
Co-Circulation of Phleboviruses and Leishmania Parasites in Sand Flies from a Single Site in Italy Monitored between 2017 and 2020

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
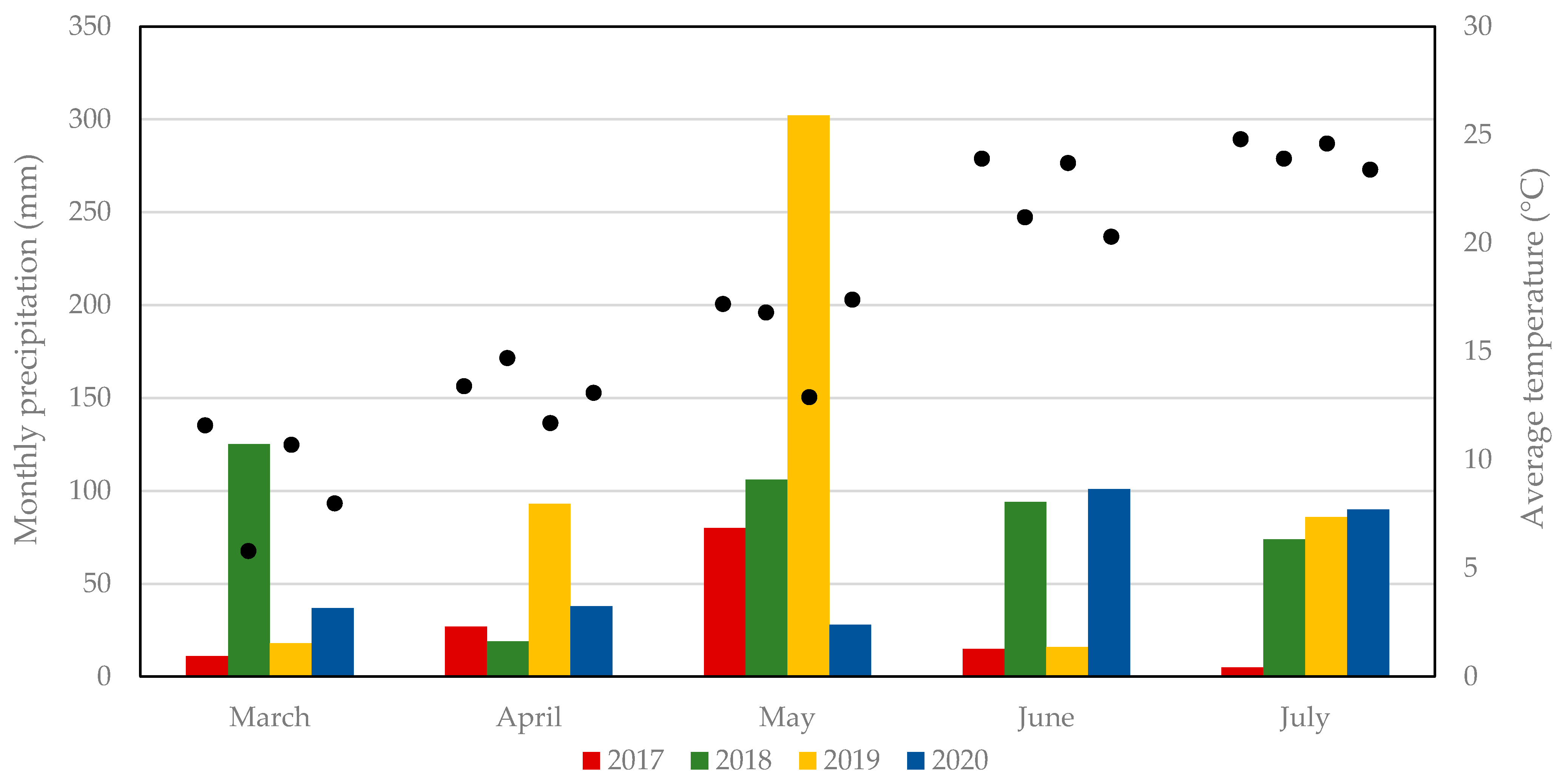
2.1. Sampled Site
2.2. Sand Fly Sampling and Identification
2.3. Pathogen Detection and Sequencing
2.4. Virus Isolation
3. Results
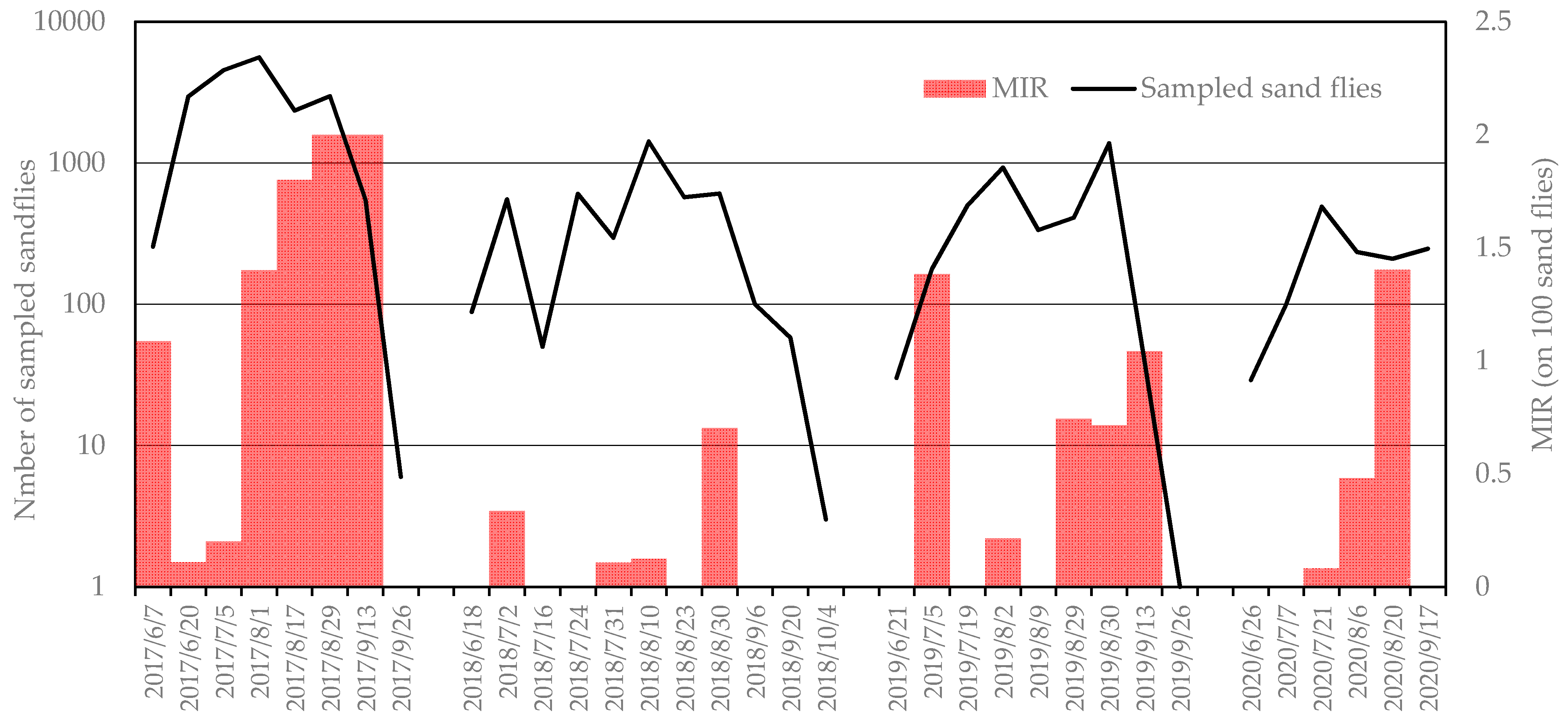
3.1. Sampled Sand Flies
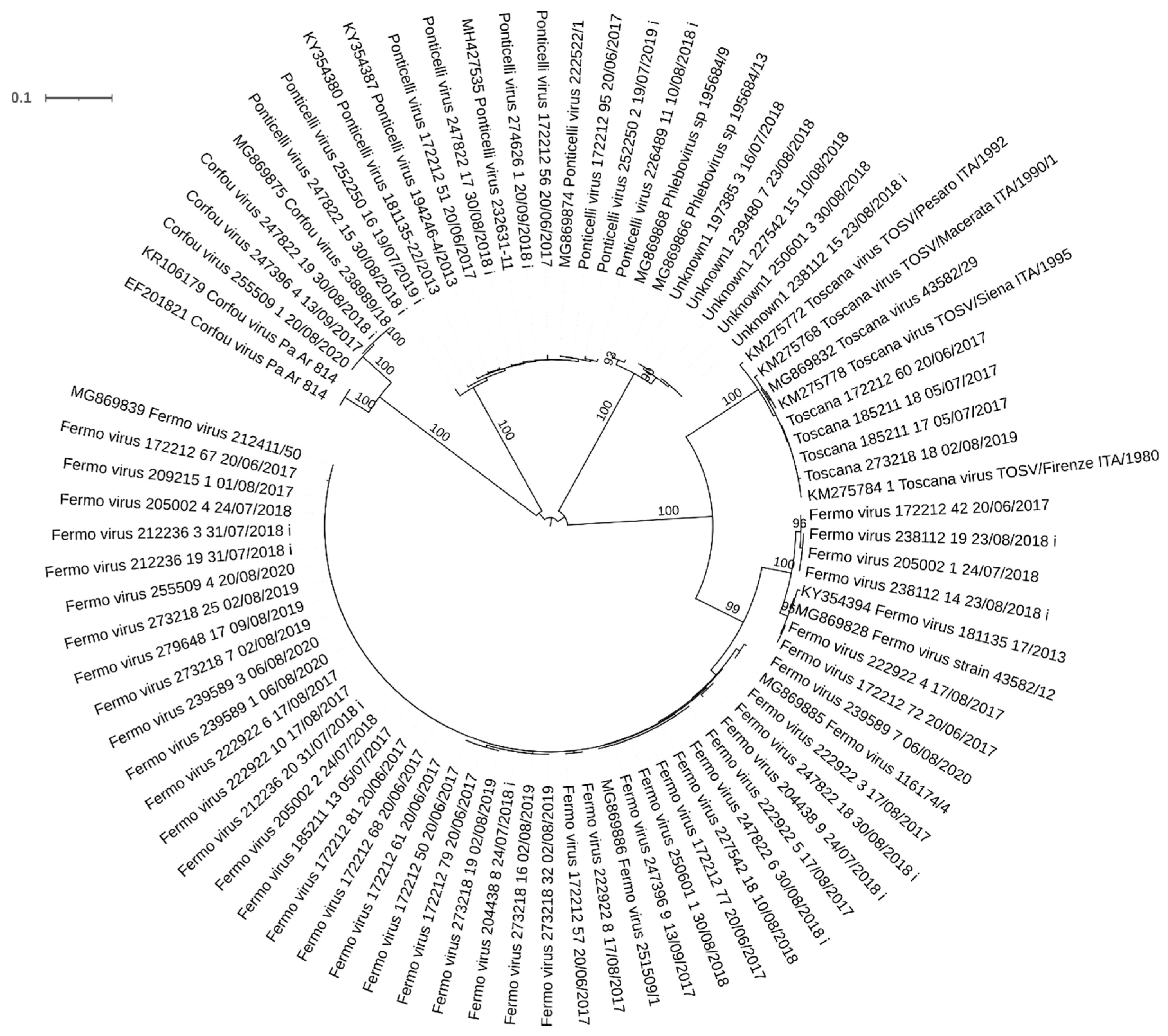
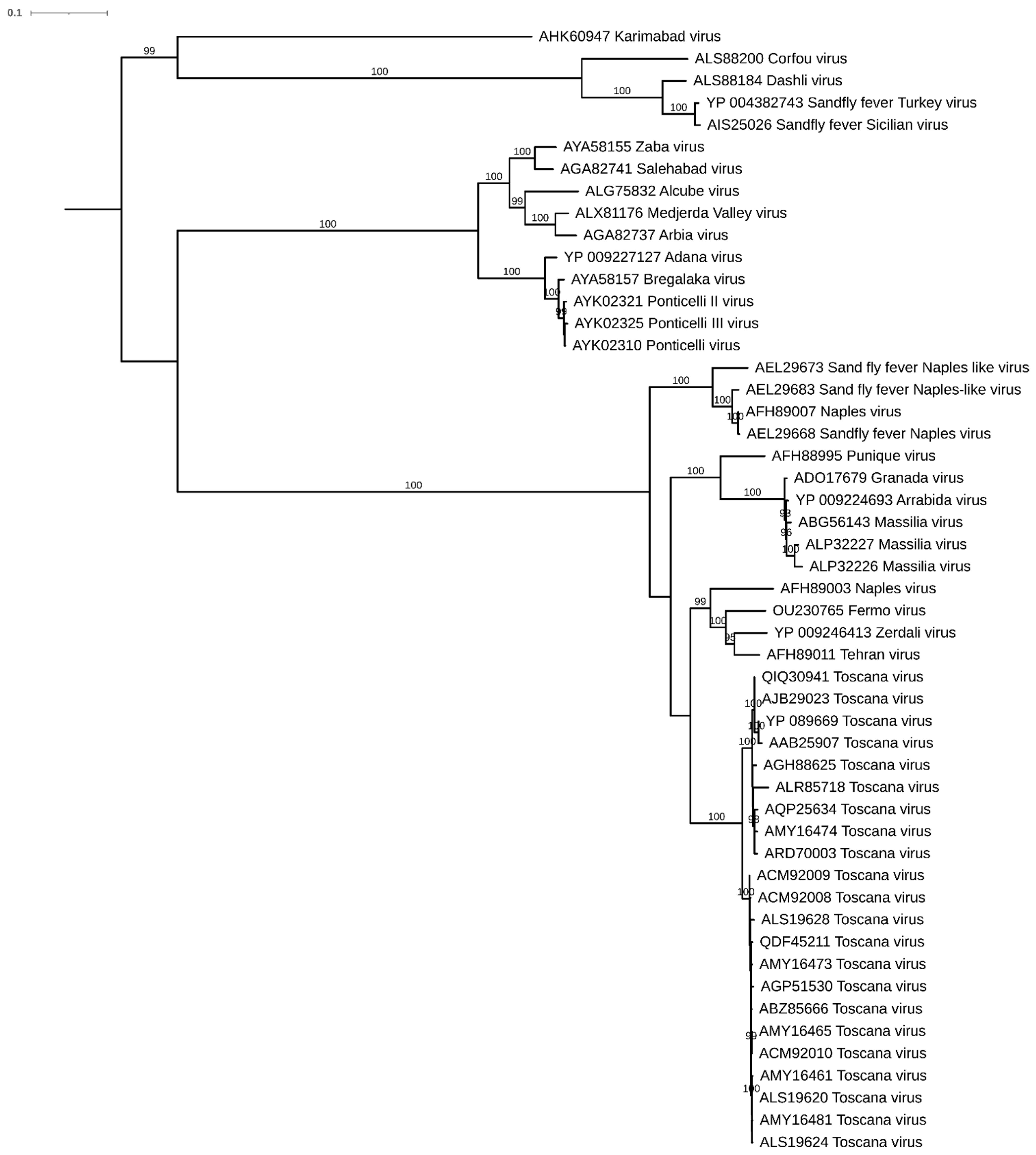
3.2. Molecular Analyses
3.3. Virus Isolation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palacios, G.; Tesh, R.; Travassos da Rosa, A.; Savji, N.; Sze, W.; Jain, K.; Serge, R.; Guzman, H.; Guevara, C.; Nunes, M.R.; et al. Characterization of the Candiru antigenic complex (Bunyaviridae: Phlebovirus), a highly diverse and reassorting group of viruses affecting humans in tropical America. J. Virol. 2011, 85, 3811–3820. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, J.H.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Amarasingh, G.K.; Anthony, S.J.; Avšič-Županc, T.; Ayllón, M.A.; Bahl, J.; Balkema-Buschmann, A.; et al. 2020 taxonomic update for phylum Negarnaviricota (Riboviria: Orthornavirae), including the large orders Bunyavirales and Mononegavirales. Arch. Virol. 2020, 165, 3023–3072. [Google Scholar] [CrossRef]
- Verani, P.; Ciufolini, M.G.; Nicoletti, L.; Balducci, M.; Sabatinelli, G.; Coluzzi, M.; Paci, P.; Amaducci, L. Ecological and epidemiological studies of Toscana virus, an arbovirus isolated from Phlebotomus. Ann. Ist. Super. Sanita. 1982, 18, 397–399. [Google Scholar] [PubMed]
- Calisher, C.H.; Weinberg, A.N.; Muth, D.J.; Lazuick, J.S. Toscana virus infection in United States citizen returning from Italy. Lancet 1987, 1, 165–166. [Google Scholar] [CrossRef]
- Moriconi, M.; Rugna, G.; Calzolari, M.; Bellini, R.; Albieri, A.; Angelini, P.; Cagarelli, R.; Landini, M.P.; Charrel, R.N.; Varani, S. Phlebotomine sand fly-borne pathogens in the Mediterranean Basin: Human leishmaniasis and phlebovirus infections. PLoS Negl. Trop. Dis. 2017, 11, e0005660. [Google Scholar] [CrossRef] [Green Version]
- Calzolari, M.; Chiapponi, C.; Bellini, R.; Bonilauri, P.; Lelli, D.; Moreno, A.; Barbieri, I.; Pongolini, S.; Lavazza, A.; Dottori, M. Isolation of three novel reassortant phleboviruses, Ponticelli I, II, III, and of Toscana virus from field-collected sand flies in Italy. Parasites Vectors 2018, 11, 84. [Google Scholar] [CrossRef] [Green Version]
- Calzolari, M.; Ferrarini, G.; Bonilauri, P.; Lelli, D.; Chiapponi, C.; Bellini, R.; Dottori, M. Co-circulation of eight different phleboviruses in sand flies collected in the Northern Apennine Mountains (Italy). Infect. Genet. Evol. 2018, 64, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M.; WHO Leishmaniasis Control Team. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE 2012, 7, e35671. [Google Scholar]
- Cardoso, L.; Schallig, H.; Persichetti, M.F.; Pennisi, M.G. New Epidemiological Aspects of Animal Leishmaniosis in Europe: The Role of Vertebrate Hosts Other Than Dogs. Pathogens 2021, 10, 307. [Google Scholar] [CrossRef]
- Pampiglione, S.; La Placa, M.; Schlick, G. Studies on mediterranean Leishmaniasis. I. An outbreak of visceral leishmaniasis in Northern Italy. Trans. R. Soc. Trop. Med. Hyg. 1974, 68, 349–359. [Google Scholar] [CrossRef]
- Varani, S.; Cagarelli, R.; Melchionda, F.; Attard, L.; Salvadori, C.; Finarelli, A.C.; Gentilomi, G.A.; Tigani, R.; Rangoni, R.; Todeschini, R.; et al. Ongoing outbreak of visceral leishmaniasis in Bologna Province, Italy, November 2012 to May 2013. Eurosurveillance 2013, 18, 20530. [Google Scholar] [CrossRef] [Green Version]
- Franceschini, E.; Puzzolante, C.; Menozzi, M.; Rossi, L.; Bedini, A.; Orlando, G.; Gennari, W.; Meacci, M.; Rugna, G.; Carra, E.; et al. Clinical and Microbiological Characteristics of Visceral Leishmaniasis Outbreak in a Northern Italian Nonendemic Area: A Retrospective Observational Study. BioMed Res. Int. 2016, 2016, 6481028. [Google Scholar] [CrossRef]
- Rugna, G.; Carra, E.; Corpus, F.; Calzolari, M.; Salvatore, D.; Bellini, R.; Di Francesco, A.; Franceschini, E.; Bruno, A.; Poglayen, G.; et al. Distinct Leishmania infantum Strains Circulate in Humans and Dogs in the Emilia-Romagna Region, Northeastern Italy. Vector Borne Zoonotic Dis. 2017, 17, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Rugna, G.; Carra, E.; Bergamini, F.; Calzolari, M.; Salvatore, D.; Corpus, F.; Gennari, W.; Baldelli, R.; Fabbi, M.; Natalini, S.; et al. Multilocus microsatellite typing (MLMT) reveals host-related population structure in Leishmania infantum from northeastern Italy. PLoS Negl. Trop. Dis. 2018, 12, e0006595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maroli, M.; Feliciangeli, M.D.; Bichaud, L.; Charrel, R.N.; Gradoni, L. Phlebotomine sandflies and the spreading of leishmaniases and other diseases of public health concern. Med. Vet. Entomol. 2013, 27, 123–147. [Google Scholar] [CrossRef]
- Calzolari, M.; Carra, E.; Rugna, G.; Bonilauri, P.; Bergamini, F.; Bellini, R.; Varani, S.; Dottori, M. Isolation and Molecular Typing of Leishmania infantum from Phlebotomus perfiliewi in a Re-Emerging Focus of Leishmaniasis, Northeastern Italy. Microorganisms 2019, 7, 644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romi, R.; Khoury, C.; Bigliocchi, F.; Maroli, M. Fact Sheet on Mites and Insects of Medical Importance. Schede Guida su Acari e Insetti di Interesse Sanitario; Istituto Superiore di Sanità: Rome, Italy, 1994. [Google Scholar]
- Dantas-Torres, F.; Tarallo, V.D.; Otranto, D. Morphological keys for the identification of Italian phlebotomine sand flies (Diptera: Psychodidae: Phlebotominae). Parasites Vectors 2014, 7, 479. [Google Scholar] [CrossRef]
- Lambert, A.J.; Lanciotti, R.S. Consensus amplification and novel multiplex sequencing method for S segment species identification of 47 viruses of the Orthobunyavirus, Phlebovirus, and Nairovirus genera of the family Bunyaviridae. J. Clin. Microbiol. 2009, 47, 2398–2404. [Google Scholar] [CrossRef] [Green Version]
- Galletti, E.; Bonilauri, P.; Bardasi, L.; Fontana, M.C.; Ramini, M.; Renzi, M.; Dosa, G.; Merialdi, G. Development of a minor groove binding probe based real-time PCR for the diagnosis and quantification of Leishmania infantum in dog specimens. Res. Vet. Sci. 2011, 91, 243–245. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Ayhan, N.; Alten, B.; Ivovic, V.; Dvořák, V.; Martinkovic, F.; Omeragic, J.; Stefanovska, J.; Petric, D.; Vaselek, S.; Baymak, D.; et al. Direct evidence for an expanded circulation area of the recently identified Balkan virus (Sandfly fever Naples virus species) in several countries of the Balkan archipelago. Parasit. Vectors 2017, 10, 402. [Google Scholar] [CrossRef] [Green Version]
- Corradetti, A. Phlebotomus and leishmaniasis in North-Central Italy (Apennine Region). Sci. Rep. Ist. Super. Sanità 1962, 2, 103–109. [Google Scholar]
- Vásquez Trujillo, A.; González Reina, A.E.; Góngora Orjuela, A.; Prieto Suárez, E.; Palomares, J.E.; Buitrago Alvarez, L.S. Seasonal variation and natural infection of Lutzomyia antunesi (Diptera: Psychodidae: Phlebotominae), an endemic species in the Orinoquia region of Colombia. Mem. Inst. Oswaldo Cruz. 2013, 108, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Duque, P.L.; Arrivillaga-Henríquez, J.; Enríquez, S.; Ron-Garrid, L.; Benítez, W.; Navarro, J.C. Spatial-Temporal Analysis of Lutzomyia trapidoi and Lutzomyia reburra (Diptera: Phlebotominae), in Rural Tourist Locations, Biosphere Reserve and Leishmaniasis Endemic Area, Ecuador. J. Med. Entomol. 2020, 57, 1905–1912. [Google Scholar] [CrossRef]
- Morini, S.; Calzolari, M.; Rossini, G.; Pascarelli, N.; Porcellini, A.; Randi, V.; Re, M.C.; Albieri, A.; Bonilaur, P.; Bellini, R.; et al. Detection of Specific Antibodies against Toscana Virus among Blood Donors in Northeastern Italy and Correlation with Sand Fly Abundance in 2014. Microorganisms 2020, 8, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattivi, A.; Massimiliani, E.; Cagarell, R.; Albieri, A. Leishmaniosi In Emilia-Romagna Aggiornamento epidemiologico 1999–2015. Report of Emilia-Romagna Region 2016. Available online: https://salute.regione.emilia-romagna.it/normativa-e-documentazione/rapporti/malattie-infettive (accessed on 10 May 2021).
- Rodhain, F.; Madulo-Leblond, G.; Hannoun, C.; Tesh, R.B. Le virus corfou: Un nouveau Phlebovirusisolé de phlébotomes en Grèce. Ann. L’institut Pasteur Virol. 1985, 136, 161–166. [Google Scholar] [CrossRef]
- Remoli, M.E.; Fortuna, C.; Marchi, A.; Bucci, P.; Argentini, C.; Bongiorno, G.; Maroli, M.; Gradoni, L.; Gramiccia, M.; Ciufolini, M.G. Viral isolates of a novel putative phlebovirus in the Marche Region of Italy. Am. J. Trop. Med. Hyg. 2014, 90, 760–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Percivalle, E.; Cassaniti, I.; Calzolari, M.; Lelli, D.; Baldanti, F. Thirteen Years of Phleboviruses Circulation in Lombardy, a Northern Italy Region. Viruses 2021, 13, 209. [Google Scholar] [CrossRef]
- Briese, T.; Bird, B.; Kapoor, V.; Nichol, S.T.; Lipkin, W.I. Batai and Ngari viruses: M segment reassortment and association with severe febrile disease outbreaks in East Africa. J. Virol. 2006, 80, 5627–5630. [Google Scholar] [CrossRef] [Green Version]
- Gerrard, S.R.; Li, L.; Barrett, A.D.; Nichol, S.T. Ngari virus is a Bunyamwera virus reassortant that can be associated with large outbreaks of hemorrhagic fever in Africa. J. Virol. 2004, 78, 8922–8926. [Google Scholar] [CrossRef] [Green Version]
- Ortalli, M.; De Pascali, A.M.; Longo, S.; Pascarelli, N.; Porcellini, A.; Ruggeri, D.; Randi, V.; Procopio, A.; Re, M.C.; Varani, S. Asymptomatic Leishmania infantum infection in blood donors living in an endemic area northeastern Italy. J. Infect. 2020, 80, 116–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bichaud, L.; Souris, M.; Mary, C.; Ninove, L.; Thirion, L.; Piarroux, R.P.; Piarroux, R.; De Lamballerie, X.; Charrel, R.N. Epidemiologic relationship between Toscana virus infection and Leishmania infantum due to common exposure to Phlebotomus perniciosus sandfly vector. PLoS Negl. Trop. Dis. 2011, 5, e1328. [Google Scholar] [CrossRef] [PubMed]
- Faucher, B.; Bichaud, L.; Charrel, R.; Mary, C.; Izri, A.; de Lamballerie, X.; Piarroux, R. Presence of sandflies infected with Leishmania infantum and Massilia virus in the Marseille urban area. Clin. Microbiol. Infect. 2014, 20, 340–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ergunay, K.; Kasap, O.E.; Orsten, S.; Oter, K.; Gunay, F.; Yoldar, A.Z.; Dincer, E.; Alten, B.; Ozkul, A. Phlebovirus and Leishmania detection in sandflies from eastern Thrace and northern Cyprus. Parasit. Vectors 2014, 7, 575. [Google Scholar] [CrossRef] [PubMed]
- Fares, W.; Dachraoui, K.; Barhoumi, W.; Cherni, S.; Chelbi, I.; Zhioua, E. Co-circulation of Toscana virus and Leishmania infantum in a focus of zoonotic visceral leishmaniasis from Central Tunisia. Acta Trop. 2020, 204, 105342. [Google Scholar] [CrossRef]
- Rath, C.T.; Schnellrath, L.C.; Damaso, C.R.; de Arruda, L.B.; Vasconcelos, P.F.D.C.; Gomes, C.; Laurenti, M.D.; Calegari, T.C.; Vivarini, Á.C.; Fasel, N.; et al. Amazonian Phlebovirus (Bunyaviridae) potentiates the infection of Leishmania (Leishmania) amazonensis: Role of the PKR/IFN1/IL-10 axis. PLoS Negl. Trop. Dis. 2019, 13, e0007500. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2017 | 2018 | 2019 | 2020 | Total | ||
|---|---|---|---|---|---|---|
| Collected sand flies | 25,966 | 7696 | 12,559 | 4289 | 50,510 | |
| Identified | Ph. perfiliewi | 768 | 1123 | 427 | 453 | 2771 |
| Ph perniciosus | 7 | 22 | 13 | 13 | 55 | |
| Tested | Leishmania/Pan-phlebo PCR | 5443 | 3363 | 5893 | 2090 | 16,789 |
| Isolation in cell cultures | 2460 | 500 | 2960 |
| 2017 | 2018 | 2019 | 2020 | Total | |
|---|---|---|---|---|---|
| Tested pools | 110 | 73 | 126 | 46 | 355 |
| Leishmania | 42 | 5 | 22 | 6 | 75 |
| Toscana virus | 4 | 2 | 6 | ||
| Fermo virus | 23 (10) | 5 (1) | 8 (2) | 6 (1) | 42 (14) |
| Ponticelli virus | 4 | 2 (1) | 1 | 7 (1) | |
| Corfou virus | 1 (1) | 1 | 2 (1) | ||
| Unknown1 | 4 | 4 |
| Data | Pool Code | Homogenate PCR | CPE 1 | Cryolysate PCR |
|---|---|---|---|---|
| 24/07/2018 | 204438-8 | Fermo virus | ++ | Fermo virus |
| 24/07/2018 | 204438-9 | Fermo virus | + | Fermo virus |
| 31/07/2018 | 212236-3 | Fermo virus | ++ | Fermo virus |
| 31/07/2018 | 212236-20 | Fermo virus | + | Fermo virus |
| 10/08/2018 | 226489-11 | Ponticelli virus | ++ | Ponticelli virus |
| 30/08/2018 | 247822-15 | Ponticelli virus | ++ | Ponticelli virus |
| 30/08/2018 | 247822-17 | Ponticelli virus | ++ | Ponticelli virus |
| 30/08/2018 | 247822-18 | Fermo virus | ++ | Ponticelli virus |
| 06/09/2018 | 256303-2 | Neg. | ++ | Ponticelli virus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calzolari, M.; Romeo, G.; Callegari, E.; Bonilauri, P.; Chiapponi, C.; Carra, E.; Rugna, G.; Taddei, R.; Lelli, D.; Dottori, M. Co-Circulation of Phleboviruses and Leishmania Parasites in Sand Flies from a Single Site in Italy Monitored between 2017 and 2020. Viruses 2021, 13, 1660. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081660
Calzolari M, Romeo G, Callegari E, Bonilauri P, Chiapponi C, Carra E, Rugna G, Taddei R, Lelli D, Dottori M. Co-Circulation of Phleboviruses and Leishmania Parasites in Sand Flies from a Single Site in Italy Monitored between 2017 and 2020. Viruses. 2021; 13(8):1660. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081660
Chicago/Turabian StyleCalzolari, Mattia, Giuseppe Romeo, Emanuele Callegari, Paolo Bonilauri, Chiara Chiapponi, Elena Carra, Gianluca Rugna, Roberta Taddei, Davide Lelli, and Michele Dottori. 2021. "Co-Circulation of Phleboviruses and Leishmania Parasites in Sand Flies from a Single Site in Italy Monitored between 2017 and 2020" Viruses 13, no. 8: 1660. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081660