–1925. [Google Scholar]
- Msimang, V.; Thompson, P.N.; Jansen van Vuren, P.; Tempia, S.; Cordel, C.; Kgaladi, J.; Khosa, J.; Burt, F.J.; Liang, J.; Rostal, M.K.; et al. Rift Valley fever virus exposure amongst farmers, farm workers, and veterinary professionals in central South Africa. Viruses 2019, 11, 140. [Google Scholar] [CrossRef] [Green Version]
- LaBeaud, A.D.; Pfeil, S.; Muiruri, S.; Dahir, S.; Sutherland, L.J.; Traylor, Z.; Gildengorin, G.; Muchiri, E.M.; Morrill, J.; Peters, C.J.; et al. Factors associated with severe human Rift Valley fever in Sangailu, Garissa County, Kenya. PLoS Negl. Trop. Dis. 2015, 9, e0003548. [Google Scholar] [CrossRef] [Green Version]
- Maes, P.; Alkhovsky, S.V.; Bao, Y.; Beer, M.; Birkhead, M.; Briese, T.; Buchmeier, M.J.; Calisher, C.H.; Charrel, R.N.; Choi, I.R.; et al. Taxonomy of the family Arenaviridae and the order Bunyavirales: Update 2018. Arch. Virol. 2018, 163, 2295–2310. [Google Scholar] [CrossRef] [Green Version]
- Bouloy, M.; Weber, F. Molecular Biology of Rift Valley Fever Virus. Open Virol. J. 2010, 4, 8–14. [Google Scholar] [CrossRef]
- Maguranp, F.; Nicoletti, L. Humoral response in Toscana virus acute neurological diseases investigated by viral-protein-specific immunoassays. Clin. Diag. Lab. Immunol. 1999, 6, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.W., Jr.; Saluzo, J.F.; Ksiazek, T.G.; Smith, J.F.; Ennis, W.; Thuren, D.; Peters, C.J.; Digoutte, J.P. Comparison of in vitro and in vivo systems for propagation of Rift Valley fever virus from clinical specimens. Res. Virol. 1989, 140, 129–138. [Google Scholar] [CrossRef]
- Garcia, S.; Crance, J.M.; Billecocq, A.; Peinnequin, A.; Jouan, A.; Bouloy, M.; Garin, D. Quantitative real-time PCR detection of Rift Valley fever virus and its application to evaluation of antiviral compounds. J. Clin. Microbiol. 2001, 39, 4456–4461. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.S.; Turell, M.J.; Knauert, F.K.; Lofts, R.S. Detection of Rift Valley fever virus in mosquitoes by RT-PCR. Mol. Cell Probes. 1997, 11, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Sall, A.A.; Macondo, E.A.; Sene, O.K.; Diagane, M.; Sylla, R.; Mondo, M.; Girault, L.; Marrama, L.; Spiegel, A.; Diallo, M.; et al. Use of reverse transcriptase PCR in early diagnosis of Rift Valley fever. Clin. Diag. Lab. Immunol. 2002, 9, 713–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Njenga, K.M.; Paweska, J.; Wanjala, R.; Rao, C.Y.; Weiner, M.; Omballa, V.; Luman, E.T.; Mutonga, D.; Sharif, A.; Panning, M.; et al. Using field quantitative real-time reverse transcription-PCR test to rapidly identify highly viremic Rift Valley fever cases. J. Clin. Microbiol. 2009, 47, 1166–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drosten, C.; Götting, S.; Schilling, S.; Asper, M.; Panning, M.; Schmitz, H.; Günter, S. Rapid detection and quantification of RNA of Ebola and Marburg viruses, Lassa virus, Crimean-Congo hemorrhagic fever virus, Rift Valley fever virus, dengue virus, and yellow fever virus by real-time reverse transcription-PCR. J. Clin. Microbiol. 2002, 40, 2323–2330. [Google Scholar] [CrossRef] [Green Version]
- Wilson, W.C.; Romit, M.; Jasperson, D.C.; Weingartl, H.; Binepal, Y.S.; Maluleke, M.R.; Wallace, D.B.; Jansen van Vuren, P.; Paweska, J.T. Development of a Rift Valley fever real-time RT-PCR assay that can detect all three genome segments. J. Virol. Methods 2013, 193, 426–431. [Google Scholar] [CrossRef] [Green Version]
- Le Roux, C.A.; Kubo, T.; Grobbelaar, A.A.; Jansen van Vuren, P.; Weyer, J.; Nel, L.H.; Swanepoel, R.; Morita, K.; Paweska, J.T. Development and evaluation of a real-time reverse transcription-loop-mediated isothermal amplification assay for rapid detection of Rift Valley fever virus in clinical specimens. J. Clin. Microbiol. 2009, 47, 645–651. [Google Scholar] [CrossRef] [Green Version]
- Zaki, A.; Coudrier, D.; Yousef, A.I.; Fakeeh, M.; Bouloy, M.; Billecocq, A. Production of monoclonal antibodies against Rift Valley fever virus. Application for rapid diagnosis tests (virus detection and ELISA) in human sera. J. Virol. Methods 2006, 131, 34–40. [Google Scholar] [CrossRef]
- Jansen van Vuren, P.; Paweska, J.T. Laboratory safe detection of nucleocapsid protein of Rift Valley fever virus in human and animal specimens by a sandwich ELISA. J. Virol. Methods 2009, 157, 15–24. [Google Scholar] [CrossRef]
- Cêtre-Sossah, C.; Pe’darrieu, A.; Juremalm, M.; Jansen Van Vuren, P.; Brun, A.; Ould EL Mamy, A.B.; Heraud, J.M.; Filipone, C.; Ravalohery, J.P.; Chaabihi, H.; et al. Development and validation of a pen side test for Rift Valley fever. PLoS Negl. Trop. Dis. 2019, 13, e0007700. [Google Scholar] [CrossRef] [Green Version]
- Swanepoel, R.; Struthers, J.K.; Erasmus, M.J.; Shepherd, S.P.; McGillivray, G.M.; Erasmus, B.J.; Barnard, B.J. Comparison of techniques for demonstrating antibodies to Rift Valley fever virus. J. Hyg. 1986, 97, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Kitchen, S.F. Laboratory infections with the virus of Rift Valley fever. Am. J. Trop. Med. Hyg. 1934, 4, 547–564. [Google Scholar] [CrossRef]
- Smithburn, K.C.; Mahaffy, A.F.; Haddow, A.J.; Kitchen, S.F.; Smith, J.F. Rift Valley fever accidental infections among laboratory workers. J. Immunol. 1949, 62, 213–227. [Google Scholar] [PubMed]
- Wichgers Schreur, P.J.; Paweska, J.T.; Kant, J.; Kortekaas, J. A novel highly sensitive, rapid and safe Rift Valley fever virus neutralization test. J. Virol. Methods 2017, 10, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Paweska, J.T.; Barnard, B.J.H.; Williams, R. The use of sucrose-acetone-extracted Rift Valley fever virus antigen derived from cell-culture in an indirect enzyme-linked immunosorbent assay and haemagglutination-inhibition test. Onderstepoort J. Vet. Res. 1995, 62, 227–233. [Google Scholar] [PubMed]
- Paweska, J.T.; Smith, S.J.; Wright, I.M.; Williams, R.; Cohen, A.S.; van Dijk, A.A.; Grobbelaar, A.A.; Croft, J.E.; Swanepoel, R.; Gerdes, G.H. Indirect enzyme-linked immunosorbent assay for the detection of antibody against Rift Valley fever in domestic and wild ruminants. Onderstepoort J. Vet. Res. 2003, 70, 49–64. [Google Scholar] [PubMed]
- Paweska, J.T.; Burt, F.J.; Anthony, F.; Smith, S.J.; Grobbleaar, A.A.; Croft, J.E.; Ksiazek, T.; Swanepoel, R. IgG-sandwich and IgM-capture enzyme-linked immunosorbent assay for detection of antibody to Rift Valley fever in domestic ruminants. J. Virol. Methods 2003, 113, 103–112. [Google Scholar] [CrossRef]
- Paweska, J.T.; Burt, F.J.; Swanepoel, R. Validation of IgG-sandwich and IgM-capture ELISA for the detection of antibody to Rift Valley fever virus in humans. J. Virol. Methods 2005, 124, 173–181. [Google Scholar] [CrossRef]
- Paweska, J.T.; Mortimer, E.; Leman, P.A.; Swanepoel, R. An inhibition enzyme-linked immunosorbent assay for the detection of antibody to Rift Valley fever virus in humans, domestic and wild ruminants. J. Virol. Methods 2005, 127, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Pepin, A.; Paweska, J.; Bouloy, M. Diagnostic specificity of ELISA-based tests for the detection of antibodies to Rift Valley fever virus in French ruminants. Rev. Méd. Vét. 2010, 161, 104–107. [Google Scholar]
- Sobarzo, A.; Paweska, J.T.; Herrman, S.; Amir, T.; Marks, R.S.; Lobel, L. Optical fiber immunosensor for the detection of IgG antibody to Rift Valley fever virus in humans. J. Virol. Methods 2007, 146, 327–334. [Google Scholar] [CrossRef]
- Fafetine, J.M.; Tijhaar, E.; Paweska, J.T.; Neves, L.C.B.G.; Hendriks, J.; Swanepoel, R.; Coetzer, J.A.W.; Egbering, H.F.; Rutten, V.P.M.G. Cloning and expression of Rift Valley fever virus nucleocapsid (N) protein and evaluation of a N-protein based indirect ELISA for the detection of specific IgG and IgM antibodies in domestic ruminants. Vet. Microbiol. 2007, 121, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Jansen van Vuren, P.; Potgieter, A.C.; Paweska, J.T.; van Dijk, A.A. Preparation and evaluation of a recombinant Rift Valley fever virus N protein for the detection of IgG and IgM antibodies in humans and animals by indirect ELISA. J. Virol. Methods 2007, 140, 106–114. [Google Scholar] [CrossRef]
- Jäckel, S.; Eiden, M.; Balkema-Buschmann, A.; Ziller, M.; Jansen van Vuren, P.; Paweska, J.T.; Groschup, M.H. A novel indirect ELISA based on glycoprotein Gn for the detection of IgG antibodies against Rift Valley fever virus in small ruminants. Res. Vet. Sci. 2013, 95, 725–730. [Google Scholar] [CrossRef]
- Faburay, B.; Wilson, W.C.; Secka, A.; Drolet, B.; McVey, D.S.; Richt, J.A. Evaluation of an indirect enzyme-linked immunosorbent assay based on recombinant baculovirus-expressed Rift Valley fever virus nucleoprotein as the diagnostic antigen. J. Clin. Microbiol. 2019, 57, e01058-19. [Google Scholar] [CrossRef] [Green Version]
- McElroy, A.K.; Albarino, C.G.; Nichol, S.T. Development of a RVFV ELISA that can distinguish infected from vaccinated animals. Virol. J. 2009, 6, 125. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.M.; Wilson, W.C.; Faburay, B.; Richt, J.; McVey, D.S.; Rowland, R.R. Multiplex detection of IgG and IgM to Rift Valley fever virus nucleoprotein, nonstructural proteins, and glycoprotein in ruminants. Vector Borne Zoonotic Dis. 2016, 16, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, J.F.; Ragan, I.K.; Rowland, R.R.; Wainaina, M.; Mbotha, D.; Wilson, W. A multiplex fluorescence microsphere immunoassay for increased understanding of Rift Valley fever immune responses in ruminants in Kenya. J. Virol. Methods 2019, 269, 70–76. [Google Scholar] [CrossRef]
- Cêtre-Sossah, C.; Billecocq, A.; Lancelot, R.; Defernez, C.; Favre, J.; Bouloy, M.; Martinez, D.; Albina, E. Evaluation of a commercial competitive ELISA for the detection of antibodies to Rift Valley fever virus in sera of domestic ruminants in France. Prevent. Vet. Med. 2009, 90, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Nah, J.-J.; Moon, J.-S.; Ko, Y.-J.; Yoo, H.-S.; Kweon, C.-H. Competitive ELISA for the detection of antibodies to Rift Valley fever virus in goats and cattle. J. Vet. Med. Sci. 2012, 74, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Upreti, D.; Cernixcchiaro, N.; Richt, J.A.; Wilson, W.C.; Clavijo, A.; Davis, A.S. Preliminary evaluation of diagnostic accuracy and precision of a competitive ELISA for detection of antibodies to Rift Valley fever virus in cattle and sheep sera. J. Virol. Methods 2018, 262, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Paweska, J.T.; Jansen van Vuren, P.; Kemp, A.; Swanepoel, R.; Buss, P.; Roy, G.; Bengis, R.G.; Gakuya, F.; Breiman, R.F.; Kariuki, N.M. A recombinant nucleocapsid-based indirect ELISA for serodiagnosis of Rift Valley fever in African wildlife. In Proceedings of the Sustainable Improvement of Animal Production and Health; Odongo, N.E., Garcia, M., Viljoen, G.J., Eds.; FAO: Rome, Italy, 2010; pp. 309–312. ISBN 978-92-5-106697-3. [Google Scholar]
- Williams, R.; Ellis, C.E.; Smith, S.J.; Potgieter, C.A.; Wallace, D.; Mareledwane, V.E.; Majiwa, P.A.O. Validation of an IgM antibody capture ELISA based on a recombinant nucleoprotein for identification of domestic ruminants infected with Rift Valley fever virus. J. Virol. Methods 2011, 177, 140–146. [Google Scholar] [CrossRef]
- Fafetine, J.M.; Jansen van Vuren, P.; Paweska, J.T. Comparison of a recombinant nucleocapsid IgG indirect ELISA with an IgG sandwich ELISA for the detection of antibodies to Rift Valley fever virus in small ruminants. Vector Borne Zoonotic Dis. 2012, 12, 1062–1064. [Google Scholar] [CrossRef]
- Ellis, C.E.; Mareledwane, V.E.; Williams, R.; Wallace, D.; Majiwa, P.A.O. Validation of an ELISA for the concurent detection of total antibodies (IgM and IgG) to Rift Valley fever virus. Onderstepoort J. Vet. Res. 2014. Art. #675.. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Adungo, F.; Konongoi, S.L.; Inoue, S.; Sang, R.; Ashur, S.; Kwallah, A.O.; Uchida, L.; Buerano, C.C.; Mwau, M.; et al. Comparison of enzyme-linked immunosorbent assay systems using Rift Valley fever virus nucleocapsid protein and inactivated virus as antigens. Virol. J. 2018, 15, 178. [Google Scholar] [CrossRef]
- Paweska, J.T.; Jansen van Vuren, P.; Swanepoel, R. Validation of an indirect ELISA based on a recombinant nucleocapsid protein of Rift Valley fever virus for the detection of IgG antibody in humans. J. Virol. Methods 2007, 146, 119–124. [Google Scholar] [CrossRef]
- Paweska, J.T.; Jansen van Vuren, P.; Kemp, A.; Buss, P.; Bengis, R.G.; Gakuya, F.; Breiman, R.F.; Kariuki, M.N.; Swanepoel, R. Recombinant nucleocapsid-based ELISA for detection of IgG antibody to Rift Valley fever virus in African buffalo. Vet Microbiol. 2008, 127, 21–28. [Google Scholar] [CrossRef]
- De Bronsvoort, B.M.C.; Bagninbom, J.-M.; Ndip, L.; Kelly, R.F.; Handel, I.; Tanya, V.N.; Morgan, K.L.; Ngwa, V.N.; Mazeri, S.; Nfon, C. Comparison of two Rift Valley fever serological tests in Cameroonian cattle populations using a Bayesian latent class approach. Frontiers. Vet. Sci. 2019, 6, 258. [Google Scholar]
- Wright, P.F.; Nilsson, E.; Van Rooij, E.M.A.; Lelenta, M.; Jeggo, M.H. Standardization and validation of enzyme-linked immunosorbent assay techniques for the detection of antibody in infectious disease diagnosis. Rev. Sci. Tech. Off. Int. Epiz. 1993, 12, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, R.H. Validation of serological assays for diagnosis of infectious diseases. Rev. Sci. Tech. Off. Int. Epiz. 1998, 17, 469–486. [Google Scholar] [CrossRef]
- Greiner, M.; Gardner, I.A. Epidemiological issues in the validation of veterinary diagnostic test. Prev. Vet. Med. 2000, 45, 3–22. [Google Scholar] [CrossRef]
- Greiner, M.; Gardner, I.A. Application of diagnostic tests in veterinary epidemiological studies. Prev. Vet. Med. 2000, 45, 43–59. [Google Scholar] [CrossRef]
- Greiner, M. Two-graph receiver operating characteristic (TG-ROC): Update version supports optimisation of cut-off values that minimize overall misclassification costs. J. Virol. Methods. 1996, 191, 93–94. [Google Scholar] [CrossRef]
- Greiner, M. Two-graph receiver operating characteristic (TG-ROC): A Microsoft-EXCEL template for the selection of cut-off values in diagnostic tests. J. Immunol. Methods. 1995, 185, 145–146. [Google Scholar] [CrossRef]
- Greiner, M.; Sohr, D.; Göbel, L. A modified ROC analysis for the selection of cut-off values and the definition of intermediate results of serodiagnostic tests. J. Virol. Methods. 1995, 185, 113–132. [Google Scholar] [CrossRef]
- Duncan, D. Multiple Range and Multiple F Tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Mitchell, S.W.; McCormick, J.B. Physicochemical inactivation of Lassa, Ebola and Marburg viruses and effect on clinical laboratory analyses. J. Clin. Microbiol. 1984, 20, 486–489. [Google Scholar] [CrossRef] [Green Version]
- Hadddock, E.; Feldman, F.; Feldman, H. Effective chemical inactivation of Ebola virus. Emerg. Infect. Dis. 2016, 22, 1292–1294. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K.L.; Banyard, A.C.; McElhinney, L.; Johson, N.; Horton, D.L.; Hernández-Triana, L.M.; Fooks, A.R. Rift Valley fever virus: A review of diagnosis and vaccination, and implications for emergence in Europe. Vaccine 2015, 33, 5520–5531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, V.; Kristiansen, P.; Norheim, G.; Yimer, S.A. Rift valley fever: Diagnostic challenges and investment needs for vaccine development. BMJ Glob. Health 2020, 5, e002694. [Google Scholar] [CrossRef]
- Caporale, V.; Nannini, D.; Ricci, L. Quality assurance in veterinary diagnostic laboratories. Rev. Sci. Tech. Off. Int. Epiz. 1998, 17, 459–468. [Google Scholar] [CrossRef] [Green Version]
- Nanini, D.; Giovannini, A.; Fiore, G.L.; Marabelli, R.; Caporale, V. Quality assurance in veterinary diagnostic laboratories. Rev. Sci. Tech. Off. Int. Epiz. 1999, 17, 571–584. [Google Scholar]
- Taverniers, I.; De Loose, M.; Van Bockstaele, E. Trends in quality in the analytical laboratory. II. Analytical method validation and quality assurance. Trends Anal. Chem. 2004, 23, 535–552. [Google Scholar] [CrossRef]
- Crowther, J.R. ELISA theory and practice. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NY, USA, 1995. [Google Scholar]
- Grobbelaar, A.A.; Weyer, J.; Leman, P.A.; Kemp, A.; Paweska, J.T.; Swanepoel, R. Molecular epidemiology of Rift Valley fever virus. Emerg. Infec. Dis. 2011, 17, 2270–2276. [Google Scholar] [CrossRef]
- Swanapeol, R.; Struthers, J.K.; Erasmus, M.J.; Shepherd, S.P.; McGollivray, G.M.; Shepherd, A.J.; Hummitzsch, D.E.; Erasmus, B.J.; Barnards, B.J. Comparative pathogenicity and antigenic cross-reactivity of Rift Valley fever and other African phleboviruses in sheep. J. Hyg. Camp. 1986, 97, 331–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizard, A.L.; Anderson, G.A.; Gasser, R.B. Determination of the optimum cut-off value of a diagnostic test. Prev. Vet. Med. 1990, 10, 137–143. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Ovine | Total | Caprine | Total | Bovine | Total | |||
|---|---|---|---|---|---|---|---|---|---|
| VNT+ 1 | VNT− 2 | Tested | VNT+ | VNT− | Tested | VNT+ | VNT− | Tested | |
| RVF-endemic | |||||||||
| Burkina Faso | 20 | 165 | 185 | 0 | 268 | 268 | 72 | 924 | 996 |
| DRC | 1 | 61 | 62 | 0 | 0 | 0 | 133 | 813 | 946 |
| Mozambique | 170 | 435 | 605 | 171 | 717 | 888 | 0 | 0 | 0 |
| Senegal | 16 | 234 | 250 | 2 | 76 | 78 | 58 | 94 | 152 |
| Uganda | 0 | 0 | 0 | 170 | 527 | 697 | 60 | 623 | 683 |
| Yemen | 68 | 704 | 772 | 26 | 115 | 141 | 33 | 54 | 87 |
| Sub-total | 275 | 1599 | 1874 | 369 | 1703 | 2072 | 356 | 2508 | 2864 |
| RVF-free | |||||||||
| France | 0 | 720 | 720 | 0 | 560 | 560 | 0 | 640 | 640 |
| Poland | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 292 | 292 |
| USA | 0 | 773 | 773 | 0 | 0 | 0 | 0 | 23 | 23 |
| Sub-total | 0 | 1493 | 1493 | 0 | 560 | 560 | 0 | 955 | 955 |
| Total | 275 | 3092 | 3367 | 369 | 2263 | 2632 | 356 | 3463 | 3819 |
| Animal Species | Cut-Off PP Value 1 | FN 2/TP 3 | D-Se 4 (95% CI) 5 | Ef 6 | Y 7 | PPV 8 (%) | NPV 9 (%) |
|---|---|---|---|---|---|---|---|
| Sheep | |||||||
| Mean + 2SD 10 | 18.63 | 1/274 | 99.6 (98–100) | 0.964 | 0.953 | 83.80 | 99.91 |
| Mean + 3SD 11 | 23.30 | 2/273 | 99.3 (97.4–99.9) | 0.963 | 0.976 | 91.75 | 99.86 |
| TG-ROC 12 | 31.23 | 7/268 | 97.5 (94.8–99) | 0.960 | 0.970 | 97.12 | 99.57 |
| Goats | |||||||
| Mean + 2SD | 19.28 | 0/369 | 100 (99–100) | 0.967 | 0.952 | 86.82 | 100.00 |
| Mean + 3SD | 24.17 | 0/369 | 100 (99–100) | 0.966 | 0.978 | 92.62 | 100.00 |
| TG-ROC | 26.57 | 0/369 | 100 (99–100) | 0.966 | 0.988 | 95.63 | 100.00 |
| Cattle | |||||||
| Mean + 2SD | 24.38 | 15/341 | 95.8 (93.1–97.6) | 0.957 | 0.921 | 88.65 | 98.70 |
| Mean + 3SD | 30.86 | 33/323 | 90.7 (87.2–93.5) | 0.948 | 0.894 | 94.22 | 97.85 |
| TG-ROC | 30.46 | 33/323 | 90.7 (87.2–93.5) | 0.947 | 0.893 | 93.86 | 97.82 |
| Species/Origin | No. VNT 1 | Cut-Off PP 2 | FP 3/TN 4 | Mean + 2SD 5 D-Sp 6 (95%CI) 7 | Cut-Off PP | FP/TN | Mean + 3SD 8 D-Sp (95% CI) | Cut-Off PP | FP/TN | TG-ROC 9 D-Sp (95% CI) |
|---|---|---|---|---|---|---|---|---|---|---|
| Sheep serum panels | ||||||||||
| Burkina Faso | 165 | 16.8 | 12/153 | 92.7 (87.6–96.2) | 20.6 | 4/161 | 97.6 (93.9–99.3) | 31.23 | 0/165 | 100 (97.8–100) |
| DRC | 61 | 11.29 | 2/59 | 96.7 (88.7–99.6) | 13.63 | 1/60 | 98.4 (91.2–100) | 31.23 | 0/61 | 100 (94.1–100) |
| Mozambique | 435 | 20.74 | 16/419 | 96.3 (94.1–97.9) | 26.54 | 9/426 | 97.9 (96.1–99) | 31.23 | 6/429 | 98.6 (97–99.5) |
| Senegal | 234 | 21.15 | 8/226 | 96.6 (93.4–98.5) | 26.17 | 4/230 | 98.3 (95.7–99.5 | 31.23 | 1/233 | 96.6 (97.6–100) |
| Yemen | 704 | 16.72 | 40/664 | 94.3 (92.3–95.9) | 20.57 | 17/687 | 97.6 (96.2–98.6) | 31.23 | 1/703 | 99.9 (99.2–100) |
| France | 720 | 24.01 | 17/703 | 97.6 | 29.29 | 1o/710 | 98.6 | 31.23 | 8/712 | 98.9 |
| USA | 773 | 29.19 | 31/742 | 96.0 | 37.84 | 19/754 | 97.5 | 31.23 | 27/746 | 96.5 |
| Total RVF-endemic panels | 1599 | 18.63 | 69/1530 | 95.7 (94.6–96.6) | 23.30 | 27/1572 | 98.3 (97.6–98.9) | 31.23 | 8/1591 | 99.5 (99–99.8) |
| Total RVF-free panels | 1493 | 27.17 | 52/1441 | 96.5 | 34.43 | 29/1464 | 98.1 | 31.23 | 35/1458 | 97.7 |
| Total all panels | 3092 | 23.48 | 103/2989 | 96.7 (96–97.3) | 29.77 | 49/3043 | 98.4 (97.9–98.8 | 31.23 | 43/3049 | 98.6 (98.1–99) |
| Goat serum panels | ||||||||||
| Burkina Faso | 268 | 17.37 | 13/255 | 95.2 | 21.79 | 6/262 | 97.8 | 26.57 | 0/268 | 100 (98.8–100) |
| Mozambique | 717 | 17.81 | 37/680 | 94.8 (93–96.3) | 22.3 | 16/701 | 97.8 (96.4–98.7) | 26.57 | 7/710 | 99 (98–99.6) |
| Senegal | 76 | 27.18 | 4/72 | 94.7 (87.1–98.5) | 34.16 | 2/74 | 97.4 (90.8–99.7) | 26.57 | 4/72 | 94.7 (87.1–98.5) |
| Uganda | 527 | 20.02 | 28/499 | 94.7 (92.4–96.4) | 24.93 | 12/515 | 97.7 (96.1–98.8) | 26.57 | 8/519 | 98.5 (97–99.3) |
| Yemen | 115 | 20.03 | 5/110 | 95.7 (90.1–98.6) | 24.75 | 3/112 | 97.4 (92.6–99.5) | 26.57 | 2/113 | 98.3 (93.9–99.8) |
| France | 560 | 22.66 | 9/551 | 98.4 | 27.91 | 5/555 | 99.1 | 26.57 | 5/555 | 99.1 |
| Total RVF-endemic panels | 1703 | 19.28 | 82/1621 | 95.2 (94.1–96.2) | 24.17 | 37/1666 | 97.8 (97–98.5) | 26.57 | 21/1682 | 98.8 (98.1–99.2) |
| Total RVF-free panels | 560 | 22.66 | 9/551 | 98.4 | 27.91 | 5/555 | 99.1 | 26.57 | 5/555 | 98.1 |
| Total all panels | 2263 | 20.39 | 80/2183 | 96.5 (95.6–97.2) | 25.49 | 35/2228 | 98.5 (97.9–98.9) | 26.57 | 26/2237 | 98.9 (98.3–99.2) |
| Cattle serum panels | ||||||||||
| Burkina Faso | 924 | 25.23 | 35/889 | 96.2 (94.8–97.3) | 32.38 | 16/908 | 98.3 (97.2–99) | 30.46 | 18/906 | 98.1 (96.9–98.8) |
| DRC | 813 | 22.58 | 32/781 | 96.1 (94.5–97.3) | 28.45 | 9/804 | 98.9 (97.9–99.5) | 30.46 | 7/806 | 99.1 (98.2–99.7) |
| Senegal | 94 | 33.39 | 3/91 | 96.8 (91–99.3) | 41.56 | 3/91 | 96.8 (91–99.3) | 30.46 | 6/88 | 93.6 (86.6–97.6) |
| Uganda | 623 | 22.96 | 31/592 | 95 (93–96.6) | 28.46 | 9/614 | 98.6 (97.3–99.3) | 30.46 | 4/619 | 99.4 (98.4–99.8) |
| Yemen | 54 | 22.74 | 2/52 | 96.3 (87.3–99.5) | 28.14 | 1/53 | 98.1 (90.1–100) | 30.46 | 0/54 | 100 (93.4–100) |
| France | 640 | 23.35 | 12/628 | 98.1 | 27.68 | 5/635 | 99.2 | 30.46 | 3/637 | 99.5 |
| Poland | 292 | 34.44 | 12/280 | 95.9 | 43.41 | 11/281 | 96.2 | 30.46 | 17/275 | 94.2 |
| USA | 23 | 15.42 | 1/22 | 95.7 | 17.47 | 0/23 | 100 | 30.46 | 0/23 | 100 |
| Total RVF-endemic panels | 2508 | 24.38 | 94/2414 | 96.3 (95.4–97) | 30.86 | 32/2476 | 98.7 (98.2–99.1) | 30.46 | 35/2473 | 98.6 (98.1–99) |
| Total RVF-free panels | 955 | 27.53 | 32/923 | 96.7 | 33.71 | 13/942 | 98.6 | 30.46 | 20/935 | 97.9 |
| Total all panels | 3463 | 25.68 | 118/3345 | 96.6 (95.9–97.2) | 32.30 | 45/3418 | 98.7 (98.3–99.1) | 30.46 | 55/3408 | 98.4 (97.9–98.8) |
| Species/Origin | Shapiro-Wilk Test 1 | Dunn’s Test 2 | Mean/Median PP 3 | SD 4/(IQR) 5 PP | Range PP |
|---|---|---|---|---|---|
| VNT-negative sheep sera | |||||
| RVF-endemic countries | <0.001 | <0.001 | 9.3/8.2 | 4.7/(6.4–10.6) | 1.8–49.2 |
| RVF-free countries | <0.001 | 12.6/11.3 | 7.3/(8.4–14.7) | 3.9–75.8 | |
| VNT-negative goat sera | |||||
| RVF-endemic countries | <0.001 | <0.001 | 9.5/8.2 | 4.9/(6.6–10.8) | 2.3–45.2 |
| RVF-free countries | <0.001 | 12.2/11.8 | 5.2/(8.5–14.8) | 4.4–69.2 | |
| VNT-negative bovine sera | |||||
| RVF-endemic countries | <0.001 | <0.001 | 11.4/9.7 | 6.5/(7.1–13.8) | 3.3–76.0 |
| RVF-free countries | <0.001 | 15.2/14.5 | 6.2/(11.5–17.8) | 4.9–68.0 |
| Assay (Cut-Off PP Value) 1 | Mean log10 Serum Titre 2/Dose Response Curve R Square 3 | |||
|---|---|---|---|---|
| Untreated | 60° 1 h | 0.5%Tween 20 15 min 60° | 0.5%Triton X-100 15 min 60° | |
| Ovine (31.23) | 3.1/0.9665 | 3.5/0.9603 | 3.5/0.9625 | 3.5/0.9651 |
| Caprine (26.57) | 3.1/0.9572 | 3.5/0.9584 | 3.5/0.9590 | 3.5/0.9603 |
| Bovine (30.46) | 3.1/0.9638 | 3.2/0.9607 | 3.2/0.9625 | 3.2/0.9640 |
| Incubation Conditions | 4 °C/37 °C 1 | 37°C/37 °C 2 | RT/RT 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Serum 4 | Mean PP 5 | SD 6 | CV 7 | Mean PP | SD | CV | Mean PP | SD | CV (%) |
| S1 | 130.96 | 2.09 | 1.60 | 125.09 | 1.23 | 1.01 | 132.51 | 1.63 | 1.11 |
| S2 | 122.95 | 3.02 | 2.45 | 121.25 | 2.65 | 2.18 | 123.80 | 1.12 | 0.91 |
| S3 | 62.70 | 2.63 | 4.20 | 59.64 | 1.54 | 2.76 | 65.65 | 1.30 | 1.98 |
| S4 | 13.50 | 0.85 | 5.47 | 10.26 | 0.45 | 4.43 | 12.57 | 0.76 | 6.03 |
| G1 | 180.70 | 2.13 | 1.18 | 175.94 | 0.41 | 0.25 | 180.08 | 1.37 | 0.76 |
| G2 | 117.35 | 0.91 | 0.77 | 108.77 | 1.09 | 1.00 | 111.50 | 2.66 | 2.38 |
| G3 | 73.11 | 1.79 | 2.44 | 73.89 | 1.13 | 1.53 | 70.60 | 1.58 | 2.24 |
| G4 | 12.46 | 0.73 | 5.86 | 8.83 | 0.23 | 2.60 | 10.62 | 0.30 | 2.79 |
| C1 | 132.56 | 2.72 | 2.05 | 126.96 | 2.63 | 1.92 | 135.68 | 1.63 | 1.19 |
| C2 | 117.29 | 0.76 | 0.64 | 116.37 | 3.13 | 2.64 | 120.48 | 1.10 | 0.92 |
| C3 | 44.06 | 0.46 | 1.04 | 44.02 | 2.01 | 4.56 | 43.52 | 2.12 | 4.87 |
| C4 | 15.27 | 0.55 | 3.63 | 12.87 | 0.33 | 3.03 | 15.40 | 0.51 | 3.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pawęska, J.T.; Jansen van Vuren, P.; Msimang, V.; Lô, M.M.; Thiongane, Y.; Mulumba-Mfumu, L.K.; Mansoor, A.; Fafetine, J.M.; Magona, J.W.; Boussini, H.; et al. Large-Scale International Validation of an Indirect ELISA Based on Recombinant Nucleocapsid Protein of Rift Valley Fever Virus for the Detection of IgG Antibody in Domestic Ruminants. Viruses 2021, 13, 1651. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081651
Pawęska JT, Jansen van Vuren P, Msimang V, Lô MM, Thiongane Y, Mulumba-Mfumu LK, Mansoor A, Fafetine JM, Magona JW, Boussini H, et al. Large-Scale International Validation of an Indirect ELISA Based on Recombinant Nucleocapsid Protein of Rift Valley Fever Virus for the Detection of IgG Antibody in Domestic Ruminants. Viruses. 2021; 13(8):1651. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081651
Chicago/Turabian StylePawęska, Janusz T., Petrus Jansen van Vuren, Veerle Msimang, Modu Moustapha Lô, Yaya Thiongane, Leopold K. Mulumba-Mfumu, Alqadasi Mansoor, José M. Fafetine, Joseph W. Magona, Hiver Boussini, and et al. 2021. "Large-Scale International Validation of an Indirect ELISA Based on Recombinant Nucleocapsid Protein of Rift Valley Fever Virus for the Detection of IgG Antibody in Domestic Ruminants" Viruses 13, no. 8: 1651. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081651