
The Molecular Tweezer CLR01 Inhibits Antibody-Resistant Cell-to-Cell Spread of Human Cytomegalovirus

 , , , , , and
, , , , , and 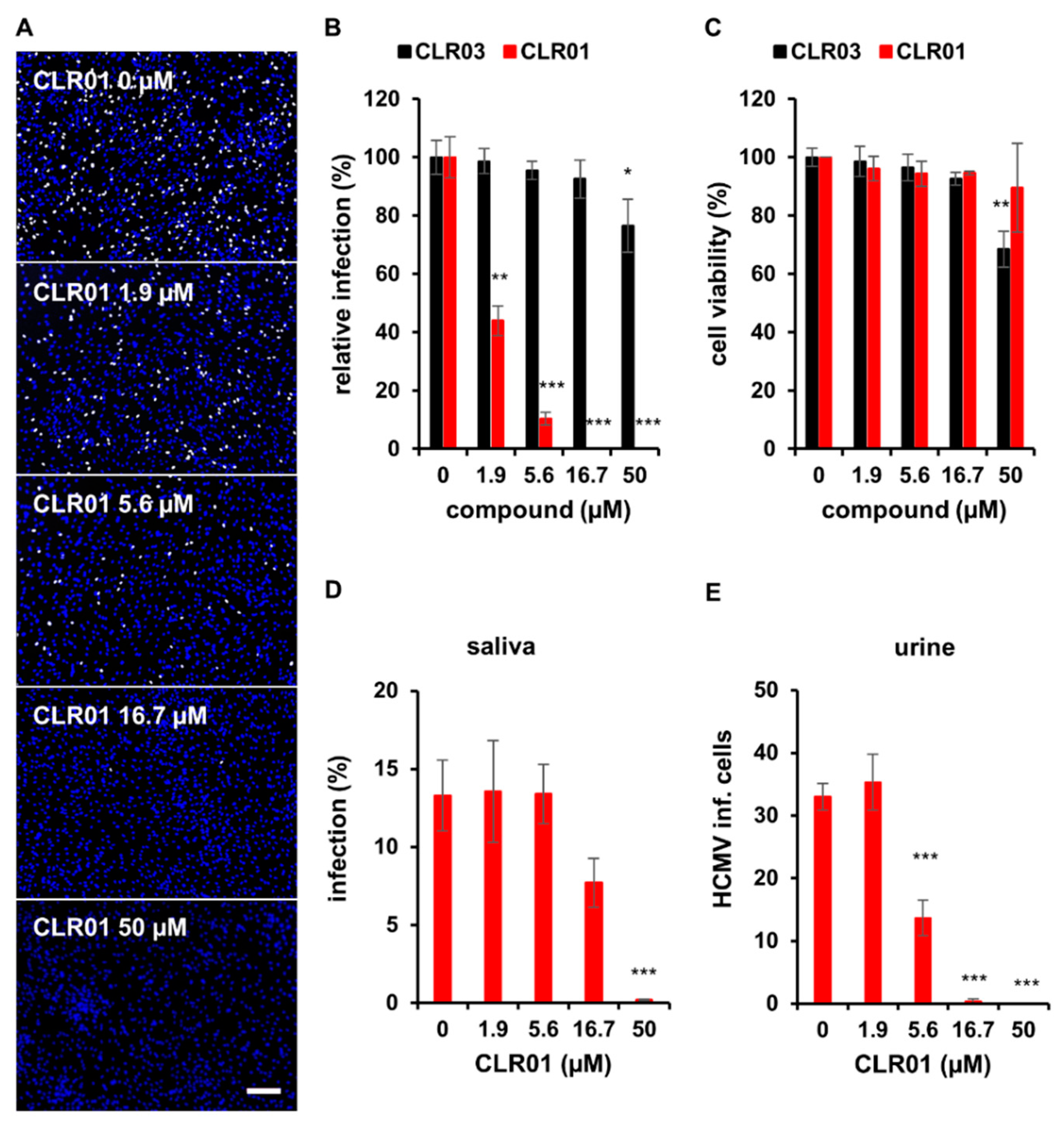{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Virus Strains
2.2. Reagents and Samples
2.3. Detection of Infected Cells
2.4. Infection Assays
2.5. Focus Expansion Assays
2.6. Cytotoxicity
2.7. Attachment Assay
2.8. RT-qPCR
2.9. Electron Microscopy
2.10. Quantifications and Statistical Analysis
3. Results
3.1. The Molecular Tweezer CLR01 Inhibits HCMV Infection
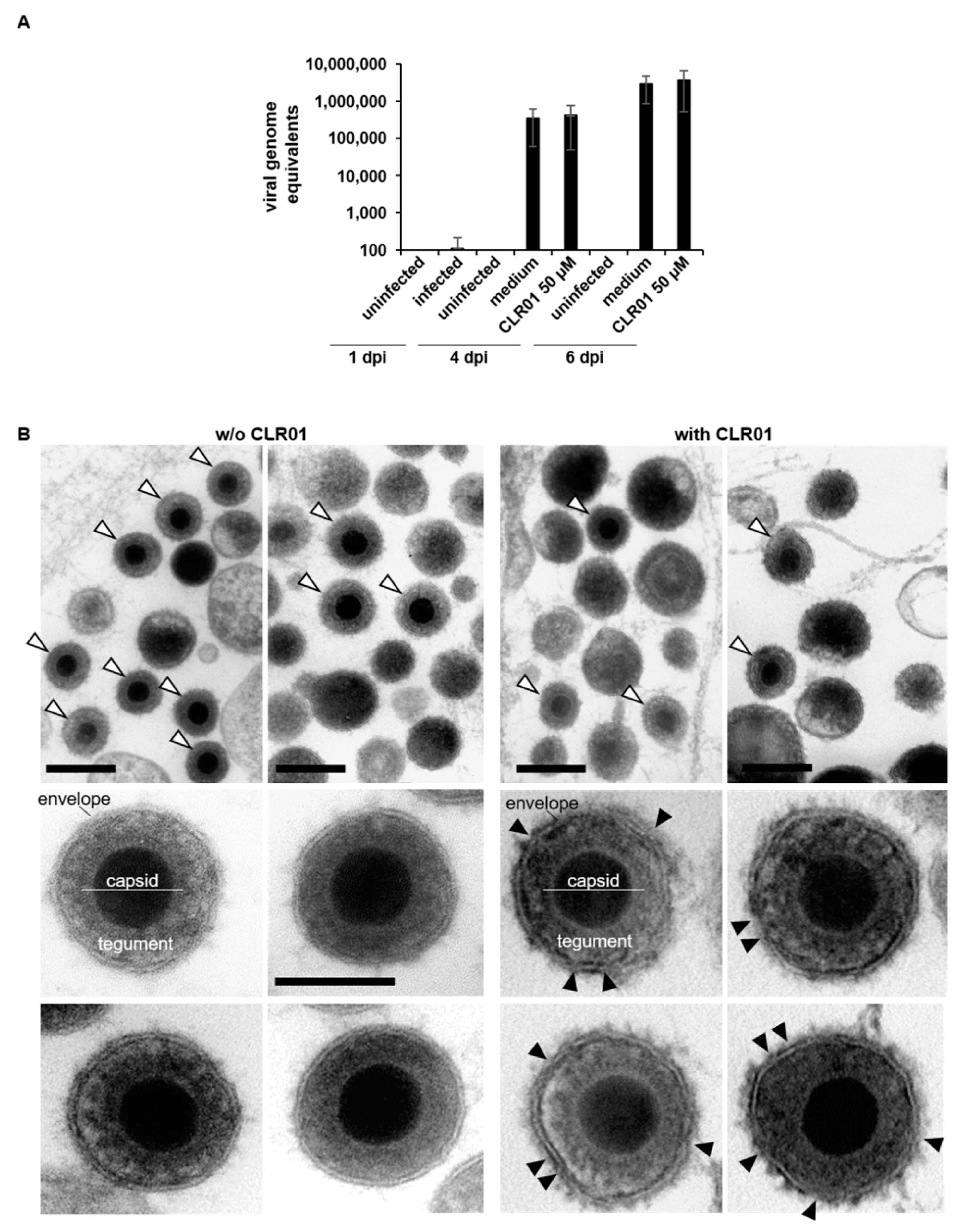
3.2. CLR01 Has a Fast-Acting Mode of Action That Prevents Entry at Various Steps
3.3. CLR01 Inhibits Direct Cell-to-Cell Spread of HCMV
3.4. Inhibition of Herpesvirus Cell-to-Cell Spread by CLR01 Is Not Restricted to HCMV
4. Discussion
4.1. CLR01 Inhibits HCMV Entry
4.2. CLR01 Potently Blocks Antibody-Resistant Cell-to-Cell Spread of HCMV
4.3. Outlook
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sattentau, Q. Avoiding the void: Cell-to-cell spread of human viruses. Nat. Rev. Microbiol. 2008, 6, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, A.; Arndt, M.A.E.; Grosse-Hovest, L.; Weichert, W.; Giebel, B.; Dittmer, U.; Hengel, H.; Jäger, D.; Schneweis, K.E.; Eis-Hübinger, A.M.; et al. Overcoming drug-resistant herpes simplex virus (HSV) infection by a humanized antibody. Proc. Natl. Acad. Sci. USA 2013, 110, 6760–6765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clementi, N.; Criscuolo, E.; Cappelletti, F.; Quaranta, P.; Pistello, M.; Diotti, R.A.; Sautto, G.A.; Tarr, A.W.; Mailland, F.; Concas, D.; et al. Entry inhibition of HSV-1 and -2 protects mice from viral lethal challenge. Antivir. Res. 2017, 143, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Mothes, W.; Sherer, N.M.; Jin, J.; Zhong, P. Virus Cell-to-Cell Transmission. J. Virol. 2010, 84, 8360–8368. [Google Scholar] [CrossRef] [Green Version]
- Ganesh, L.; Leung, K.; Loré, K.; Levin, R.; Panet, A.; Schwartz, O.; Koup, R.A.; Nabel, G.J. Infection of specific dendritic cells by CCR5-tropic human immunodeficiency virus type 1 promotes cell-mediated transmission of virus resistant to broadly neutralizing antibodies. J. Virol. 2004, 78, 11980–11987. [Google Scholar] [CrossRef] [Green Version]
- Jacob, C.L.; Lamorte, L.; Sepulveda, E.; Lorenz, I.C.; Gauthier, A.; Franti, M. Neutralizing antibodies are unable to inhibit direct viral cell-to-cell spread of human cytomegalovirus. Virology 2013, 444, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Murrell, I.; Bedford, C.; Ladell, K.; Miners, K.L.; Price, D.A.; Tomasec, P.; Wilkinson, G.W.G.; Stanton, R.J. The pentameric complex drives immunologically covert cell-cell transmission of wild-type human cytomegalovirus. Proc. Natl. Acad. Sci. USA 2017, 114, 6104–6109. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Kim, J.H.; Ranjan, P.; Metcalfe, M.G.; Cao, W.; Mishina, M.; Gangappa, S.; Guo, Z.; Boyden, E.S.; Zaki, S.; et al. Influenza virus exploits tunneling nanotubes for cell-to-cell spread. Sci. Rep. 2017, 7, 40360. [Google Scholar] [CrossRef]
- Sinzger, C.; Schmidt, K.; Knapp, J.; Kahl, M.; Beck, R.; Waldman, J.; Hebart, H.; Einsele, H.; Jahn, G. Modification of human cytomegalovirus tropism through propagation in vitro is associated with changes in the viral genome. J. Gen. Virol. 1999, 80, 2867–2877. [Google Scholar] [CrossRef]
- Jackson, J.W.; Sparer, T. There is always another way! cytomegalovirus’ multifaceted dissemination schemes. Viruses 2018, 10, 383. [Google Scholar] [CrossRef] [Green Version]
- Sattentau, Q.J. The direct passage of animal viruses between cells. Curr. Opin. Virol. 2011, 1, 396–402. [Google Scholar] [CrossRef]
- Panasiuk, M.; Rychłowski, M.; Derewońko, N.; Bieńkowska-Szewczyk, K. Tunneling nanotubes (TNT) as a novel route of cell-to-cell spread of herpesviruses. J. Virol. 2018, 92, JVI.00090-18. [Google Scholar] [CrossRef] [Green Version]
- Falk, J.J.; Winkelmann, M.; Laib Sampaio, K.; Paal, C.; Schrezenmeier, H.; Alt, M.; Stanton, R.; Krawczyk, A.; Lotfi, R.; Sinzger, C. Large-Scale Screening of HCMV-Seropositive Blood Donors Indicates that HCMV Effectively Escapes from Antibodies by Cell-Associated Spread. Viruses 2018, 10, 500. [Google Scholar] [CrossRef] [Green Version]
- Fokkens, M.; Schrader, T.; Klärner, F.-G. A molecular tweezer for lysine and arginine. J. Am. Chem. Soc. 2005, 127, 14415–14421. [Google Scholar] [CrossRef]
- Sinha, S.; Lopes, D.H.J.; Du, Z.; Pang, E.S.; Shanmugam, A.; Lomakin, A.; Talbiersky, P.; Tennstaedt, A.; McDaniel, K.; Bakshi, R.; et al. Lysine-specific molecular tweezers are broad-spectrum inhibitors of assembly and toxicity of amyloid proteins. J. Am. Chem. Soc. 2011, 133, 16958–16969. [Google Scholar] [CrossRef] [Green Version]
- Schrader, T.; Bitan, G.; Klärner, F.-G. Molecular tweezers for lysine and arginine—Powerful inhibitors of pathologic protein aggregation. Chem. Commun. 2016, 52, 11318–11334. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, N.; Pereira-Henriques, A.; Attar, A.; Klärner, F.G.; Schrader, T.; Bitan, G.; Gales, L.; Saraiva, M.J.; Almeida, M.R. Molecular Tweezers Targeting Transthyretin Amyloidosis. Neurotherapeutics 2014, 11, 450–461. [Google Scholar] [CrossRef]
- Prabhudesai, S.; Sinha, S.; Attar, A.; Kotagiri, A.; Fitzmaurice, A.G.; Lakshmanan, R.; Ivanova, M.I.; Loo, J.A.; Klärner, F.G.; Schrader, T.; et al. A Novel “Molecular Tweezer” Inhibitor of α-Synuclein Neurotoxicity in Vitro and in Vivo. Neurotherapeutics 2012, 9, 464–476. [Google Scholar] [CrossRef] [Green Version]
- Mittal, S.; Bravo-Rodriguez, K.; Sanchez-Garcia, E. Mechanism of Inhibition of Beta Amyloid Toxicity by Supramolecular Tweezers. J. Phys. Chem. B 2018, 122, 4196–4205. [Google Scholar] [CrossRef]
- Attar, A.; Ripoli, C.; Riccardi, E.; Maiti, P.; Li Puma, D.D.; Liu, T.; Hayes, J.; Jones, M.R.; Lichti-Kaiser, K.; Yang, F.; et al. Protection of primary neurons and mouse brain from Alzheimer’s pathology by molecular tweezers. Brain 2012, 135, 3735–3748. [Google Scholar] [CrossRef] [Green Version]
- Attar, A.; Chan, W.-T.C.; Klärner, F.-G.; Schrader, T.; Bitan, G. Safety and pharmacological characterization of the molecular tweezer CLR01—A broad-spectrum inhibitor of amyloid proteins’ toxicity. BMC Pharmacol. Toxicol. 2014, 15, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lump, E.; Castellano, L.M.; Meier, C.; Seeliger, J.; Erwin, N.; Sperlich, B.; Stürzel, C.M.; Usmani, S.; Hammond, R.M.; von Einem, J.; et al. A molecular tweezer antagonizes seminal amyloids and HIV infection. eLife 2015, 4, e05397. [Google Scholar] [CrossRef] [PubMed]
- Röcker, A.E.; Müller, J.A.; Dietzel, E.; Harms, M.; Krüger, F.; Heid, C.; Sowislok, A.; Riber, C.F.; Kupke, A.; Lippold, S.; et al. The molecular tweezer CLR01 inhibits Ebola and Zika virus infection. Antivir. Res. 2018, 152, 26–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weil, T.; Groß, R.; Röcker, A.; Bravo-Rodriguez, K.; Heid, C.; Sowislok, A.; Le, M.; Erwin, N.; Dwivedi, M.; Bart, S.; et al. Supramolecular Mechanism of Viral Envelope Disruption by Molecular Tweezers. J. Am. Chem. Soc. 2020, 142, 17024–17038. [Google Scholar] [CrossRef]
- Bavari, S.; Bosio, C.M.; Wiegand, E.; Ruthel, G.; Will, A.B.; Geisbert, T.W.; Hevey, M.; Schmaljohn, C.; Schmaljohn, A.; Aman, M.J. Lipid Raft Microdomains. J. Exp. Med. 2002, 195, 593–602. [Google Scholar] [CrossRef]
- Bender, F.C.; Whitbeck, J.C.; Ponce de Leon, M.; Lou, H.; Eisenberg, R.J.; Cohen, G.H. Specific association of glycoprotein B with lipid rafts during herpes simplex virus entry. J. Virol. 2003, 77, 9542–9552. [Google Scholar] [CrossRef] [Green Version]
- Low, H.; Mukhamedova, N.; Cui, H.L.; McSharry, B.P.; Avdic, S.; Hoang, A.; Ditiatkovski, M.; Liu, Y.; Fu, Y.; Meikle, P.J.; et al. Cytomegalovirus Restructures Lipid Rafts via a US28/CDC42-Mediated Pathway, Enhancing Cholesterol Efflux from Host Cells. Cell Rep. 2016, 16, 186–200. [Google Scholar] [CrossRef] [Green Version]
- Lippold, S.; Braun, B.; Krüger, F.; Harms, M.; Müller, J.A.; Groß, R.; Münch, J.; von Einem, J. Natural inhibitor of human cytomegalovirus in human seminal plasma. J. Virol. 2019, 93, e01855-18. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, A.; Krauss, J.; Eis-Hubinger, A.M.; Daumer, M.P.; Schwarzenbacher, R.; Dittmer, U.; Schneweis, K.E.; Jager, D.; Roggendorf, M.; Arndt, M.A.E. Impact of Valency of a Glycoprotein B-Specific Monoclonal Antibody on Neutralization of Herpes Simplex Virus. J. Virol. 2011, 85, 1793–1803. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Shenk, T. Human Cytomegalovirus UL131 Open Reading Frame Is Required for Epithelial Cell Tropism. J. Virol. 2005, 79, 10330–10338. [Google Scholar] [CrossRef] [Green Version]
- Sampaio, K.L.; Jahn, G.; Sinzger, C. Applications for a dual fluorescent human cytomegalovirus in the analysis of viral entry. Methods Mol. Biol. 2013, 1064, 201–209. [Google Scholar] [CrossRef]
- Taylor, J.M.; Lin, E.; Susmarski, N.; Yoon, M.; Zago, A.; Ware, C.F.; Pfeffer, K.; Miyoshi, J.; Takai, Y.; Spear, P.G. Alternative Entry Receptors for Herpes Simplex Virus and Their Roles in Disease. Cell Host Microbe 2007, 2, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Talbiersky, P.; Bastkowski, F.; Klärner, F.G.; Schrader, T. Molecular clip and tweezer introduce new mechanisms of enzyme inhibition. J. Am. Chem. Soc. 2008, 130, 9824–9828. [Google Scholar] [CrossRef]
- Andreoni, M.; Faircloth, M.; Vugler, L.; Britt, W.J. A rapid microneutralization assay for the measurement of neutralizing antibody reactive with human cytomegalovirus. J. Virol. Methods 1989, 23, 157–167. [Google Scholar] [CrossRef]
- Falk, J.J.; Winkelmann, M.; Stöhr, D.; Alt, M.; Schrezenmeier, H.; Krawczyk, A.; Lotfi, R.; Sinzger, C. Identification of elite neutralizers with broad and potent neutralizing activity against human cytomegalovirus (hcmv) in a population of hcmv-seropositive blood donors. J. Infect. Dis. 2018, 218, 876–885. [Google Scholar] [CrossRef]
- Lorz, K.; Hofmann, H.; Berndt, A.; Tavalai, N.; Mueller, R.; Schlotzer-Schrehardt, U.; Stamminger, T. Deletion of Open Reading Frame UL26 from the Human Cytomegalovirus Genome Results in Reduced Viral Growth, Which Involves Impaired Stability of Viral Particles. J. Virol. 2006, 80, 5423–5434. [Google Scholar] [CrossRef] [Green Version]
- Tavalai, N.; Papior, P.; Rechter, S.; Leis, M.; Stamminger, T. Evidence for a Role of the Cellular ND10 Protein PML in Mediating Intrinsic Immunity against Human Cytomegalovirus Infections. J. Virol. 2006, 80, 8006–8018. [Google Scholar] [CrossRef] [Green Version]
- Read, C.; Schauflinger, M.; Nikolaenko, D.; Walther, P.; Von Einem, J. Regulation of human cytomegalovirus secondary envelopment by a C-terminal tetralysine motif in pUL71. J. Virol. 2019, 93, e02244-18. [Google Scholar] [CrossRef] [Green Version]
- Compton, T.; Nowlin, D.M.; Cooper, N.R. Initiation of Human Cytomegalovirus Infection Requires Initial Interaction with Cell Surface Heparan Sulfate. Virology 1993, 193, 834–841. [Google Scholar] [CrossRef]
- Schauflinger, M.; Fischer, D.; Schreiber, A.; Chevillotte, M.; Walther, P.; Mertens, T.; von Einem, J. The tegument protein UL71 of human cytomegalovirus is involved in late envelopment and affects multivesicular bodies. J. Virol. 2011, 85, 3821–3832. [Google Scholar] [CrossRef] [Green Version]
- Dingwell, K.S.; Brunetl, C.R.; Hendricks, R.L.; Tang, Q.; Tang, M.; Rainbow, A.J.; Johnson, D.C. Herpes simplex virus glycoproteins E and I facilitate cell-to- cell spread in vivo and across junctions of cultured cells. J. Virol. 1994, 68, 834–845. [Google Scholar] [CrossRef] [Green Version]
- Ralph, M.; Bednarchik, M.; Tomer, E.; Rafael, D.; Zargarian, S.; Gerlic, M.; Kobiler, O. Promoting simultaneous onset of viral gene expression among cells infected with herpes simplex virus-1. Front. Microbiol. 2017, 8, 2152. [Google Scholar] [CrossRef] [Green Version]
- Waldman, W.; Sneddon, J.; Stephens, R.; Roberts, W. Enhanced endothelial cytopathogenicity induced by a cytomegalovirus strain propagated in endothelial cells. J. Med. Virol. 1989, 28, 223–230. [Google Scholar] [CrossRef]
- Digel, M.; Sampaio, K.L.; Jahn, G.; Sinzger, C. Evidence for direct transfer of cytoplasmic material from infected to uninfected cells during cell-associated spread of human cytomegalovirus. J. Clin. Virol. 2006, 37, 10–20. [Google Scholar] [CrossRef]
- Silva, M.C.; Schröer, J.; Shenk, T. Human cytomegalovirus cell-to-cell spread in the absence of an essential assembly protein. Proc. Natl. Acad. Sci. USA 2005, 102, 2081. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Lee, R.; Adler, S.P.; McVoy, M.A. Antibody inhibition of human cytomegalovirus spread in epithelial cell cultures. J. Virol. Methods 2013, 192, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.C.; Huber, M.T. Directed egress of animal viruses promotes cell-to-cell spread. J. Virol. 2002, 76, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agelidis, A.M.; Shukla, D. Cell entry mechanisms of HSV: What we have learned in recent years. Future Virol. 2015, 10, 1145. [Google Scholar] [CrossRef] [Green Version]
- Isaacson, M.K.; Compton, T. Human Cytomegalovirus Glycoprotein B Is Required for Virus Entry and Cell-to-Cell Spread but Not for Virion Attachment, Assembly, or Egress. J. Virol. 2009, 83, 3891–3903. [Google Scholar] [CrossRef] [Green Version]
- Weiler, N.; Paal, C.; Adams, K.; Calcaterra, C.; Fischer, D.; Stanton, R.J.; Stöhr, D.; Sampaio, K.L.; Sinzger, C. Role of Envelope Glycoprotein Complexes in Cell-Associated Spread of Human Cytomegalovirus. Viruses 2021, 13, 614. [Google Scholar] [CrossRef]
- Vanarsdall, A.L.; Ryckman, B.J.; Chase, M.C.; Johnson, D.C. Human Cytomegalovirus Glycoproteins gB and gH/gL Mediate Epithelial Cell-Cell Fusion When Expressed either in cis or in trans. J. Virol. 2008, 82, 11837–11850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Frascaroli, G.; Lebbink, R.J.; Ostermann, E.; Brune, W. Human cytomegalovirus glycoprotein B variants affect viral entry, cell fusion, and genome stability. Proc. Natl. Acad. Sci. USA 2019, 116, 18021–18030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galitska, G.; Biolatti, M.; De Andrea, M.; Leone, A.; Coscia, A.; Bertolotti, L.; Ala, U.; Bertino, E.; Dell’Oste, V.; Landolfo, S. Biological relevance of Cytomegalovirus genetic variability in congenitally and postnatally infected children. J. Clin. Virol. 2018, 108, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Malbec, M.; Porrot, F.; Rua, R.; Horwitz, J.; Klein, F.; Halper-Stromberg, A.; Scheid, J.F.; Eden, C.; Mouquet, H.; Nussenzweig, M.C.; et al. Broadly neutralizing antibodies that inhibit HIV-1 cell to cell transmission. J. Exp. Med. 2013, 210, 2813–2821. [Google Scholar] [CrossRef] [Green Version]
- Cagno, V.; Tintori, C.; Civra, A.; Cavalli, R.; Tiberi, M.; Botta, L.; Brai, A.; Poli, G.; Tapparel, C.; Lembo, D.; et al. Novel broad spectrum virucidal molecules against enveloped viruses. PLoS ONE 2018, 13, e0208333. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brenner, S.; Braun, B.; Read, C.; Weil, T.; Walther, P.; Schrader, T.; Münch, J.; von Einem, J. The Molecular Tweezer CLR01 Inhibits Antibody-Resistant Cell-to-Cell Spread of Human Cytomegalovirus. Viruses 2021, 13, 1685. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091685
Brenner S, Braun B, Read C, Weil T, Walther P, Schrader T, Münch J, von Einem J. The Molecular Tweezer CLR01 Inhibits Antibody-Resistant Cell-to-Cell Spread of Human Cytomegalovirus. Viruses. 2021; 13(9):1685. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091685
Chicago/Turabian StyleBrenner, Sina, Berenike Braun, Clarissa Read, Tatjana Weil, Paul Walther, Thomas Schrader, Jan Münch, and Jens von Einem. 2021. "The Molecular Tweezer CLR01 Inhibits Antibody-Resistant Cell-to-Cell Spread of Human Cytomegalovirus" Viruses 13, no. 9: 1685. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091685