
Potential Diagnostic and Prognostic Biomarkers for Adenovirus Respiratory Infection in Children and Young Adults

 , ,
, ,  , , ,
, , ,  , and
, and
Abstract
:1. Introduction
1.1. Viral Structure and Classification
1.2. Epidemiology and Transmission
1.3. Clinical Features
1.4. Adenovirus Persistence and Reactivation
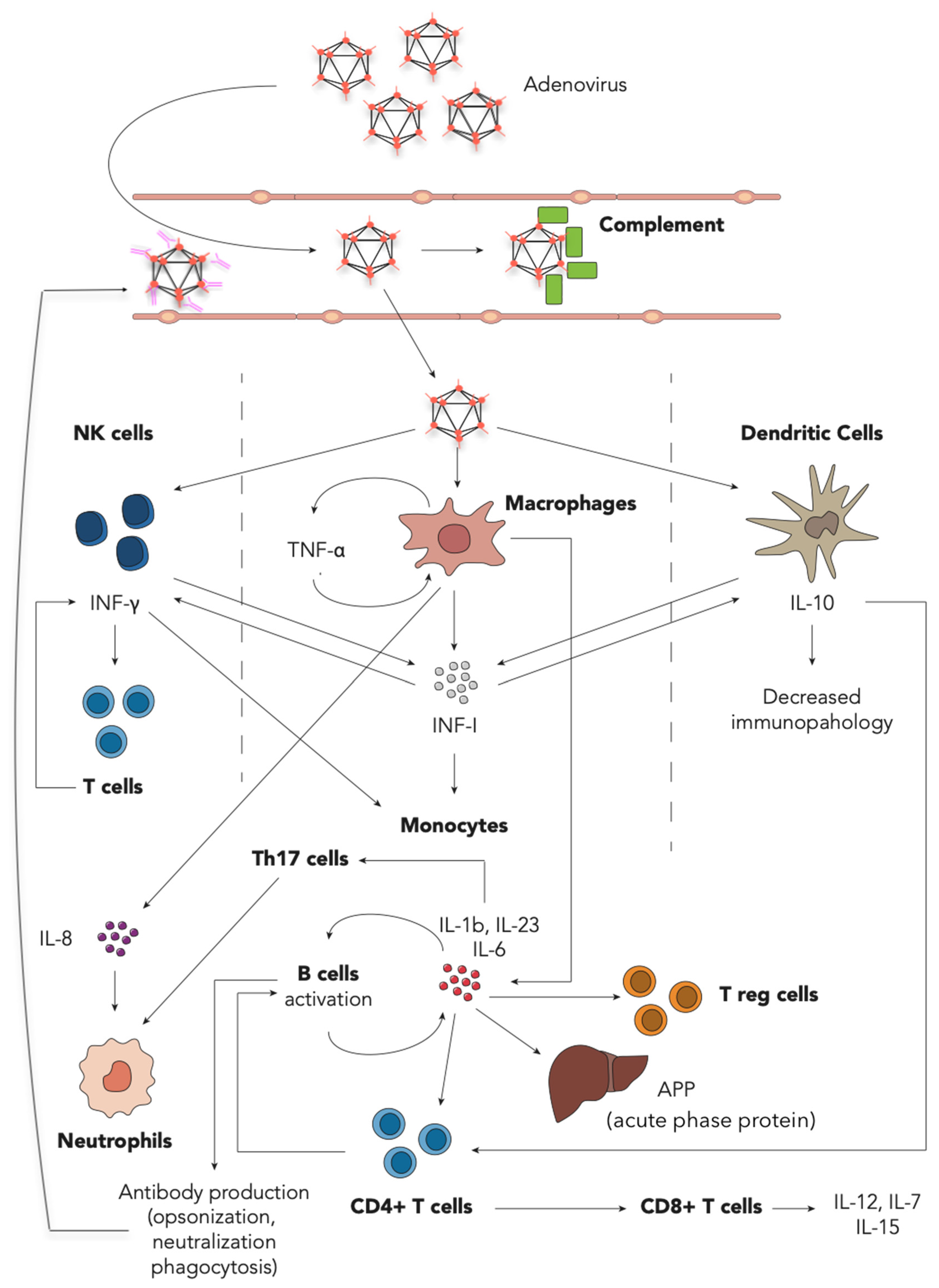
1.5. Immune Response
1.6. Diagnosis of Adenoviral Infection and Disease
1.7. Treating HAdV Infection
1.8. Potential Markers Based on Immunology
1.8.1. Hematological Changes
1.8.2. Cytokine Changes
2. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
| Authors | Kit Used | Analyzer and Software | Notes |
|---|---|---|---|
| Nakamura et al. [69] | MILLIPLEX MAP Human Cytokine/Chemokine Kit (Millipore, Billerica, MA, USA) | Luminex 100 TM analyzer (Luminex, Tokyo, Japan) | - |
| Fuchs et al. [70] | Human Cytokine/Chemokine Magnetic Bead Panel (Milliplex MAP kit Cat. # HCYTOMAG-60 K, Millipore Corp., Billerica, MA, USA) | Magpix Luminex (Luminex Corporation, Austin, TX, USA) and xponent software (version 4.2, Luminex Corp, Austin, TX, USA) | - |
| Moro et al. [99] | 25-plex bead immunoassay kit (Biosource International, Camarillo, CA, USA) | Luminex 100 IS xMap multiplex system (Luminex Corporation, Austin, TX, USA) and Star Station V. 2.0 software (Applied Cytometry Systems, Sacramento, CA, USA) | Samples were stored at −80 °C until use, thawed at room temperature, and diluted 1:1 before analysis, according to the manufacturer’s instructions |
| Lim et al. [84] | Quantikine HS Human IL-6 Immunoassay, HS Human TNF-α, Human CXCL8/IL-8 Immunoassay, Quantikine Human IFN-γ and Human IL-10 Quant HS. ELISA kit, R&D, Minneapolis, MN, USA). | - | Assays were performed following the manufacturer’s instructions |
| Kawasaki et al. [72] | Enzyme-linked immunosorbent assay kits from Endogen (Endogen, Inc., Woburn, MA, USA) | - | - |
| Chen et al. [83] | Human cytokine kit (Milliplex, Catalog no. MPXHCYTO-60 K-16, Millipore, Billerica, MA, USA) | FLEXMAP 3D system (Luminex, Austin, TX, USA) | Assays were performed following the manufacturer’s instructions |
| Kleiner et al. [94] | Magnetic bead-based multiplex immunoassays (Bio-Plex) (BIO-RAD Laboratories, Milano, Italy) | Bio-Plex 200 reader and Bio-Plex Manager software | - |
| Biserni et al. [75] | MILLIPLEX MAP Human Cytokine/Chemokine Kit (Millipore, Billerica, MA, USA) | Luminex 100 TM analyzer (Luminex, Austin, TX, USA) | - |
| Mistchenko et al. [100] | ELISA kits for Human Cytokines from R&D Systems (Minneapolis, MN, USA) | - | - |
| Sun et al. [73] | - | - | Retrospective collection of IL-6 concentration |
| Nakamura et al. [69] | MILLIPLEX MAP Human Cytokine/Chemokine Kit (Millipore, Billerica, MA, USA) | Luminex 100 TM analyzer (Luminex, Tokyo, Japan) |
References
- Van Houten, C.B.; Cohen, A.; Engelhard, D.; Hays, J.P.; Karlsson, R.; Moore, E.; Fernández, D.; Kreisberg, R.; Collins, L.V.; de Waal, W.; et al. Antibiotic misuse in respiratory tract infections in children and adults-a prospective, multicentre study (TAILORED Treatment). Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Dadgostar, P. Antimicrobial Resistance: Implications and Costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- L’Huillier, A.G.; Mardegan, C.; Cordey, S.; Luterbacher, F.; Papis, S.; Hugon, F.; Kaiser, L.; Gervaix, A.; Posfay-Barbe, K.; Galetto-Lacour, A. Enterovirus, parechovirus, adenovirus and herpes virus type 6 viraemia in fever without source. Arch. Dis. Child. 2020, 105, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Nemerow, G.R.; Stewart, P.L.; Reddy, V.S. Structure of human adenovirus. Curr. Opin. Virol. 2012, 2, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Morris, S.J.; David, W.D.; Donald, S.; James, C. Homologous recombination in E3 genes of human adenovirus species D. J. Virol. 2013, 87, 12481–12488. [Google Scholar] [CrossRef] [Green Version]
- Walsh, M.P.; Seto, J.; Jones, M.S.; Chodosh, J.; Xu, W.; Seto, D. Computational analysis identifies human adenovirus type 55 as a re-emergent acute respiratory disease pathogen. J. Clin. Microbiol. 2010, 48, 991–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lion, T. Adenovirus infections in immunocompetent and immunocompromised patients. Clin. Microbiol. Rev. 2014, 27, 441–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, J.P., 3rd; Kajon, A.E. Adenovirus: Epidemiology, Global Spread of Novel Serotypes, and Advances in Treatment and Prevention. Semin. Respir. Crit. Care Med. 2016, 37, 586–602. [Google Scholar]
- Lion, T. Adenovirus persistence, reactivation, and clinical management. FEBS Lett. 2019, 593, 3571–3582. [Google Scholar] [CrossRef] [Green Version]
- Khanal, S.; Ghimire, P.; Dhamoon, A.S. The Repertoire of Adenovirus in Human Disease: The Innocuous to the Deadly. Biomedicines 2018, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Binder, A.M.; Biggs, H.M.; Haynes, A.K.; Chommanard, C.; Lu, X.; Erdman, D.D.; Watson, J.T.; Gerber, S.I. Human Adenovirus Surveillance—United States, 2003–2016. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 1039–1042. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhou, J.N.; Li, H.; He, C.Y.; Dai, Q.S.; Li, X.L.; He, J.F.; He, H.; Li, M.B.; Jiang, L.I.; et al. An outbreak of epidemic keratoconjunctivitis caused by human adenovirus type 8 in primary school, southwest China. BMC Infect. Dis. 2019, 19, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akello, J.O.; Kamgang, R.; Barbani, M.T.; Suter-Riniker, F.; Leib, S.L.; Ramette, A. Epidemiology of Human Adenoviruses: A 20-Year Retrospective Observational Study in Hospitalized Patients in Bern, Switzerland. Clin. Epidemiol. 2020, 12, 353–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiroi, S.; Morikawa, S.; Nakata, K.; Kase, T. Surveillance of Adenovirus Respiratory Infections in Children from Osaka, Japan. Jpn. J. Infect. Dis. 2017, 70, 666–668. [Google Scholar] [CrossRef] [Green Version]
- Sammons, J.S.; Graf, E.H.; Townsend, S.; Hoegg, C.L.; Smathers, S.A.; Coffin, S.E.; Williams, K.; Mitchell, S.L.; Nawab, U.; Munson, D.; et al. Outbreak of Adenovirus in a Neonatal Intensive Care Unit: Critical Importance of Equipment Cleaning During Inpatient Ophthalmologic Examinations. Ophthalmology 2019, 126, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Ison, M.G.; Hayden, R.T. Adenovirus. Microbiol. Spectr. 2016, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Alharbi, S.; Van Caeseele, P.; Consunji-Araneta, R.; Zoubeidi, T.; Fanella, S.; Souid, A.K.; Alsuwaidi, A.R. Epidemiology of severe pediatric adenovirus lower respiratory tract infections in Manitoba, Canada, 1991–2005. BMC Infect. Dis. 2012, 12, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocholl, C.; Gerber, K.; Daly, J.; Pavia, A.T.; Byington, C.L. Adenoviral infections in children: The impact of rapid diagnosis. Pediatrics 2004, 113, e51–e56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, Y.F.; Koh, W.V.; Kan, C.; Dua, P.A.; Lim, A.E.; Liaw, C.J.; Gao, Q.H.; Chng, J.; Lee, V.J.; Tan, B.H.; et al. Epidemiologic analysis of respiratory viral infections among Singapore military servicemen in 2016. BMC Infect. Dis. 2018, 18, 123. [Google Scholar] [CrossRef] [Green Version]
- Gray, G.C.; McCarthy, T.; Lebeck, M.G.; Schnurr, D.P.; Russell, K.L.; Kajon, A.E.; Landry, M.L.; Leland, D.S.; Storch, G.A.; Ginocchio, C.C.; et al. Genotype prevalence and risk factors for severe clinical adenovirus infection, United States 2004–2006. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2007, 45, 1120–1131. [Google Scholar] [CrossRef] [Green Version]
- Al Qurashi, Y.M.; Guiver, M.; Cooper, R.J. Sequence typing of adenovirus from samples from hematological stem cell transplant recipients. J. Med. Virol. 2011, 83, 1951–1958. [Google Scholar] [CrossRef]
- Schwartz, K.L.; Richardson, S.E.; MacGregor, D.; Mahant, S.; Raghuram, K.; Bitnun, A. Adenovirus-Associated Central Nervous System Disease in Children. J. Pediatr. 2019, 205, 130–137. [Google Scholar] [CrossRef]
- Shi, T.; Chen, C.; Huang, L.; Fan, H.; Lu, G.; Yang, D.; Zhao, C.; Zhang, D. Risk factors for mortality from severe community-acquired pneumonia in hospitalized children transferred to the pediatric intensive care unit. Pediatr. Neonatol. 2020, 61, 577–583. [Google Scholar] [CrossRef]
- Scott, M.K.; Chommanard, C.; Lu, X.; Appelgate, D.; Grenz, L.; Schneider, E.; Gerber, S.I.; Erdman, D.D.; Thomas, A. Human Adenovirus Associated with Severe Respiratory Infection, Oregon, USA, 2013–2014. Emerg. Infect. Dis. 2016, 22, 1044–1051. [Google Scholar] [CrossRef] [Green Version]
- Shachor-Meyouhas, Y.; Hadash, A.; Kra-Oz, Z.; Shafran, E.; Szwarcwort-Cohen, M.; Kassis, I. Adenovirus Respiratory Infection among Immunocompetent Patients in a Pediatric Intensive Care Unit During 10-year period: Co-morbidity is common. Isr. Med. Assoc. J. IMAJ 2019, 21, 595–598. [Google Scholar]
- Santos, C.A.Q.; Rhee, Y.; Czapka, M.T.; Kazi, A.S.; Proia, L.A. Make Sure You Have a Safety Net: Updates in the Prevention and Management of Infectious Complications in Stem Cell Transplant Recipients. J. Clin. Med. 2020, 9, 865. [Google Scholar] [CrossRef] [Green Version]
- Hierholzer, J.C. Adenoviruses in the immunocompromised host. Clin. Microbiol. Rev. 1992, 5, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Hum, R.M.; Deambrosis, D.; Lum, S.H.; Davies, E.; Bonney, D.; Guiver, M.; Turner, A.; Wynn, R.F.; Hiwarkar, P. Molecular monitoring of adenovirus reactivation in faeces after haematopoietic stem-cell transplantation to predict systemic infection: A retrospective cohort study. Lancet Haematol. 2018, 5, e422–e429. [Google Scholar] [CrossRef]
- Proenca-Modena, J.L.; de Souza Cardoso, R.; Criado, M.F.; Milanez, G.P.; de Souza, W.M.; Parise, P.L.; Bertol, J.W.; de Jesus, B.L.S.; Prates, M.C.M.; Silva, M.L.; et al. Human adenovirus replication and persistence in hypertrophic adenoids and palatine tonsils in children. J. Med. Virol. 2019, 91, 1250–1262. [Google Scholar] [CrossRef] [PubMed]
- Wurzel, D.F.; Marchant, J.M.; Yerkovich, S.T.; John, W.; Ian, M.; Brent Masters, I.; Anne, B.C. Prospective characterization of protracted bacterial bronchitis in children. Chest 2014, 145, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Colom, A.J.; Teper, A.M. Post-infectious bronchiolitis obliterans. Pediatr. Pulmonol. 2019, 54, 212–219. [Google Scholar] [PubMed]
- Kälin, S.; Amstutz, B.; Gastaldelli, M.; Wolfrum, N.; Boucke, K.; Havenga, M.; DiGennaro, F.; Liska, N.; Hemmi, S.; Greber, U.F. Macropinocytotic uptake and infection of human epithelial cells with species B2 adenovirus type 35. J. Virol. 2010, 84, 5336–5350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greber, U.F.; Flatt, J.W. Adenovirus Entry: From Infection to Immunity. Annu. Rev. Virol. 2019, 6, 177–197. [Google Scholar] [CrossRef]
- Luisoni, S.; Suomalainen, M.; Boucke, K.; Tanner, L.B.; Wenk, M.R.; Guan, X.L.; Grzybek, M.; Coskun, Ü.; Greber, U.F. Co-option of Membrane Wounding Enables Virus Penetration into Cells. Cell Host Microbe 2015, 18, 75–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.G.; Nemerow, G.R. Mechanism of adenovirus neutralization by Human alpha-defensins. Cell Host Microbe 2008, 3, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.G.; Silvestry, M.; Lindert, S.; Lu, W.; Nemerow, G.R.; Stewart, P.L. Insight into the mechanisms of adenovirus capsid disassembly from studies of defensin neutralization. PLoS Pathog. 2010, 6, e1000959. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Tian, J.; Smith, J.S.; Byrnes, A.P. Clearance of adenovirus by Kupffer cells is mediated by scavenger receptors, natural antibodies, and complement. J. Virol. 2008, 82, 11705–11713. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, A.J.; Mallery, D.L.; Watkinson, R.E.; Dickson, C.F.; James, L.C. Sequential ubiquitination and deubiquitination enzymes synchronize the dual sensor and effector functions of TRIM21. Proc. Natl. Acad. Sci. USA 2015, 112, 10014–10019. [Google Scholar] [CrossRef] [Green Version]
- Di Paolo, N.C.; Baldwin, L.K.; Irons, E.E.; Papayannopoulou, T.; Tomlinson, S.; Shayakhmetov, D.M. IL-1α and complement cooperate in triggering local neutrophilic inflammation in response to adenovirus and eliminating virus-containing cells. PLoS Pathog. 2014, 10, e1004035. [Google Scholar] [CrossRef]
- Di Paolo, N.C.; Miao, E.A.; Iwakura, Y.; Murali-Krishna, K.; Aderem, A.; Flavell, R.A.; Papayannopoulou, T.; Shayakhmetov, D.M. Virus binding to a plasma membrane receptor triggers interleukin-1 alpha-mediated proinflammatory macrophage response in vivo. Immunity 2009, 31, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Maler, M.D.; Nielsen, P.J.; Stichling, N.; Cohen, I.; Ruzsics, Z.; Wood, C.; Engelhard, P.; Suomalainen, M.; Gyory, I.; Huber, M.; et al. Key Role of the Scavenger Receptor MARCO in Mediating Adenovirus Infection and Subsequent Innate Responses of Macrophages. MBio 2017, 8, e00670-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labzin, L.I.; Bottermann, M.; Rodriguez-Silvestre, P.; Foss, S.; Andersen, J.T.; Vaysburd, M.; Clift, D.; James, L.C. Antibody and DNA sensing pathways converge to activate the inflammasome during primary human macrophage infection. EMBO J. 2019, 38, e101365. [Google Scholar] [CrossRef]
- Guidotti, L.G.; Chisari, F.V. Noncytolytic control of viral infections by the innate and adaptive immune response. Annu. Rev. Immunol. 2001, 19, 65–91. [Google Scholar] [CrossRef] [PubMed]
- Schoenberger, S.P.; Toes, R.E.; van der Voort, E.I.; Offringa, R.; Melief, C.J. T-cell help for cytotoxic T lymphocytes is mediated by CD40-CD40L interactions. Nature 1998, 393, 480–483. [Google Scholar] [CrossRef] [PubMed]
- Heemskerk, B.; Van Vreeswijk, T.; Veltrop-Duits, L.A.; Sombroek, C.C.; Franken, K.; Verhoosel, R.M.; Hiemstra, P.; Van Leeuwen, D.; Ressing, M.E.; Toes, R.; et al. Adenovirus-specific CD4+ T cell clones recognizing endogenous antigen inhibit viral replication in vitro through cognate interaction. J. Immunol. 2006, 177, 8851–8859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leen, A.M.; Christin, A.; Khalil, M.; Weiss, H.; Gee, A.P.; Brenner, M.K.; Heslop, H.E.; Rooney, C.M.; Bollard, C.M. Identification of hexon-specific CD4 and CD8 T-cell epitopes for vaccine and immunotherapy. J. Virol. 2008, 82, 546–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutnick, N.A.; Carnathan, D.; Demers, K.; Makedonas, G.; Ertl, H.C.; Betts, M.R. Adenovirus-specific human T cells are pervasive, polyfunctional, and cross-reactive. Vaccine 2010, 28, 1932–1941. [Google Scholar] [CrossRef] [Green Version]
- Veltrop-Duits, L.A.; Van Vreeswijk, T.; Heemskerk, B.; Thijssen, J.C.P.; El Seady, R.; Der Zijde, E.M.J.-V.; Claas, E.C.J.; Lankester, A.C.; Van Tol, M.J.D.; Schilham, M.W. High titers of pre-existing adenovirus serotype-specific neutralizing antibodies in the host predict viral reactivation after allogeneic stem cell transplantation in children. Clin. Infect. Dis. 2011, 52, 1405–1413. [Google Scholar] [CrossRef]
- Hirsch, H.H.; Martino, R.; Ward, K.N.; Boeckh, M.; Einsele, H.; Ljungman, P. Fourth European Conference on Infections in Leukaemia (ECIL-4): Guidelines for diagnosis and treatment of human respiratory syncytial virus, parainfluenza virus, metapneumovirus, rhinovirus, and coronavirus. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2013, 56, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Florescu, D.F.; Stohs, E.J. Approach to infection and disease due to adenoviruses in solid organ transplantation. Curr. Opin. Infect. Dis. 2019, 32, 300–306. [Google Scholar] [CrossRef]
- Zhang, Q.; Jing, S.; Cheng, Z.; Yu, Z.; Dehghan, S.; Shamsaddini, A.; Yan, Y.; Li, M.; Seto, D. Comparative genomic analysis of two emergent human adenovirus type 14 respiratory pathogen isolates in China reveals similar yet divergent genomes. Emerg. Microbes. Infect. 2017, 1, 6–e92. [Google Scholar] [CrossRef] [Green Version]
- Wallot, M.A.; Dohna-Schwake, C.; Auth, M.; Nadalin, S.; Fiedler, M.; Malagó, M.; Broelsch, C.; Voit, T. Disseminated adenovirus infection with respiratory failure in pediatric liver transplant recipients: Impact of intravenous cidofovir and inhaled nitric oxide. Pediatr. Transplant. 2006, 10, 121–127. [Google Scholar] [CrossRef]
- Zhang, Y.; Sakthivel, S.K.; Bramley, A.; Jain, S.; Haynes, A.; Chappell, J.D.; Hymas, W.; Lenny, N.; Patel, A.; Qi, C.; et al. Serology Enhances Molecular Diagnosis of Respiratory Virus Infections Other than Influenza in Children and Adults Hospitalized with Community-Acquired Pneumonia. J. Clin. Microbiol. 2016, 55, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Lion, T.; Kosulin, K.; Landlinger, C.; Rauch, M.; Preuner, S.; Jugovic, D.; Pötschger, U.; Lawitschka, A.; Peters, C.; Fritsch, G.; et al. Monitoring of adenovirus load in stool by real-time PCR permits early detection of impending invasive infection in patients after allogeneic stem cell transplantation. Leukemia 2010, 24, 706–714. [Google Scholar] [CrossRef]
- Jeulin, H.; Salmon, A.; Bordigoni, P.; Venard, V. Diagnostic value of quantitative PCR for adenovirus detection in stool samples as compared with antigen detection and cell culture in haematopoietic stem cell transplant recipients. Clin. Microbiol. Infect. 2011, 17, 1674–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morozumi, M.; Shimizu, H.; Matsushima, Y.; Mitamura, K.; Tajima, T.; Iwata, S.; Ubukata, K. Evaluation of new immunochromatographic assay kit for adenovirus detection in throat swab: Comparison with culture and real-time PCR results. J. Infect. Chemother. Off. J. Jpn. Soc. Chemother. 2014, 20, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, J.; Wright, N.; Ferrenberg, J.; Huang, M.L.; Cent, A.; Corey, L.; Morrow, R. Comparison of real-time PCR assays with fluorescent-antibody assays for diagnosis of respiratory virus infections in children. J. Clin. Microbiol. 2006, 44, 2382–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.Y.; Hwang, D.; Chiu, N.C.; Weng, L.C.; Liu, H.F.; Mu, J.J.; Liu, C.P.; Chi, H. Increased Detection of Viruses in Children with Respiratory Tract Infection Using PCR. Int. J. Environ. Res. Public Health 2020, 17, 564. [Google Scholar] [CrossRef] [Green Version]
- Shimada, Y.; Ariga, T.; Tagawa, Y.; Aoki, K.; Ohno, S.; Ishiko, H. Molecular diagnosis of human adenoviruses d and e by a phylogeny-based classification method using a partial hexon sequence. J. Clin. Microbiol. 2004, 42, 1577–1584. [Google Scholar] [CrossRef] [Green Version]
- Ebner, K.; Suda, M.; Watzinger, F.; Lion, T. Molecular detection and quantitative analysis of the entire spectrum of human adenoviruses by a two-reaction real-time PCR assay. J. Clin. Microbiol. 2005, 43, 3049–3053. [Google Scholar] [CrossRef] [Green Version]
- Dou, Y.; Li, Y.; Ma, C.; Zhu, H.; Du, J.; Liu, H.; Liu, Q.; Chen, R.; Tan, Y. Rapid diagnosis of human adenovirus B, C and E in the respiratory tract using multiplex quantitative polymerase chain reaction. Mol. Med. Rep. 2018, 18, 2889–2897. [Google Scholar] [CrossRef] [Green Version]
- Hanson, K.E.; Azar, M.M.; Banerjee, R.; Chou, A.; Colgrove, R.C.; Ginocchio, C.C.; Hayden, M.K.; Holodiny, M.; Jain, S.; Koo, S.; et al. Molecular testing for acute respiratory tract infections: Clinical and diagnostic recommendations from the IDSA’s Diagnostics Committee. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2020, 71, 2744–2751. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.S.; Tsai, C.L.; Chang, J.; Hsu, T.C.; Lin, S.; Lee, C.C. Multiplex PCR system for the rapid diagnosis of respiratory virus infection: Systematic review and meta-analysis. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2018, 24, 1055–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulini, I.; Siqueira-Silva, J.; Thomaz, L.; Rocha, L.; Harsi, C.; Bellei, N.; Granato, C. Development of a prototype immunochromatographic test for rapid diagnosis of respiratory adenovirus infection. Braz. J. Infect. Dis. Off. Publ. Braz. Soc. Infect. Dis. 2017, 21, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Huijskens, E.G.; Biesmans, R.C.; Buiting, A.G.; Obihara, C.C.; Rossen, J. W Diagnostic value of respiratory virus detection in symptomatic children using real-time PCR. Virol. J. 2012, 9, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthes-Martin, S.; Feuchtinger, T.; Shaw, P.J.; Engelhard, D.; Hirsch, H.H.; Cordonnier, C.; Ljungman, P. Fourth European Conference on Infections in LeukemiaEuropean guidelines for diagnosis and treatment of adenovirus infection in leukemia and stem cell transplantation: Summary of ECIL-4 (2011). Transpl. Infect. Dis. 2012, 14, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Kodama, E.; Shigeta, S.; Suzuki, T.; De Clercq, E. Application of a gastric cancer cell line (MKN-28) for anti-adenovirus screening using the MTT method. Antivir. Res. 1996, 31, 159–164. [Google Scholar] [CrossRef]
- Lugthart, G.; Oomen, M.A.; Jol-van der Zijde, C.M.; Ball, L.M.; Bresters, D.; Kollen, W.J.W.; Smiers, F.J.; Vermont, C.L.; Bredius, R.G.M.; Schilham, M.W. The effect of cidofovir on adenovirus plasma DNA levels in stem cell transplantation recipients without T cell reconstitution. Biol. Blood Marrow. Transplant. 2015, 21, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, H.; Fujisawa, T.; Suga, S.; Taniguchi, K.; Nagao, M.; Ito, M.; Ochiai, H.; Konagaya, M.; Hanaoka, N.; Fujimoto, T. Species differences in circulation and inflammatory responses in children with common respiratory adenovirus infections. J. Med. Virol. 2018, 90, 873–880. [Google Scholar] [CrossRef]
- Fuchs, A.; Gotta, V.; Decker, M.L.; Szinnai, G.; Baumann, P.; Bonhoeffer, J.; Ritz, N.; ProPAED study group. Cytokine kinetic profiles in children with acute lower respiratory tract infection: A post hoc descriptive analysis from a randomized control trial. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2018, 24, 1341.e1–1341.e7. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.J.; Rhie, Y.J.; Seo, W.H.; Jang, G.Y.; Choi, B.M.; Lee, J.H.; Lee, C.K.; Kim, Y.K. Clinical manifestations of respiratory adenoviral infection among hospitalized children in Korea. Pediatr. Int. 2013, 55, 450–454. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Hosoya, M.; Katayose, M.; Suzuki, H. Correlation between serum interleukin 6 and C-reactive protein concentrations in patients with adenoviral respiratory infection. Pediatr. Infect. Dis. J. 2002, 21, 370–374. [Google Scholar] [CrossRef]
- Sun, J.; Xiao, Y.; Zhang, M.; Ao, T.; Lang, S.; Wang, J. Serum Inflammatory Markers in Patients with Adenovirus Respiratory Infection. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 3848–3855. [Google Scholar] [CrossRef] [PubMed]
- Wald, E.R.; Applegate, K.E.; Bordley, C.; Darrow, D.H.; Glode, M.P.; Marcy, S.M.; Nelson, C.E.; Rosenfeld, R.M.; Shaikh, N.; Smith, M.J.; et al. Clinical practice guideline for the diagnosis and management of acute bacterial sinusitis in children aged 1 to 18 years. Pediatrics 2013, 132, e262–e280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biserni, G.B.; Dondi, A.; Masetti, R.; Bandini, J.; Dormi, A.; Conti, F.; Pession, A.; Lanari, M. Immune Response against Adenovirus in Acute Upper Respiratory Tract Infections in Immunocompetent Children. Vaccines 2020, 8, 602. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Zampiero, A.; Bianchini, S.; Mori, A.; Scala, A.; Tagliabue, C.; Sciarrabba, C.S.; Fossali, E.; Piralla, A.; Principi, N. Epidemiology and Clinical Characteristics of Respiratory Infections Due to Adenovirus in Children Living in Milan, Italy, during 2013 and 2014. PLoS ONE 2016, 11, e0152375. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.F.; Shen, F.C.; Wang, S.L.; Kuo, P.H.; Tsai, H.P.; Liu, C.C.; Wang, J.R.; Chi, C.Y. Molecular Epidemiology and Clinical Manifestations of Adenovirus Respiratory Infections in Taiwanese Children. Medicine 2016, 95, e3577. [Google Scholar] [CrossRef]
- Yang, X.; Wang, Q.; Liang, B.; Wu, F.; Li, H.; Liu, H.; Sheng, C.; Ma, Q.; Yang, C.; Xie, J.; et al. An outbreak of acute respiratory disease caused by a virus associated RNA II gene mutation strain of human adenovirus 7 in China, 2015. PLoS ONE 2017, 12, e0172519. [Google Scholar] [CrossRef]
- Noyola, E.; Noor, A.; Sweeney, N.; Chan, J.; Ramesh, R.; Calixte, R.; Krilov, L.R. Prevalence of Bandemia in Respiratory Viral Infections: A Pediatric Emergency Room Experience. Front. Pediatr. 2020, 8, 576676. [Google Scholar] [CrossRef]
- Li, K.; Li, L.; Wang, X.; Li, H.; Chen, J.; Liu, L.; Shao, J.; Xu, Y.; He, L.; Gong, S.; et al. Comparative analysis of clinical features of SARS-CoV-2 and adenovirus infection among children. Virol. J. 2020, 17, 193. [Google Scholar] [CrossRef]
- Tang, Y.; Li, X.; Cao, L.; Chen, Y.; Yan, W.; Xu, Q.; Lv, H. Characteristics and Indications of Kawasaki Disease Among Infants Under 6 Months. Front. Pediatr. 2020, 8, 470. [Google Scholar] [CrossRef]
- Dallman, P. Pediatrics, 16th ed.; Appleton-Century-Crofts: New York, NY, USA, 1977. [Google Scholar]
- Chen, W.W.; Nie, W.M.; Xu, W.; Xie, Y.X.; Tu, B.; Zhao, P.; Qin, E.Q.; Zhang, Y.H.; Zhang, X.; Li, W.G.; et al. Cross-sectional study of the relationship of peripheral blood cell profiles with severity of infection by adenovirus type 55. BMC Infect. Dis. 2014, 14, 147. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.U.; Choi, J.Y.; Jeong, H.J.; Ko, J.H.; Lee, J.E.; Rhee, C.K. Comparison of clinical characteristics and inflammatory cytokines between hypoxemic and non-hypoxemic human adenovirus 55 pneumonia. J. Thorac. Dis. 2020, 12, 4044–4056. [Google Scholar] [CrossRef]
- Yoon, H.; Jhun, B.W.; Kim, S.J.; Kim, K. Clinical characteristics and factors predicting respiratory failure in adenovirus pneumonia. Respirology 2016, 21, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Zsengellér, Z.; Otake, K.; Hossain, S.A.; Berclaz, P.Y.; Trapnell, B.C. Internalization of adenovirus by alveolar macrophages initiates early proinflammatory signaling during acute respiratory tract infection. J. Virol. 2000, 74, 9655–9667. [Google Scholar] [CrossRef] [Green Version]
- Helmy, K.Y.; Katschke KJJr Gorgani, N.N.; Kljavin, N.M.; Elliott, J.M.; Diehl, L.; Scales, S.J.; Ghilardi, N.; Campagne, M.V. CRIg: A macrophage complement receptor required for phagocytosis of circulating pathogens. Cell 2006, 124, 915–927. [Google Scholar] [CrossRef] [Green Version]
- Tam, J.C.; Bidgood, S.R.; McEwan, W.A.; James, L.C. Intracellular sensing of complement C3 activates cell autonomous immunity. Science 2014, 345, 1256070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottermann, M.; Foss, S.; Caddy, S.L.; Clift, D.; van Tienen, L.M.; Vaysburd, M.; Cruickshank, J.; O’Connell, K.; Clark, J.; Mayes, K.; et al. Complement C4 Prevents Viral Infection through Capsid Inactivation. Cell Host Microbe 2019, 25, 617–629.e7. [Google Scholar] [CrossRef] [Green Version]
- Arnberg, N. Adenovirus receptors: Implications for tropism, treatment and targeting. Rev. Med. Virol. 2009, 19, 165–178. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Parkin, J.; Cohen, B. An overview of the immune system. Lancet 2001, 357, 1777–1789. [Google Scholar] [CrossRef]
- Simon, A.K.; Hollander, G.A.; McMichael, A. Evolution of the immune system in humans from infancy to old age. Proc. Biol. Sci. 2015, 282, 20143085. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, G.; Marcuzzi, A.; Zanin, V.; Monasta, L.; Zauli, G. Cytokine levels in the serum of healthy subjects. Mediat. Inflamm. 2013, 2013, 434010. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Li, C.J.; Long, Y.; Sun, D.M.; Wang, R.G.; Zhang, Y. Clinical features of children with severe adenovirus pneumonia and hemophagocytic syndrome: An analysis of 30 case. Chin. J. Contemp. Pediatr. 2020, 22, 744–748. [Google Scholar]
- Yang, T.I.; Li, W.L.; Chang, T.H.; Lu, C.Y.; Chen, J.M.; Lee, P.I.; Huang, L.M.; Chang, L.Y. Adenovirus replication and host innate response in primary human airway epithelial cells. Journal of microbiology, immunology, and infection. Wei Mian Yu Gan Ran Za Zhi 2019, 52, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Mihara, M.; Hashizume, M.; Yoshida, H.; Suzuki, M.; Shiina, M. IL-6/IL-6 receptor system and its role in physiological and pathological conditions. Clin. Sci. 2012, 122, 143–159. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Moro, M.R.; Bonville, C.A.; Suryadevara, M.; Cummings, E.; Faddoul, D.; Kobayaa, H.; Branigan, P.J.; Domachowske, J.B. Clinical features, adenovirus types, and local production of inflammatory mediators in adenovirus infections. Pediatr. Infect. Dis. J. 2009, 28, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Mistchenko, A.S.; Diez, R.A.; Mariani, A.L.; Robaldo, J.; Maffey, A.F.; Bayley-Bustamante, G.; Grinstein, S. Cytokines in adenoviral disease in children: Association of interleukin-6, interleukin-8, and tumor necrosis factor alpha levels with clinical outcome. J. Pediatr. 1994, 124, 714–720. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, A. Dendritic Cell-Airway Epithelial Cell Cross-Talk Changes with Age and Contributes to Chronic Lung Inflammatory Diseases in the Elderly. Int. J. Mol. Sci. 2017, 18, 1206. [Google Scholar] [CrossRef] [Green Version]

| Localization | Clinical Manifestations | Most Affected Groups | Main Serotypes |
|---|---|---|---|
| Respiratory tract | Upper respiratory tract infection Lower respiratory tract infection | Pediatric patients Adult patients Outbreaks in hospitals, chronic care facilities, closed settings (e.g., military recruits) | Types 1–5, 7, 14, and 21 associated with small airways dysfunction and bronchiectasis in children, chronic obstructive pulmonary disease in adults |
| Gastrointestinal tract | Gastroenteritis, diarrhea Rare complications: hemorrhagic colitis, hepatitis, cholecystitis, pancreatitis | Young children Outbreaks in hospitals, chronic care facilities, closed settings (e.g., military recruits), patients with HIV [7] | Types 40 and 41 |
| Eye | Epidemic keratoconjunctivitis Pharyngoconjunctival fever Non-specific conjunctivitis | Outbreaks in hospitals, chronic care facilities, closed settings (e.g., military recruits) | Types 8, 19, 37 (most common) Types 3, 4, 7, 11, 14 (less common) Types 53, 54, 56 associated with outbreaks [20] |
| Urinary tract | Dysuria, hematuria, hemorrhagic cystitis, renal allograft dysfunction | Hematopoietic stem cell transplant and solid organ transplant recipients | Types 11, 34, 35, 3, 7, 21 [8] |
| Disseminated disease | Disseminated | Hematopoietic stem cell transplant recipients, severe combined immunodeficiency (SCID) and patients undergoing antiblastic treatment. | Species A, B, C, D (most common) and F [7,21] |
| Rare manifestations | Encephalitis, meningitis Myocarditis, cardiomyopathy Mononucleosis-like syndrome Bronchopulmonary dysplasia Intestinal intussusception Sudden infant death | Mainly described in children [22] | Types 2, 3, 7 neurologic manifestations |
| Marker | Advantages | Disadvantages | Limitations |
|---|---|---|---|
| CRP/procalcitonin | Early rise in serum in acute infections | May not differentiate HAdV and bacterial infections | Potential application in URTIs |
| White Blood Cells | Elevated in the acute HAdV infections | May not differentiate viral and bacterial infections | Observed only in some species [69] |
| Neutrophils | Elevated in HAdV rather than other viral infections | Typical also of bacterial infections | May not be suitable in older patients |
| Monocytes | Elevated in URTIs, prognostic significance | Not exclusive marker for HAdV | Prognostic value studied only in young adults |
| T CD4+ | Elevated in URTIs and silent HAdV infections | - | Observed in some types, Not routinely assessed |
| T reg | - | - | Few and conflicting data, Not routinely assessed |
| IL-10 | Early rise in serum in acute infections, prognostic significance | May not differentiate viral and bacterial infections [70] | Prognostic significance evaluated only in severe infections |
| IL-6 | Elevated in HAdV rather than other viral infections, prognostic significance, may be assessed in nasal swabs | May not differentiate HAdV and bacterial infections | - |
| IL-8 | Prognostic significance in children, may be assessed in nasal swabs, more stable along the disease course | Loss of prognostic value in young adults | - |
| IL-1β and TNF-α | - | Not exclusive marker for HAdV | Unclear course in HAdV infection |
| IFN-γ | Elevated in the acute HAdV infections | Not exclusive marker for HAdV in children | Prognostic value evaluated only in severe infections in young adults |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biserni, G.B.; Scarpini, S.; Dondi, A.; Biagi, C.; Pierantoni, L.; Masetti, R.; Sureshkumar, S.; Rocca, A.; Lanari, M. Potential Diagnostic and Prognostic Biomarkers for Adenovirus Respiratory Infection in Children and Young Adults. Viruses 2021, 13, 1885. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091885
Biserni GB, Scarpini S, Dondi A, Biagi C, Pierantoni L, Masetti R, Sureshkumar S, Rocca A, Lanari M. Potential Diagnostic and Prognostic Biomarkers for Adenovirus Respiratory Infection in Children and Young Adults. Viruses. 2021; 13(9):1885. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091885
Chicago/Turabian StyleBiserni, Giovanni Battista, Sara Scarpini, Arianna Dondi, Carlotta Biagi, Luca Pierantoni, Riccardo Masetti, Sugitha Sureshkumar, Alessandro Rocca, and Marcello Lanari. 2021. "Potential Diagnostic and Prognostic Biomarkers for Adenovirus Respiratory Infection in Children and Young Adults" Viruses 13, no. 9: 1885. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091885