
Meat Exudate for Detection of African Swine Fever Virus Genomic Material and Anti-ASFV Antibodies

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Pigs and Viruses
2.3. Inoculation and Sample Collection
2.4. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Statistics
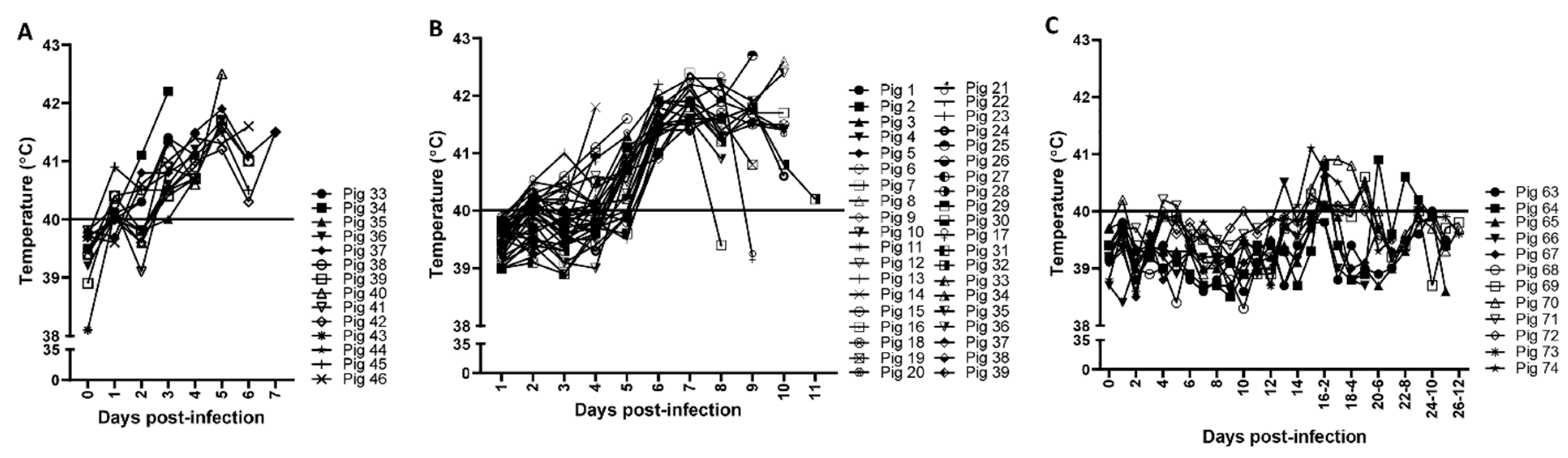
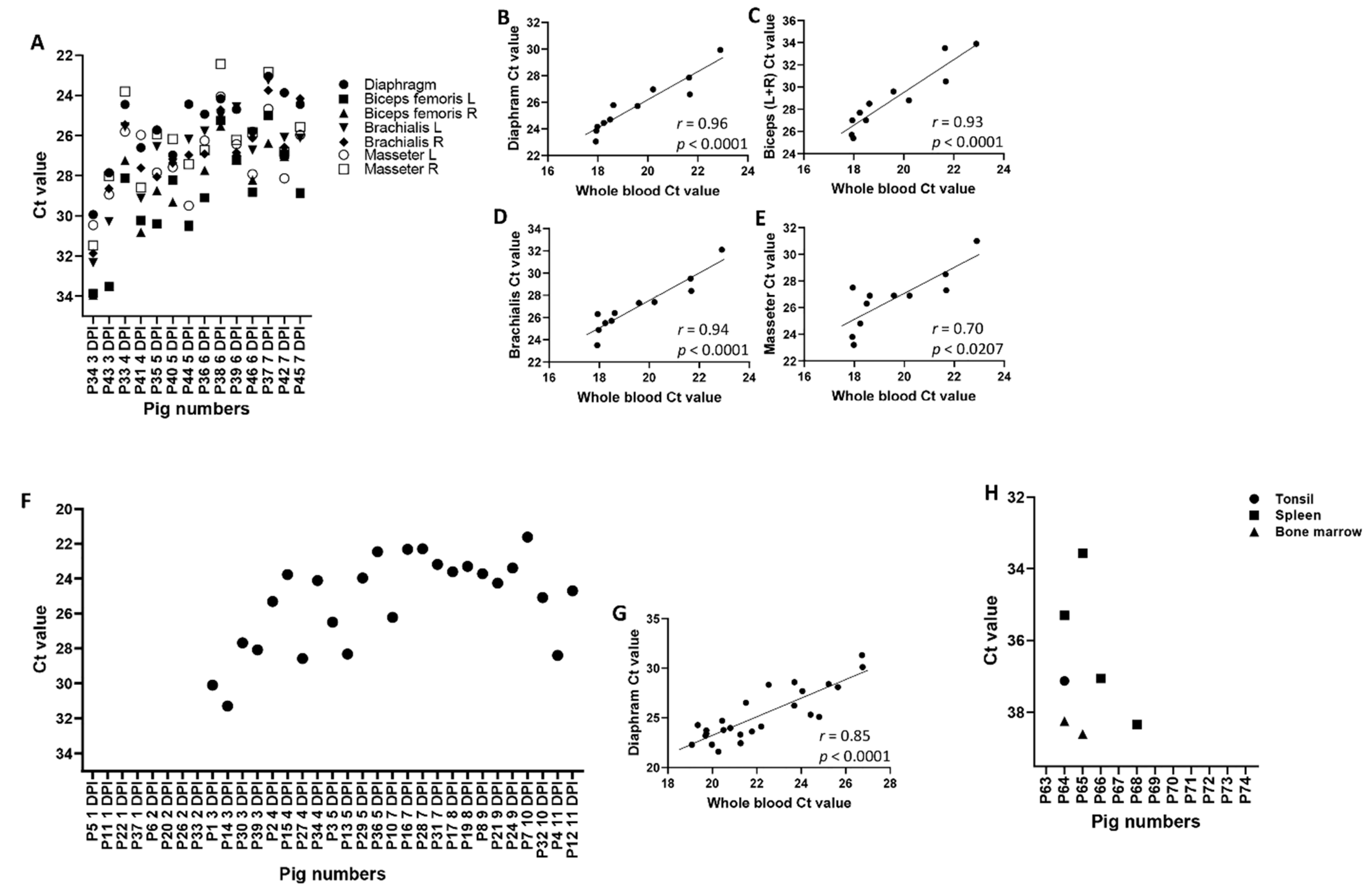
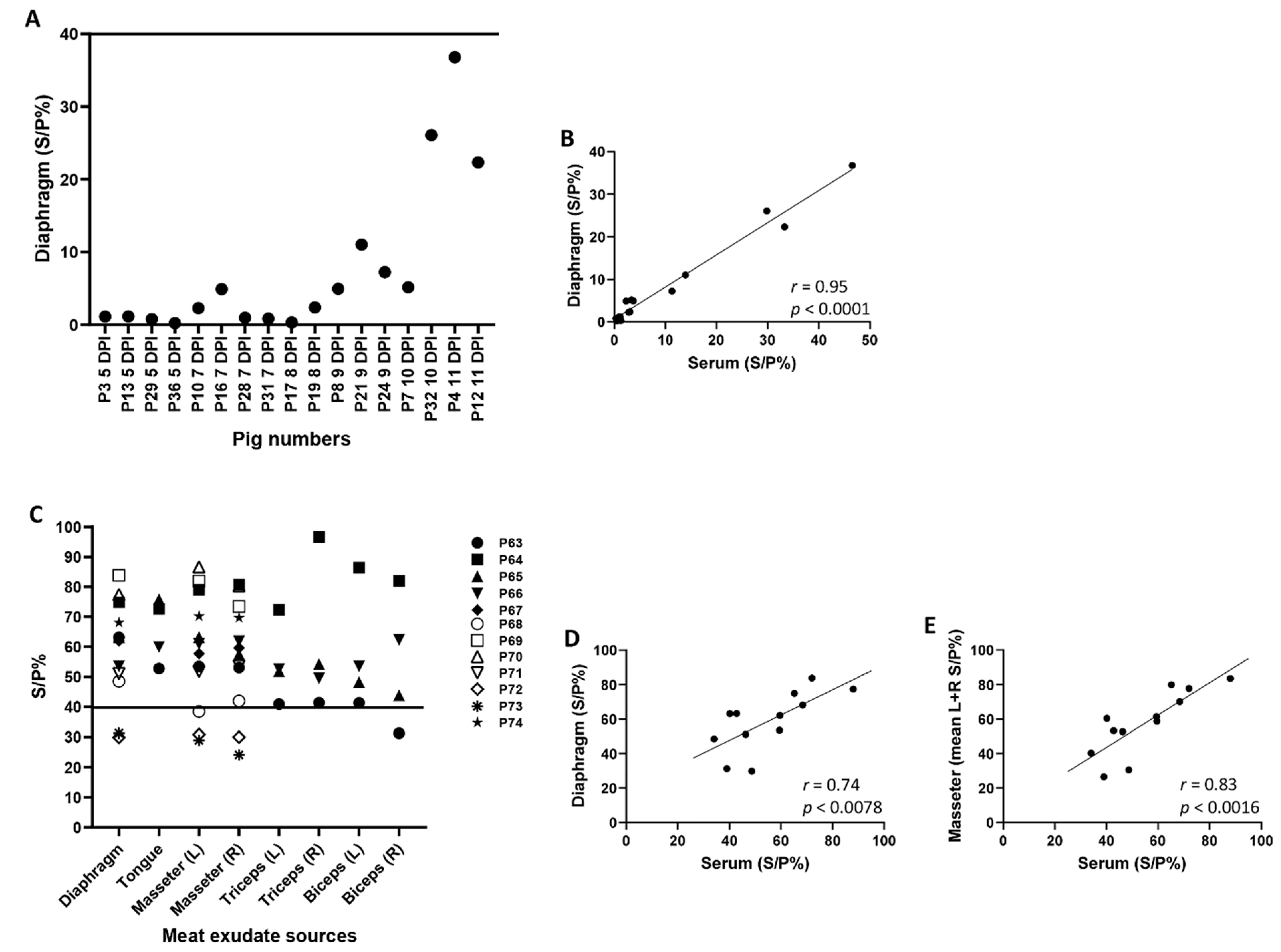
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Penrith, M.L.; Vosloo, W. Review of African swine fever: Transmission, spread and control. J. S. Afr. Vet. Assoc. 2009, 80, 58–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, R.E. On a form of swine fever occurring in British East Africa (Kenya Colony). J. Comp. Pathol. Ther. 1921, 34, 159–191. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.M.; Azevedo, J.R. La peste porcine africaine au Portugal. Bull. Off. Int. Epizoot. 1961, 55, 88–106. [Google Scholar]
- Costard, S.; Wieland, B.; de Glanville, W.; Jori, F.; Rowlands, R.; Vosloo, W.; Roger, F.; Pfeiffer, D.U.; Dixon, L.K. African swine fever: How can global spread be prevented? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2683–2696. [Google Scholar] [CrossRef] [Green Version]
- Alexander, F.C. Experiences with African swine fever in Haiti. Ann. N. Y. Acad. Sci. 1992, 653, 251–256. [Google Scholar] [CrossRef]
- Wilkinson, P. African swine fever virus. In Virus Infections of Porcines; Pensaert, M.B., Ed.; Elsevier Science: New York, NY, USA, 1989; pp. 17–35. [Google Scholar]
- European Food Safety Authority; Depner, K.; Gortazar, C.; Guberti, V.; Masiulis, M.; More, S.; Oļševskis, E.; Thulke, H.H.; Viltrop, A.; Woźniakowski, G.; et al. Epidemiological analyses of African swine fever in the Baltic States and Poland: (Update September 2016–September 2017). EFSA J. 2017, 15, e05068. [Google Scholar] [PubMed] [Green Version]
- Berg, C.; Botner, A.; Browman, H.; De Koeijer, A.; Domingo, M.; Ducrot, C.; Edwards, S.; Fourichon, C.; Koenen, F.; More, S.; et al. African swine fever EFSA panel on animal health and welfare (AHAW). EFSA J. 2015, 13, 4163. [Google Scholar]
- European Food Safety Authority; Anette, B.; Anette, B.; Theodora, C.V.; Klaus, D.; Daniel, D.; Vittorio, G.; Georgina, H.; Daniela, K.; Annick, L.; et al. Epidemiological analyses of African swine fever in the European Union (November 2018 to October 2019). EFSA J. 2020, 18, e05996. [Google Scholar]
- Dixon, L.K.; Escribano, J.; Martins, C.; Rock, D.L.; Salas, M.; Wilkinson, P.J. Asfarviridae. In Virus Taxonomy; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; VIIIth Report of the ICTV; Elsevier/Academic Press: London, UK, 2005; pp. 135–143. [Google Scholar]
- Rowlands, R.J.; Michaud, V.; Heath, L.; Hutchings, G.; Oura, C.; Vosloo, W.; Dwarka, R.; Onashvili, T.; Albina, E.; Dixon, L.K. African swine fever virus isolate, Georgia, 2007. Emerg. Infect. Dis. 2008, 14, 1870–1874. [Google Scholar] [CrossRef]
- Beltrán-Alcrudo, D.; Lubroth, J.; Depner, K.; De La Rocque, S. African swine fever in the Caucasus. FAO Empres Watch 2008, 1, 1–8. [Google Scholar]
- Bastos, A.D.S.; Penrith, M.-L.; Macome, F.; Pinto, F.; Thomson, G.R. Co-circulation of two genetically distinct viruses in an outbreak of African swine fever in Mozambique: No evidence for individual co-infection. Vet. Microbiol. 2004, 103, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Boshoff, C.I.; Bastos, A.D.; Gerber, L.J.; Vosloo, W. Genetic characterisation of African swine fever viruses from outbreaks in southern Africa (1973–1999). Vet. Microbiol. 2007, 121, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Penrith, M.L.; Lopes Pereira, C.; Lopes da Silva, M.M.R.; Quembo, C.; Nhamusso, A.; Banze, J. African swine fever in Mozambique: Review, risk factors and considerations for control. Onderstepoort J. Vet.Res. 2007, 74, 149–160. [Google Scholar] [PubMed]
- Zhou, X.; Li, N.; Luo, Y.; Liu, Y.; Miao, F.; Chen, T.; Zhang, S.; Cao, P.; Li, X.; Tian, K.; et al. Emergence of African Swine Fever in China, 2018. Transbound. Emerg. Dis. 2018, 65, 1482–1484. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Sun, Y.; Qiu, H.J. African swine fever: An unprecedented disaster and challenge to China. Infect. Dis. Poverty. 2018, 7, 111. [Google Scholar] [CrossRef]
- Halasa, T.; Botner, A.; Mortensen, S.; Christensen, H.; Toft, N.; Boklund, A. Simulating the epidemiological and economic effects of an African swine fever epidemic in industrialized swine populations. Vet. Microbiol. 2016, 193, 7–16. [Google Scholar] [CrossRef]
- Russia: Economic impact of African Swine Fever. Available online: https://www.pig333.com/latest_swine_news/russia-economic-impact-of-african-swine-fever_5019/ (accessed on 14 July 2021).
- OIE-WAHIS. Available online: https://wahis.oie.int/#/report-info?reportId=36844 (accessed on 3 August 2021).
- Rendleman, C.; Spinelli, F. An economic assessment of the costs and benefits of African Swine fever prevention. Anim. Health Insight 1994. [Google Scholar]
- Gallardo, M.C.; Reoyo, A.T.; Fernandez-Pinero, J.; Iglesias, I.; Munoz, M.J.; Arias, M.L. African swine fever: A global view of the current challenge. Porc. Health Manag. 2015, 1, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Vizcaíno, J.; Mur, L.; Gomez-Villamandos, J.; Carrasco, L. An update on the epidemiology and pathology of African swine fever. J. Comp. Pathol. 2015, 152, 9–21. [Google Scholar] [CrossRef]
- Salguero, F.J. Comparative pathology and pathogenesis of African swine fever infection in swine. Front. Vet. Sci. 2020, 7, 282. [Google Scholar] [CrossRef]
- Mebus, C.; Dardiri, A. Additional characteristics of disease caused by the African swine fever viruses isolated from Brazil and the Dominican Republic. Proc. Annu. Meet. USA Health Assoc. 1979, 83, 227–239. [Google Scholar]
- OIE. African Swine Fever Technical Disease Card. Available online: https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/AFRICAN_SWINE_FEVER.pdf (accessed on 14 July 2021).
- OIE. OIE Terrestrial Manual. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.08.01_ASF.pdf (accessed on 14 July 2021).
- Gomez-Laguna, J.; Gutierrez, A.; Pallares, F.J.; Salguero, F.J.; Ceron, J.J.; Carrasco, L. Haptoglobin and C-reactive protein as biomarkers in the serum, saliva and meat juice of pigs experimentally infected with porcine reproductive and respiratory syndrome virus. Vet. J. 2010, 185, 83–87. [Google Scholar] [CrossRef]
- Gutierrez, A.M.; Martinez-Subiela, S.; Ceron, J.J. Diagnostic accuracy of porcine acute phase proteins in meat juice for detecting disease at abattoir. Vet. Rec. 2015, 177, 15. [Google Scholar] [CrossRef]
- Ivanova, P.T.; Myers, D.S.; Milne, S.B.; McClaren, J.L.; Thomas, P.G.; Brown, H.A. Lipid composition of viral envelope of three strains of influenza virus-not all viruses are created equal. ACS Infect. Dis. 2015, 1, 399–452. [Google Scholar] [CrossRef] [Green Version]
- Meemken, D.; Tangemann, A.H.; Meermeier, D.; Gundlach, S.; Mischok, D.; Greiner, M.; Klein, G.; Blaha, T. Establishment of serological herd profiles for zoonoses and production diseases in pigs by “meat juice multi-serology”. Prev. Vet. Med. 2014, 113, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Savage, A.W.; Warriss, P.D.; Jolley, P.D. The amount and composition of the proteins in drip from stored pig meat. Meat Sci. 1990, 27, 289–303. [Google Scholar] [CrossRef]
- Di Luca, A.; Mullen, A.M.; Elia, G.; Davey, G.; Hamill, R.M. Centrifugal drip is an accessible source for protein indicators of pork ageing and water-holding capacity. Meat Sci. 2011, 88, 261–270. [Google Scholar] [CrossRef]
- Lohse, L.; Uttenthal, A.; Rasmussen, T.B.; Nielsen, J. Diagnostic value of meat juice in early detection of Classical swine fever virus infection. J. Vet. Diagn. Investig. 2011, 23, 1005–1008. [Google Scholar] [CrossRef] [Green Version]
- Kaden, V.; Lange, E.; Nagel-Kohl, U.; Bruer, W. Meat juice as diagnostic sample for virological and serological diagnosis of classical swine fever. DTW Dtsch. Tierarztl. Wochenschr. 2009, 116, 173–179. [Google Scholar]
- Yeo, S.; Yang, M.; Nyachoti, M.; Rauh, R.; Callahan, J.D.; Nfon, C. Detection of Foot-and-Mouth Disease Virus in Swine Meat Juice. Pathogens 2020, 9, 424. [Google Scholar] [CrossRef]
- De Lange, K.; Haddad, N.; Le Potier, M.F.; Agier, C.; Le Vee, M.; Amar, P.; Toma, B. Specificity of three ELISA-gE kits for screening pig meat for antibodies to Aujeszky’s disease. Vet. Rec. 2003, 153, 621–624. [Google Scholar] [CrossRef] [PubMed]
- Le Potier, M.F.; Fournier, A.; Houdayer, C.; Hutet, E.; Auvigne, V.; Hery, D.; Sanaa, M.; Toma, B. Use of muscle exudates for the detection of anti-gE antibodies to Aujeszky’s disease virus. Vet. Rec. 1998, 143, 385–387. [Google Scholar] [CrossRef] [PubMed]
- Fabisiak, M.; Podgorska, K.; Skrzypiec, E.; Szczotka, A.; Stadejek, T. Detection of porcine circovirus type 2 (PCV2) and porcine reproductive and respiratory syndrome virus (PRRSV) antibodies in meat juice samples from Polish wild boar (Sus scrofa L.). Acta Vet. Hung. 2013, 61, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Wacheck, S.; Sarno, E.; Martlbauer, E.; Zweifel, C.; Stephan, R. Seroprevalence of anti-hepatitis E virus and anti-Salmonella antibodies in pigs at slaughter in Switzerland. J. Food Prot. 2012, 75, 1483–1485. [Google Scholar] [CrossRef] [PubMed]
- Poonsuk, K.; Cheng, T.Y.; Ji, J.; Zimmerman, J.; Gimenez-Lirola, L. Detection of porcine epidemic diarrhea virus (PEDV) IgG and IgA in muscle tissue exudate (“meat juice”) specimens. Porc. Health Manag. 2018, 4, 31. [Google Scholar] [CrossRef]
- Haresnape, J.M.; Wilkinson, P.J.; Mellor, P.S. Isolation of African swine fever virus from ticks of the Ornithodoros moubata complex (Ixodoidea: Argasidae) collected within the African swine fever enzootic area of Malawi. Epidemiol. Infect. 1988, 101, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Zani, L.; Forth, J.H.; Forth, L.; Nurmoja, I.; Leidenberger, S.; Henke, J.; Carlson, J.; Breidenstein, C.; Viltrop, A.; Hoper, D.; et al. Deletion at the 5’-end of Estonian ASFV strains associated with an attenuated phenotype. Sci. Rep. 2018, 8, 6510. [Google Scholar] [CrossRef]
- Boinas, F.S.; Hutchings, G.H.; Dixon, L.K.; Wilkinson, P.J. Characterization of pathogenic and non-pathogenic African swine fever virus isolates from Ornithodoros erraticus inhabiting pig premises in Portugal. J. Gen. Virol. 2004, 85, 2177–2187. [Google Scholar] [CrossRef]
- Tignon, M.; Gallardo, C.; Iscaro, C.; Hutet, E.; Van der Stede, Y.; Kolbasov, D.; De Mia, G.M.; Le Potier, M.F.; Bishop, R.P.; Arias, M.; et al. Development and inter-laboratory validation study of an improved new real-time PCR assay with internal control for detection and laboratory diagnosis of African swine fever virus. J. Virol. Methods 2011, 178, 161–170. [Google Scholar] [CrossRef]
- Moniwa, M.; Clavijo, A.; Li, M.; Collignon, B.; Kitching, P.R. Performance of a foot-and-mouth disease virus reverse transcription-polymerase chain reaction with amplification controls between three real-time instruments. J. Vet. Diagn. Investig. 2007, 19, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Berger-Schoch, A.E.; Bernet, D.; Doherr, M.G.; Gottstein, B.; Frey, C.F. Toxoplasma gondii in Switzerland: A serosurvey based on meat juice analysis of slaughtered pigs, wild boar, sheep and cattle. Zoonoses Public Health 2011, 58, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Deksne, G.; Kirjusina, M. Seroprevalence of Toxoplasma gondii in domestic pigs (Sus scrofa domestica) and wild boars (Sus scrofa) in Latvia. J. Parasitol. 2013, 99, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Richomme, C.; Afonso, E.; Tolon, V.; Ducrot, C.; Halos, L.; Alliot, A.; Perret, C.; Thomas, M.; Boireau, P.; Gilot-Fromont, E. Seroprevalence and factors associated with Toxoplasma gondii infection in wild boar (Sus scrofa) in a Mediterranean island. Epidemiol. Infect. 2010, 138, 1257–1266. [Google Scholar] [CrossRef] [Green Version]
- Methner, U.; Rammler, N.; Fehlhaber, K.; Rosler, U. Salmonella status of pigs at slaughter--bacteriological and serological analysis. Int. J. Food Microbiol. 2011, 151, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Yonemitsu, K.; Minami, S.; Noguchi, K.; Kuwata, R.; Shimoda, H.; Maeda, K. Detection of anti-viral antibodies from meat juice of wild boars. J. Vet. Med. Sci. 2019, 81, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Petrovan, V.; Gimenez-Lirola, L.G.; Zimmerman, J.J.; Rowland, R.R.; Fang, Y.J.P. Development of a Blocking Enzyme-Linked Immunosorbent Assay for Detection of Antibodies against African Swine Fever Virus. Pathogens 2021, 10, 760. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | |||||||||||||||||||||||||
| ASFV Malawi LIL 18/2: Whole Blood and Meat Exudate Ct Values | |||||||||||||||||||||||||
| P34 3 DPI | P43 3 DPI | P33 4 DPI | P41 4 DPI | P35 5 DPI | P40 5 DPI | P38 6 DPI | P39 6 DPI | P46 6 DPI | P37 7 DPI | P42 7 DPI | |||||||||||||||
| Whole blood | 22.9 | 21.7 | 18.2 | 21.7 | 19.6 | 20.2 | 18.0 | 18.5 | 18.6 | 17.9 | 17.9 | ||||||||||||||
| Diaphragm | 29.9 | 27.9 | 24.5 | 26.6 | 25.7 | 27.0 | 24.2 | 24.7 | 25.8 | 23.1 | 23.9 | ||||||||||||||
| Biceps (L) | 33.9 | 33.5 | 28.1 | 30.2 | 30.4 | 28.2 | 25.3 | 27.2 | 28.8 | 25.0 | 26.9 | ||||||||||||||
| Biceps (R) | 34.0 | 33.5 | 27.2 | 30.8 | 28.7 | 29.3 | 25.5 | 26.8 | 28.2 | 26.4 | 27.0 | ||||||||||||||
| Brachialis (L) | 32.3 | 30.3 | 25.6 | 29.1 | 26.5 | 27.4 | 25.0 | 24.6 | 26.7 | 23.2 | 26.1 | ||||||||||||||
| Brachialis (R) | 31.9 | 28.6 | 25.5 | 27.6 | 28.1 | 27.4 | 24.7 | 26.9 | 26.1 | 23.8 | 26.6 | ||||||||||||||
| Masseter (L) | 30.5 | 28.9 | 25.8 | 26.0 | 27.8 | 27.6 | 24.1 | 26.4 | 27.9 | 24.7 | 28.1 | ||||||||||||||
| Masseter (R) | 31.5 | 28.0 | 23.8 | 28.6 | 25.9 | 26.2 | 22.4 | 26.2 | 25.8 | 22.8 | 26.8 | ||||||||||||||
| B | |||||||||||||||||||||||||
| ASFV Estonia: Whole Blood and Diaphragm Ct Values | |||||||||||||||||||||||||
| P1 3 DPI | P14 3 DPI | P30 3 DPI | P39 3 DPI | P2 4 DPI | P15 4 DPI | P27 4 DPI | P34 4 DPI | P3 5 DPI | P13 5 DPI | P29 5 DPI | P36 5 DPI | P10 7 DPI | P16 7 DPI | P28 7 DPI | P31 7 DPI | P17 8 DPI | P19 8 DPI | P8 9 DPI | P21 9 DPI | P24 9 DPI | P7 10 DPI | P32 10 DPI | P4 11 DPI | P12 11 DPI | |
| Whole blood | 26.8 | 26.7 | 24.1 | 25.7 | 24.4 | 20.5 | 23.7 | 22.2 | 21.5 | 22.5 | 20.8 | 21.3 | 23.7 | 20.0 | 19.1 | 19.7 | 21.8 | 21.3 | 19.7 | 19.3 | 19.7 | 20.3 | 24.8 | 25.2 | 20.4 |
| Diaphragm | 30.1 | 31.3 | 27.7 | 28.1 | 25.3 | 23.8 | 28.6 | 24.1 | 26.5 | 28.3 | 24.0 | 22.5 | 26.2 | 22.3 | 22.3 | 23.2 | 23.6 | 23.3 | 23.7 | 24.3 | 23.4 | 21.6 | 25.1 | 28.4 | 24.7 |
| A | |||||||||||||||||||
| P3 5 DPI | P13 5 DPI | P29 5 DPI | P36 5 DPI | P10 7 DPI | P16 7 DPI | P28 7 DPI | P31 7 DPI | P17 8 DPI | P19 8 DPI | P23 8 DPI | P25 8 DPI | P8 9 DPI | P21 9 DPI | P24 9 DPI | P7 10 DPI | P32 10 DPI | P4 11 DPI | P12 11 DPI | |
| Serum | 13.0 | 16.2 | 15.8 | 22.1 | 33.5 | 14.6 | 25.8 | 18.8 | 21.7 | 56.5 | 38.9 | 40.8 | 45.1 | 44.9 | 36.7 | 37.5 | 84.2 | 83.0 | 46.0 |
| B | |||||||||||||||||||
| P3 5 DPI | P13 5 DPI | P29 5 DPI | P36 5 DPI | P10 7 DPI | P16 7 DPI | P28 7 DPI | P31 7 DPI | P17 8 DPI | P19 8 DPI | P23 8 DPI | P25 8 DPI | P8 9 DPI | P21 9 DPI | P24 9 DPI | P7 10 DPI | P32 10 DPI | P4 11 DPI | P12 11 DPI | |
| Serum | 0.9 | 1.1 | 0.3 | 0.5 | 2.8 | 2.3 | 0.8 | 0.0 | 1.3 | 3.0 | 1.9 | 2.5 | 3.7 | 13.9 | 11.3 | 3.4 | 29.8 | 46.6 | 33.3 |
| C | |||||||||||||||||||
| Pig number | DPI 0 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 16-2 | 17-3 | 18-4 | 20-6 | 21-7 | 23-9 | 24-10 | 25-11 | 26-12 |
| 63 | 5.0 | 9.9 | 8.0 | 25.9 | 37.8 | 38.2 | 33.4 | 41.6 | 39.5 | 33.9 | 35.2 | 39.9 | 42.1 | 58.7 | 61.7 | 73.9 | 70.5 | 71.2 | |
| 64 | 10.5 | 10.4 | 13.5 | 24.8 | 26.3 | 32.9 | 19.6 | 26.1 | 19.9 | 15.9 | 13.6 | 15.1 | 17.3 | 73.6 | 74.7 | 75.4 | 69.1 | 70.1 | |
| 65 | 14.5 | 17.2 | 1.9 | 16.8 | 23.6 | 21.4 | 10.7 | 16.6 | 11.5 | 8.5 | 8.4 | 8.2 | 14.0 | 40.5 | 48.8 | 69.9 | 62.3 | 58.0 | |
| 66 | 5.6 | 9.1 | 3.8 | 24.7 | 37.6 | 37.9 | 34.4 | 42.4 | 47.3 | 43.0 | 50.3 | 58.1 | 64.5 | ||||||
| 67 | 9.8 | 15.2 | 3.1 | 14.7 | 13.1 | 17.8 | 6.5 | 10.4 | 10.2 | 5.7 | 10.4 | 7.5 | 12.6 | 54.8 | 66.5 | ||||
| 68 | 9.5 | 13.8 | 17.2 | 33.8 | 48.8 | 51.2 | 52.0 | 54.6 | 63.5 | ||||||||||
| 69 | 13.8 | 18.6 | 5.3 | 24.8 | 22.5 | 30.1 | 12.0 | 24.6 | 29.7 | 15.4 | 13.4 | 11.9 | 12.4 | 56.7 | 57.5 | 81.9 | 70.2 | 75.9 | 70.6 |
| 70 | 8.6 | 14.7 | 4.1 | 19.2 | 17.4 | 21.4 | 14.4 | 22.2 | 28.8 | 12.9 | 12.1 | 10.7 | 15.1 | 40.1 | 56.1 | 44.0 | 43.7 | 60.7 | 60.2 |
| 71 | 10.3 | 11.1 | 17.6 | 43.6 | 49.5 | 48.7 | 50.6 | 51.8 | 63.4 | ||||||||||
| 72 | 11.3 | 15.6 | 8.0 | 22.6 | 29.7 | 32.3 | 25.0 | 28.8 | 34.7 | 20.9 | 21.2 | 22.0 | 22.2 | 46.4 | 63.8 | ||||
| 73 | 11.9 | 11.5 | 7.5 | 25.0 | 27.2 | 37.3 | 46.8 | 44.4 | 49.5 | 35.9 | 25.1 | 35.3 | 44.8 | 59.6 | 63.3 | 65.0 | 62.2 | 64.9 | 62.8 |
| 74 | 13.0 | 11.4 | 3.2 | 19.5 | 19.7 | 18.6 | 19.2 | 11.6 | 26.3 | 16.1 | 16.2 | 28.3 | 39.6 | 83.4 | 85.1 | ||||
| D | |||||||||||||||||||
| Pig number | DPI 0 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 16-2 | 17-3 | 18-4 | 20-6 | 21-7 | 23-9 | 24-10 | 25-11 | 26-12 |
| 63 | 1.0 | 0.3 | 1.0 | 10.4 | 21.1 | 27.4 | 34.1 | 34.5 | 39.9 | 41.4 | 42.0 | 44.5 | 50.1 | 40.7 | 50.6 | 43.7 | 48.7 | 42.8 | |
| 64 | 0.7 | 0.4 | 0.5 | 0.2 | 1.0 | 1.2 | 1.6 | 1.5 | 1.5 | 2.4 | 9.8 | 16.1 | 27.8 | 69.6 | 81.4 | 78.3 | 84.7 | 65.2 | |
| 65 | 0.1 | 0.4 | -0.4 | 0.9 | 0.6 | 0.3 | 0.9 | 0.4 | 0.4 | 0.5 | 0.3 | 0.7 | 1.1 | 27.1 | 38.3 | 46.9 | 56.6 | 40.2 | |
| 66 | 0.6 | 0.2 | 0.8 | 3.6 | 9.7 | 15.0 | 16.7 | 28.3 | 35.7 | 50.9 | 56.8 | 56.5 | 59.4 | ||||||
| 67 | 0.7 | 0.6 | 0.4 | 0.6 | 0.8 | 0.5 | 1.2 | 1.2 | 2.2 | 2.6 | 2.4 | 2.9 | 5.6 | 51.9 | 59.6 | ||||
| 68 | 0.9 | 0.6 | 1.2 | 8.7 | 22.0 | 31.1 | 30.8 | 18.3 | 34.1 | ||||||||||
| 69 | 0.5 | 0.8 | 1.0 | 1.0 | 1.5 | 5.5 | 8.9 | 14.5 | 18.7 | 19.4 | 20.9 | 22.9 | 21.6 | 60.8 | 77.2 | 78.9 | 76.3 | 81.0 | 72.0 |
| 70 | 0.3 | 0.7 | 0.3 | 0.0 | 0.6 | 0.9 | 4.0 | 6.9 | 9.4 | 12.7 | 12.3 | 14.3 | 17.6 | 70.1 | 91.8 | 92.8 | 83.1 | 84.7 | 88.0 |
| 71 | 0.5 | 0.8 | 2.7 | 5.9 | 15.8 | 22.8 | 32.2 | 43.3 | 46.3 | ||||||||||
| 72 | 0.2 | 0.3 | 0.4 | 1.5 | 3.8 | 6.8 | 12.8 | 20.1 | 30.5 | 32.1 | 30.3 | 29.9 | 32.0 | 38.5 | 48.7 | ||||
| 73 | 0.4 | 0.5 | 0.9 | 2.8 | 9.7 | 20.5 | 27.5 | 34.7 | 34.1 | 35.1 | 32.7 | 31.5 | 35.3 | 36.9 | 36.3 | 40.9 | 40.6 | 42.1 | 39.0 |
| 74 | 0.3 | 1.0 | 1.2 | 3.4 | 8.6 | 15.4 | 20.6 | 29.3 | 30.6 | 30.6 | 34.3 | 48.4 | 60.5 | 71.0 | 68.4 | ||||
| ASFV OURT/88/3 Final DPI: ID Screen ELISA Comparison for Serum and Meat Exudate Samples | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P63 | P64 | P65 | P66 | P67 | P68 | P69 | P70 | P71 | P72 | P73 | P74 | |
| Serum | 42.8 | 65.2 | 40.2 | 59.4 | 59.6 | 34.1 | 72.0 | 88.0 | 46.3 | 48.7 | 39.0 | 68.4 |
| Diaphragm | 63.2 | 74.9 | 63.1 | 53.5 | 62.0 | 48.5 | 83.8 | 77.4 | 51.1 | 29.9 | 31.3 | 68.1 |
| Tongue | 52.8 | 72.7 | 75.8 | 60.0 | ||||||||
| Masseter (L) | 53.5 | 79.0 | 63.2 | 61.0 | 57.7 | 38.5 | 81.9 | 86.6 | 51.8 | 30.9 | 28.9 | 70.2 |
| Masseter (R) | 53.1 | 80.7 | 57.5 | 61.8 | 59.6 | 42.0 | 73.5 | 80.3 | 53.6 | 30.0 | 24.1 | 69.7 |
| Triceps (L) | 40.9 | 72.3 | 51.9 | 52.6 | ||||||||
| Triceps (R) | 41.4 | 96.6 | 54.2 | 49.6 | ||||||||
| Biceps (L) | 41.4 | 86.4 | 48.3 | 53.5 | ||||||||
| Biceps (R) | 31.3 | 82.0 | 43.8 | 62.4 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onyilagha, C.; Nash, M.; Perez, O.; Goolia, M.; Clavijo, A.; Richt, J.A.; Ambagala, A. Meat Exudate for Detection of African Swine Fever Virus Genomic Material and Anti-ASFV Antibodies. Viruses 2021, 13, 1744. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091744
Onyilagha C, Nash M, Perez O, Goolia M, Clavijo A, Richt JA, Ambagala A. Meat Exudate for Detection of African Swine Fever Virus Genomic Material and Anti-ASFV Antibodies. Viruses. 2021; 13(9):1744. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091744
Chicago/Turabian StyleOnyilagha, Chukwunonso, Mikyla Nash, Orlando Perez, Melissa Goolia, Alfonso Clavijo, Juergen A. Richt, and Aruna Ambagala. 2021. "Meat Exudate for Detection of African Swine Fever Virus Genomic Material and Anti-ASFV Antibodies" Viruses 13, no. 9: 1744. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091744