
Identification and Characterization of a New Type of Holin-Endolysin Lysis Cassette in Acidovorax oryzae Phage AP1


 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Bioinformatics Analysis
2.3. Standard DNA Manipulation, PCR and DNA Sequencing
2.4. Recombinant Plasmids Construction
2.5. Growth Measurement
2.6. Protein Expression, Purification and Western-Blotting
2.7. Detection of β-Galactosidase Activity
2.8. Live/Dead Cell Staining
2.9. Microscopy Analysis
2.10. Bacterial Two-Hybrid Assays
3. Results
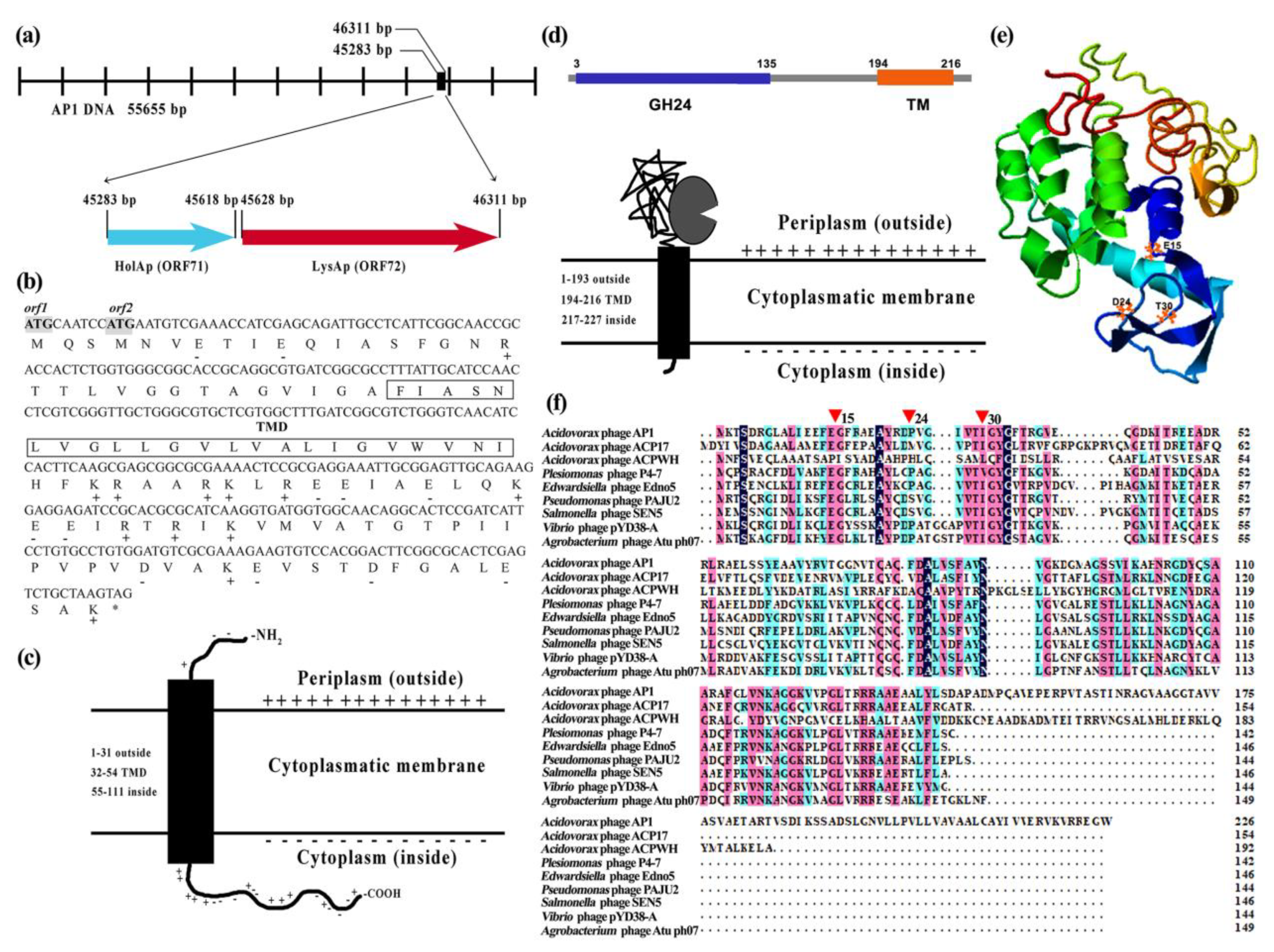
3.1. In Silico Description of AP1 Lysis Cassette
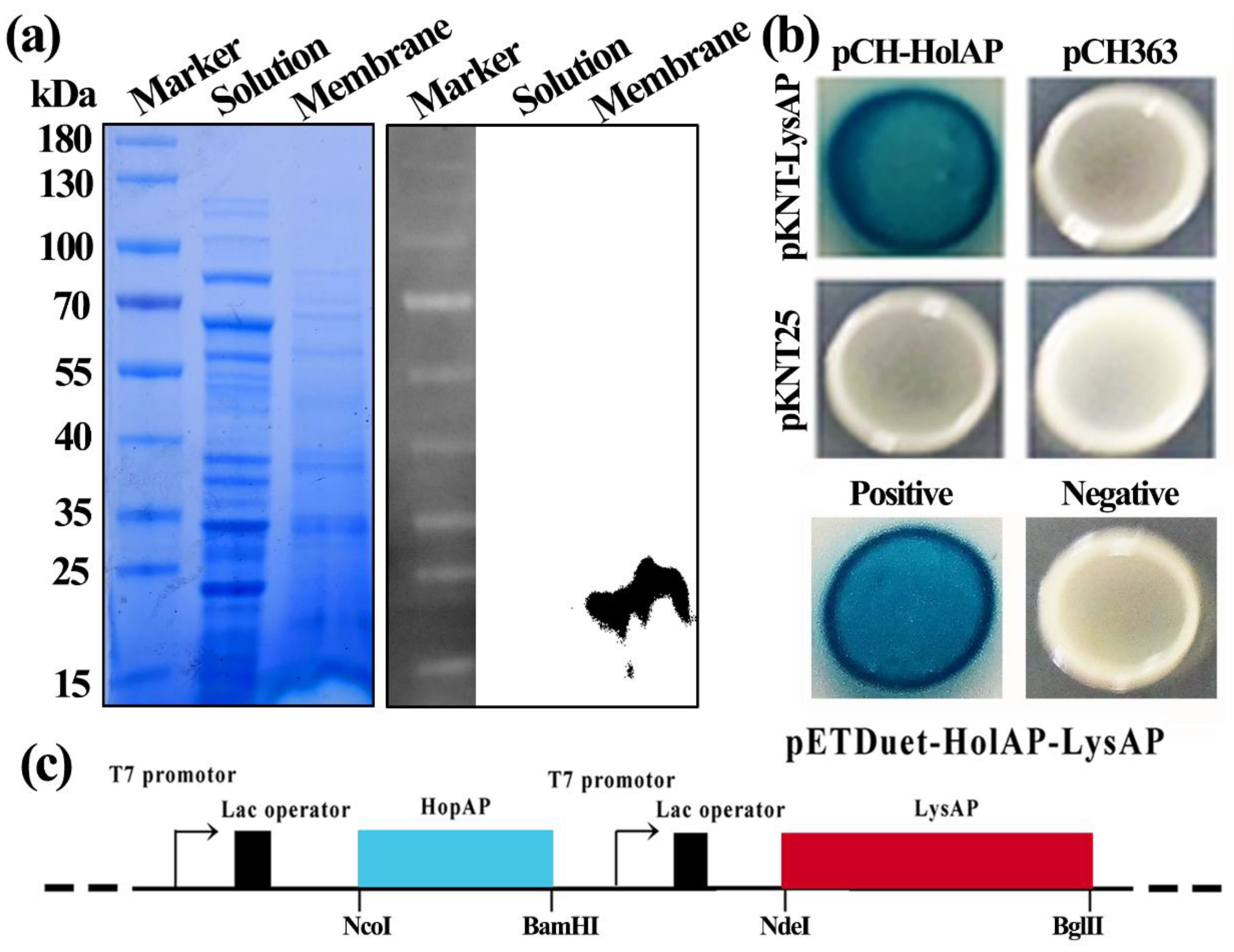
3.2. Holin HolAP Inserts in the Cell Membrane and Interacts with Endolysin LysAP
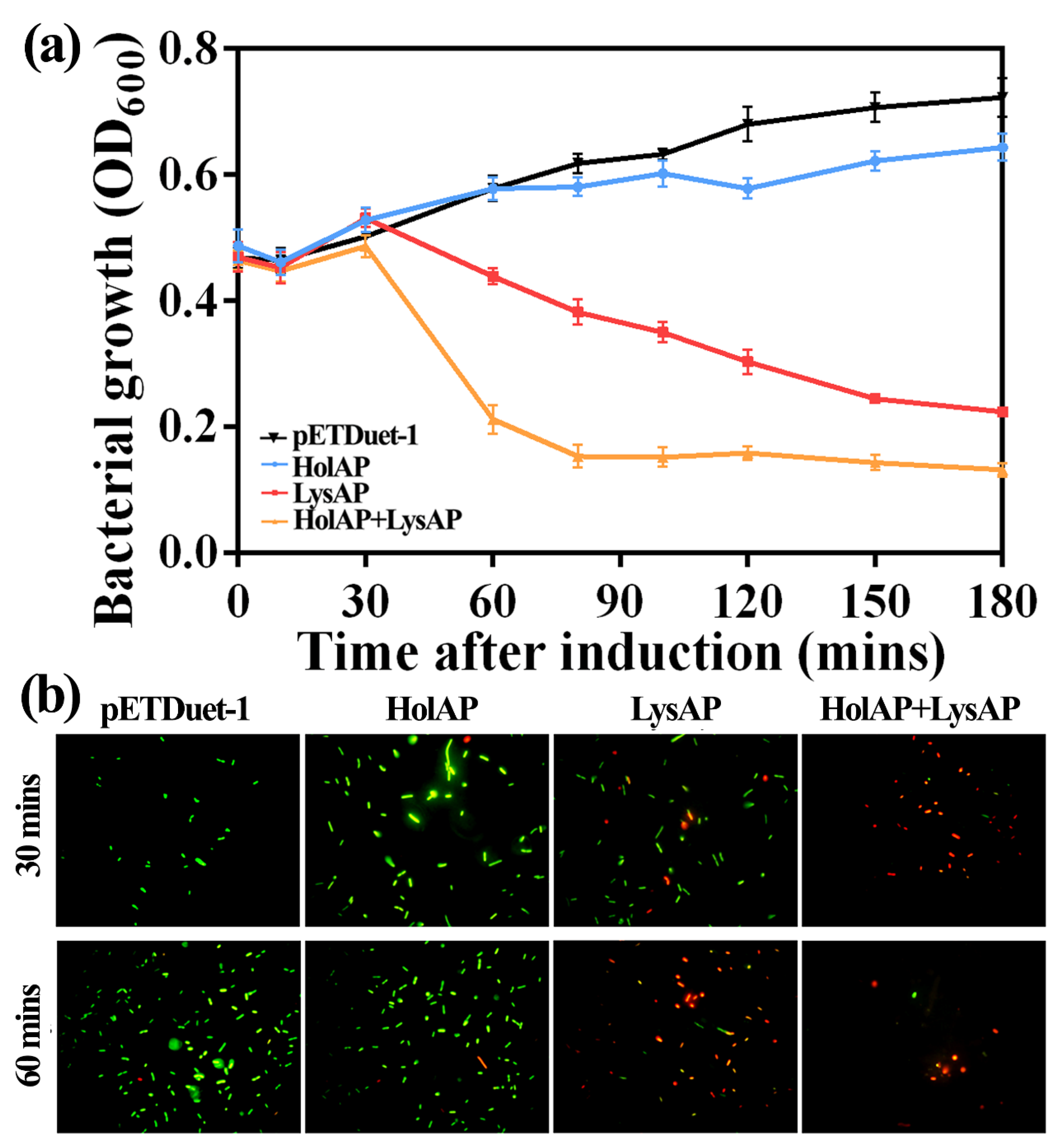
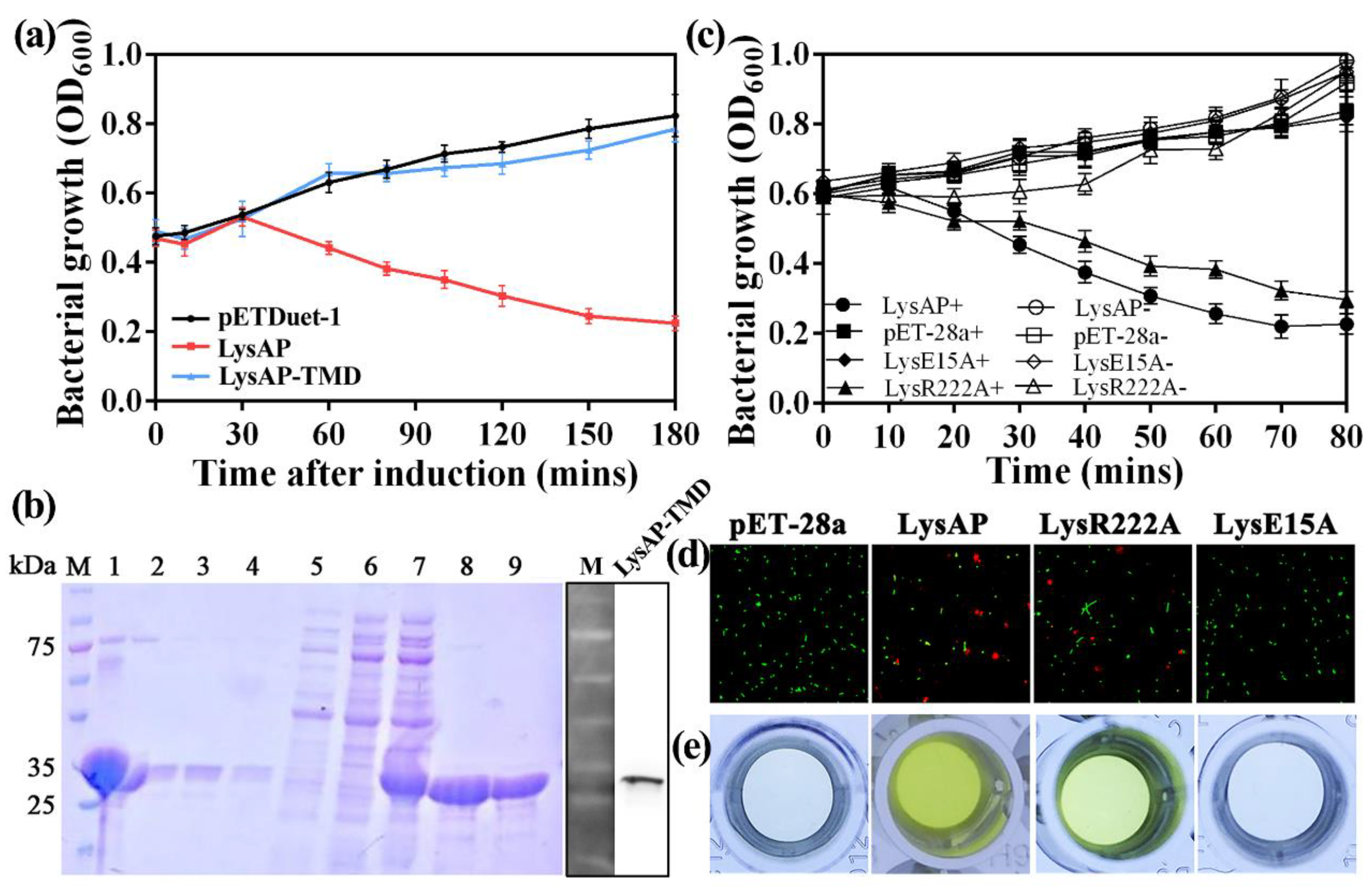
3.3. Expression of AP1 Lysis Cassette Leads to Cell Lysis
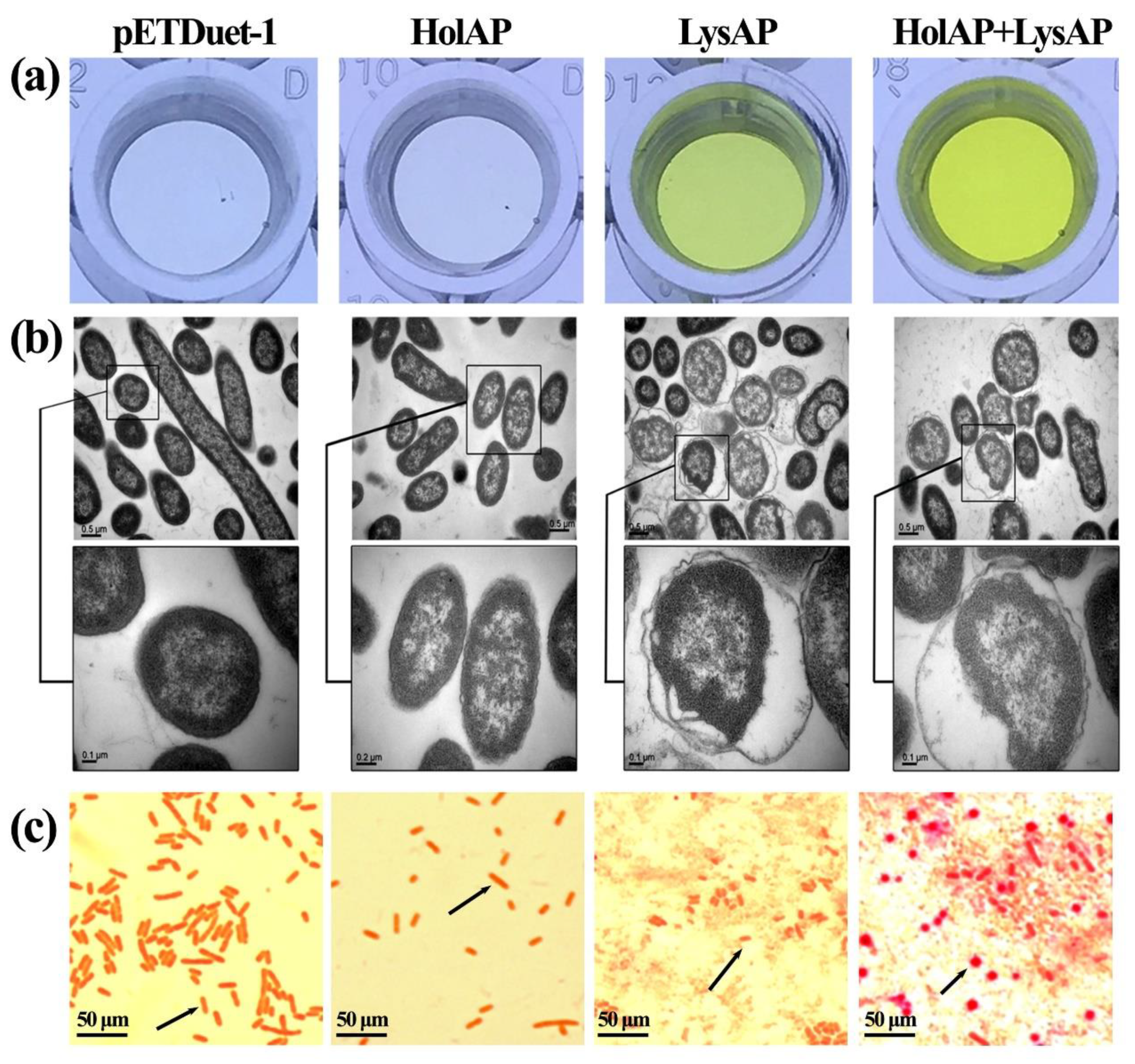
3.4. Expression of AP1 Lysis Cassette Proteins Affect Membrane Integrity and Induce Morphological Changes
3.5. The C-Terminal Transmembrane Domain (TMD) Plays an Important Role in LysAP Lysis
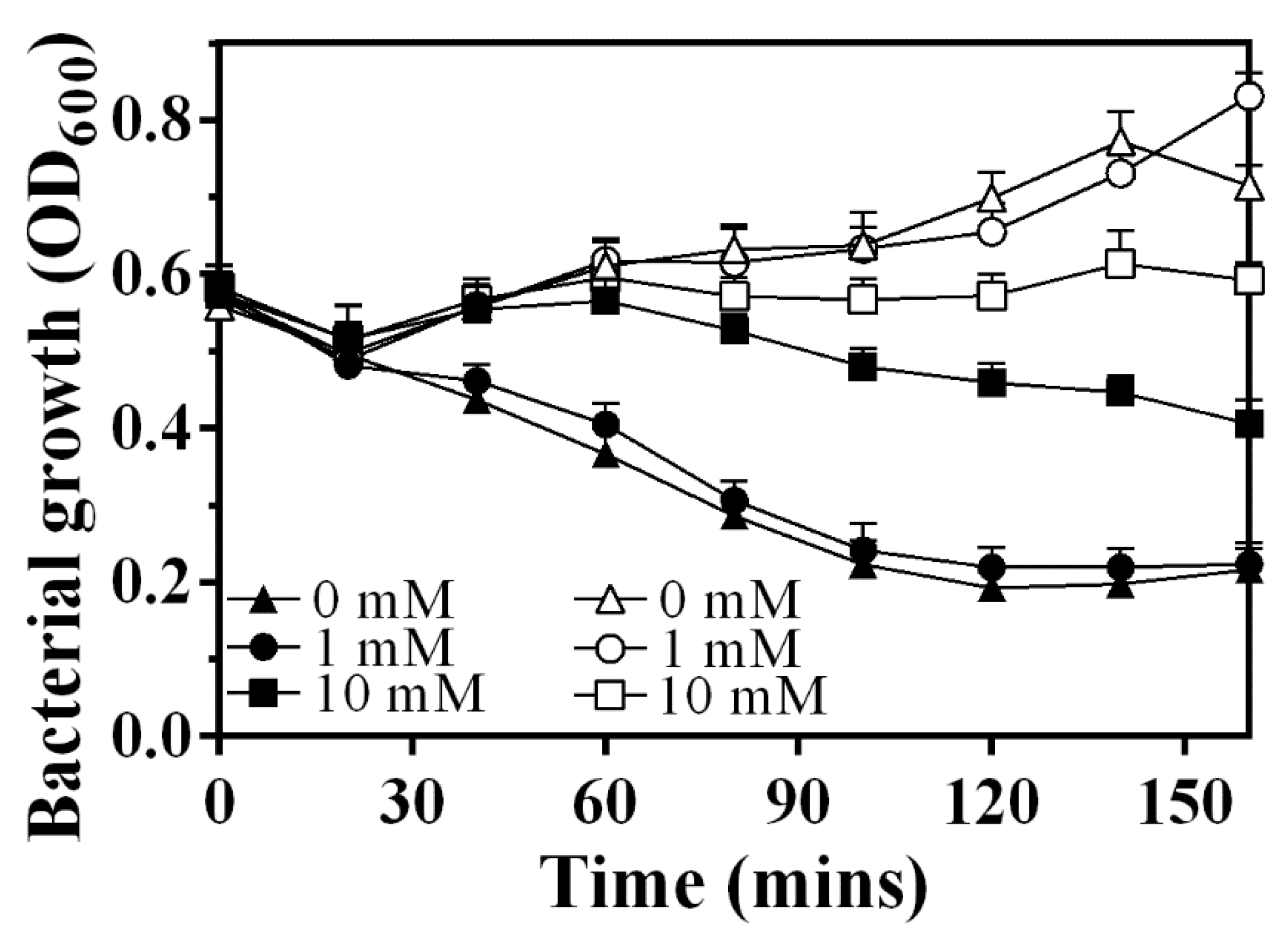
3.6. Sec System Is Involved in the Release Process of LysAP
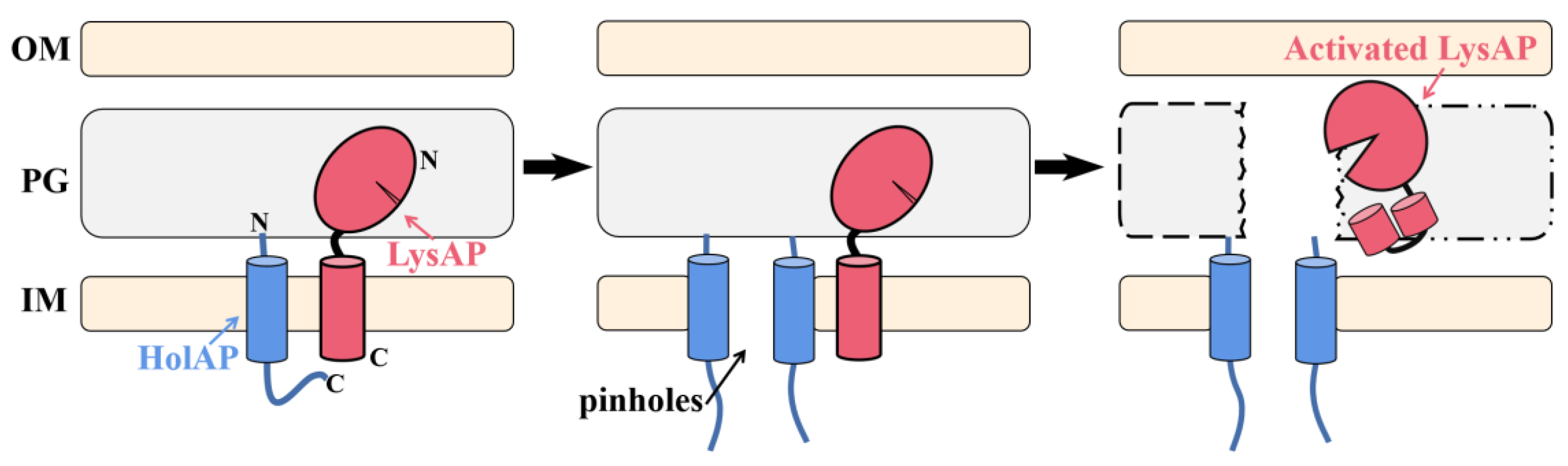
3.7. Lysis Model for C-Terminal SAR Endolysin-Holin Cassette
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bernhardt, T.G.; Wang, I.-N.; Struck, D.K.; Young, R. Breaking free: “Protein antibiotics” and phage lysis. Res. Microbiol. 2002, 153, 493–501. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, Y.; Xu, X.; Ahmed, T.; Yang, Y.; Loh, B.; Leptihn, S.; Yan, C.; Chen, J.; Li, B. The Holin-Endolysin Lysis System of the OP2-Like Phage X2 Infecting Xanthomonas oryzae pv. oryzae. Viruses 2021, 13, 1949. [Google Scholar] [CrossRef] [PubMed]
- Briers, Y.; Walmagh, M.; Van Puyenbroeck, V.; Cornelissen, A.; Cenens, W.; Aertsen, A.; Oliveira, H.; Azeredo, J.; Verween, G.; Pirnay, J.-P.; et al. Engineered Endolysin-Based “Artilysins” To Combat Multidrug-Resistant Gram-Negative Pathogens. mBio 2014, 5, e01379-14. [Google Scholar] [CrossRef] [Green Version]
- Lai, W.C.B.; Chen, X.; Ho, M.K.Y.; Xia, J.; Leung, S.S.Y. Bacteriophage-derived endolysins to target gram-negative bacteria. Int. J. Pharm. 2020, 589, 119833. [Google Scholar] [CrossRef]
- Nie, T.; Meng, F.; Lu, F.; Sun, J.; Bie, X.; Lu, Z.; Lu, Y. Molecular dynamics insight of novel Enzybiotic Salmcide-p1 lysis peptidoglycan to inhibit Salmonella Typhimurium. Food Control 2022, 132, 108564. [Google Scholar] [CrossRef]
- Ahammad, T.; Khan, R.H.; Sahu, I.D.; Drew, D.L.; Faul, E.; Li, T.; McCarrick, R.M.; Lorigan, G.A. Pinholin S21 mutations induce structural topology and conformational changes. Biochim. Biophys. Acta (BBA)—Biomembr. 2021, 1863, 183771. [Google Scholar] [CrossRef]
- Xu, H.; Bao, X.; Hong, W.; Wang, A.; Wang, K.; Dong, H.; Hou, J.; Govinden, R.; Deng, B.; Chenia, H.Y. Biological Characterization and Evolution of Bacteriophage T7-△holin During the Serial Passage Process. Front. Microbiol. 2021, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Young, R. Phage lysis: Do we have the hole story yet? Curr. Opin. Microbiol. 2013, 16, 790–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil Nascimento, J.; Guerreiro-Pereira, M.C.; Costa, S.F.; São-José, C.; Santos, M.A. Nisin-Triggered Activity of Lys44, the Secreted Endolysin from Oenococcus oeni Phage fOg44. J. Bacteriol. 2008, 190, 457–461. [Google Scholar] [CrossRef] [Green Version]
- Catalao, M.J.; Gil, F.; Moniz-Pereira, J.; São-José, C.; Pimentel, M. Diversity in bacterial lysis systems: Bacteriophages show the way. FEMS Microbiol. Rev. 2013, 37, 554–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, B.; Qi, X.; Wang, X.; Ren, H.; Liu, W.; Zhang, C. Characterization of a Novel Bacteriophage swi2 Harboring Two Lysins Can Naturally Lyse Escherichia coli. Front. Microbiol. 2021, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Arulandu, A.; Struck, D.K.; Swanson, S.; Sacchettini, J.C.; Young, R. Disulfide Isomerization After Membrane Release of Its SAR Domain Activates P1 Lysozyme. Science 2005, 307, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Struck, D.K.; Deaton, J.; Wang, I.-N.; Young, R. A signal-arrest-release sequence mediates export and control of the phage P1 endolysin. Proc. Natl. Acad. Sci. USA 2004, 101, 6415–6420. [Google Scholar] [CrossRef] [Green Version]
- Ogunyemi, S.O.; Fang, Y.; Qiu, W.; Li, B.; Chen, J.; Yang, M.; Hong, X.; Luo, J.; Wang, Y.; Sun, G. Role of type IV secretion system genes in virulence of rice bacterial brown stripe pathogen Acidovorax oryzae strain RS-2. Microb. Pathog. 2019, 126, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chai, Y.; Guo, J.-H.; Losick, R. Evidence for Cyclic Di-GMP-Mediated Signaling in Bacillus subtilis. J. Bacteriol. 2012, 194, 5080–5090. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W.; Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 2001; pp. 11803–12500. [Google Scholar]
- Zhang, M.; Wang, X.; Ahmed, T.; Liu, M.; Wu, Z.; Luo, J.; Tian, Y.; Jiang, H.; Wang, Y.; Sun, G.; et al. Identification of Genes Involved in Antifungal Activity of Burkholderia seminalis against Rhizoctonia solani Using Tn5 Transposon Mutation Method. Pathogens 2020, 9, 797. [Google Scholar] [CrossRef]
- Briers, Y.; Peeters, L.M.; Volckaert, G.; Lavigne, R. The lysis cassette of bacteriophage фKMV encodes a signal-arrest-release endolysin and a pinholin. Bacteriophage 2011, 1, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Springer, K.; Reuter, S.; Knüpfer, M.; Schmauder, L.; Sänger, P.-A.; Felsl, A.; Fuchs, T.M. Activity of a Holin-Endolysin System in the Insecticidal Pathogenicity Island of Yersinia enterocolitica. J. Bacteriol. 2018, 200, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunyemi, S.O.; Abdallah, Y.; Zhang, M.; Fouad, H.; Hong, X.; Ibrahim, E.; Masum, M.I.; Hossain, A.; Mo, J.; Li, B. Green synthesis of zinc oxide nanoparticles using different plant extracts and their antibacterial activity against Xanthomonas oryzae pv. oryzae. Artif. Cells Nanomed. Biotechnol. 2019, 47, 341–352. [Google Scholar] [CrossRef] [Green Version]
- Karimova, G.; Pidoux, J.; Ullmann, A.; Ladant, D. A bacterial two-hybrid system based on a reconstituted signal transduction pathway. Proc. Natl. Acad. Sci. USA 1998, 95, 5752–5756. [Google Scholar] [CrossRef] [Green Version]
- Gontijo, M.T.P.; Jorge, G.P.; Brocchi, M. Current Status of Endolysin-Based Treatments against Gram-Negative Bacteria. Antibiotics 2021, 10, 1143. [Google Scholar] [CrossRef]
- Oliveira, H.; Boas, D.V.; Mesnage, S.; Kluskens, L.D.; Lavigne, R.; Sillankorva, S.; Secundo, F.; Azeredo, J. Structural and Enzymatic Characterization of ABgp46, a Novel Phage Endolysin with Broad Anti-Gram-Negative Bacterial Activity. Front. Microbiol. 2016, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- Khan, F.M.; Gondil, V.S.; Li, C.; Jiang, M.; Li, J.; Yu, J.; Wei, H.; Yang, H. A Novel Acinetobacter baumannii Bacteriophage Endolysin LysAB54 With High Antibacterial Activity Against Multiple Gram-Negative Microbes. Front. Cell. Infect. Microbiol. 2021, 11, 11. [Google Scholar] [CrossRef]
- Chen, N.; Hong, F.-L.; Wang, H.-H.; Yuan, Q.-H.; Ma, W.-Y.; Gao, X.-N.; Shi, R.; Zhang, R.-J.; Sun, C.-S.; Wang, S.-B. Modified Recombinant Proteins Can Be Exported via the Sec Pathway in Escherichia coli. PLoS ONE 2012, 7, e42519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, H.; Yang, H.; Li, J.; Xiao, S.; Hu, S.; Yan, F.; Xia, L.; Zhang, Y. Screening of a Plesiomonas shigelloides phage and study of the activity of its lysis system. Virus Res. 2021, 306, 198581. [Google Scholar] [CrossRef]
- Li, X.; Zhang, C.; Wei, F.; Yu, F.; Zhao, Z. Bactericidal activity of a holin-endolysin system derived from Vibrio alginolyticus phage HH109. Microb. Pathog. 2021, 159, 105135. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.; Rajaure, M.; Pang, T.; Young, R. The Spanin Complex Is Essential for Lambda Lysis. J. Bacteriol. 2012, 194, 5667–5674. [Google Scholar] [CrossRef] [Green Version]
- Holt, A.; Cahill, J.; Ramsey, J.; Martin, C.; O’Leary, C.; Moreland, R.; Maddox, L.T.; Galbadage, T.; Sharan, R.; Sule, P.; et al. Phage-encoded cationic antimicrobial peptide required for lysis. J. Bacteriol. 2021. [Google Scholar] [CrossRef]
- Cahill, J.; Young, R. Phage Lysis: Multiple Genes for Multiple Barriers. In Advances in Virus Research; Kielian, M., Mettenleiter, T.C., Roossinck, M.J., Eds.; Academic Press Ltd-Elsevier Science Ltd: London, UK, 2019; Volume 103, pp. 33–70. [Google Scholar]
- Park, T.; Struck, D.K.; Deaton, J.F.; Young, R. Topological dynamics of holins in programmed bacterial lysis. Proc. Natl. Acad. Sci. USA 2006, 103, 19713–19718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, I.-N. Lysis Timing and Bacteriophage Fitness. Genetics 2006, 172, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Krupovič, M.; Bamford, D.H. Holin of bacteriophage lambda: Structural insights into a membrane lesion. Mol. Microbiol. 2008, 69, 781–783. [Google Scholar] [CrossRef]
- Bavda, V.R.; Jain, V. Deciphering the Role of Holin in Mycobacteriophage D29 Physiology. Front. Microbiol. 2020, 11, 883. [Google Scholar] [CrossRef]
- Ramankulov, Y.; Young, R. Genetic analysis of the T4 holin: Timing and topology. Gene 2001, 265, 25–36. [Google Scholar] [CrossRef]
- Niu, X.; Guiltinan, M.J. DNA binding specificity of the wheat bZIP protein EmBP-1. Nucleic Acids Res. 1994, 22, 4969–4978. [Google Scholar] [CrossRef] [Green Version]
- To, K.H.; Young, R. Probing the Structure of the S105 Hole. J. Bacteriol. 2014, 196, 3683–3689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamczyk-Popławska, M.; Tracz-Gaszewska, Z.; Lasota, P.; Kwiatek, A.; Piekarowicz, A. Haemophilus influenzae HP1 Bacteriophage Encodes a Lytic Cassette with a Pinholin and a Signal-Arrest-Release Endolysin. Int. J. Mol. Sci. 2020, 21, 4013. [Google Scholar] [CrossRef]
- Pang, T.; Savva, C.G.; Fleming, K.G.; Struck, D.K.; Young, R. Structure of the lethal phage pinhole. Proc. Natl. Acad. Sci. USA 2009, 106, 18966–18971. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Source or Reference | |
|---|---|---|
| Strains | ||
| RS-2 | Acidovorax oryzae, pathogen of bacterial brown stripe | Lab collection |
| DH5α | F-Φ80d lacZΔM15Δ(lacZYA-argF) U169 recA1 endA1, hsdR17(rk-, mk+) phoAsupE44 λ- thi-1 gyrA96 relA, Escherichia coli | Vazyme |
| BL21(DE3) | Host for overexpressing proteins driven by T7 promoter, Escherichia coli | Invitrogen |
| BTH101 | Host for overexpressing proteins in bacterial two-hybrid, Escherichia coli | Chen et al. [15] |
| Phage | ||
| AP1 | phage of Acidovorax oryzae | Lab collection |
| Plasmids | ||
| pETDuet-1 | AmpR; expression vector with HIS label | Promega |
| pETDuet-HolAP | AmpR; recombinant expression vector with HIS label with HolAP | This study |
| pETDuet-LysAP | AmpR; recombinant expression vector with HIS label with LysAP | This study |
| pETDuet-HolAP-LysAP | AmpR; recombinant expression vector with HIS label with HolAP and LysAP | This study |
| pETDuet-LysAPΔTMD | AmpR; recombinant expression vector with HIS label with LysAPΔTMD | This study |
| pET-28a(+) | KanR; expression vector with HIS label | Novagen |
| pET-28a-HolAP | KanR; recombinant expression vector with HIS label with HolAP | This study |
| pET-28a-LysAP | KanR; recombinant expression vector with HIS label with LysAP | This study |
| pET-28a-LysAPΔTMD | KanR; recombinant expression vector with HIS label with LysAPΔTMD | This study |
| pET-28a-LysR222A | KanR; point mutation recombinant expression vector with HIS label with LysAP | This study |
| pET-28a-LysE15A | KanR; point mutation recombinant expression vector with HIS label with LysAP | This study |
| pKNT25 | KanR; expression vector for bacterial two-hybrid test | Chen et al. [15] |
| pKNT-LysAP | KanR; recombinant expression vector for B2H test | This study |
| pCH363 | AmpR; expression vector for bacterial two-hybrid test | Chen et al. [15] |
| pCH-HolAP | AmpR; recombinant expression vector for B2H test | This study |
| Primers Name | Sequences (5′-3′) | Length |
|---|---|---|
| pETDuet-LysF | CTATACATATGATGAAAACCTCTGATCGCGGAC | 684 bp |
| pETDuet-LysR | GAAGATCTTGACCACCCCTCTCGCCG | |
| pET28a-LysF | CGGGATCCATGAAAACCTCTGATCGCGGAC | 684 bp |
| pET28a-LysR | CCCAAGCTTTGACCACCCCTCTCGCCG | |
| Lys△TMDF | CGGGATCCATGAAAACCTCTGATCGCGGACTCGC | 615 bp |
| Lys△TMDR | CCCAAGCTTTGACCACCCCTCTCGCCGCACCTTCACACGCTCGTCAGCGCTCGACTTGATG | |
| E15A-F | AGCAAATGGGTCGCGGATCCATGAAAACCTCTGATCGCGGACTCGCGCTGATCGAAGAATTCGCGGGCTTC | 684 bp |
| E15A-R | TCGAGTGCGGCCGCAAGCTTTCATGACCACCCCTCTCGCC | |
| R222A-F | AGCAAATGGGTCGCGGATCCATGAAAACCTCTGATCGCGGACTCGC | 684 bp |
| R222A-R | TCGAGTGCGGCCGCAAGCTTTCATGACCACCCCTCTGCCCGCACCTTCACACG | |
| pETDuet-HolF | TATGCCATGGATGCAATCCATGAATGTCGAAAC | 336 bp |
| pETDuet-HolR | CGGGATCCCTTAGCAGACTCGAGTGCG | |
| pET28a-HolF | CGGGATCCATGCAATCCATGAATGTCGAAACC | 336 bp |
| pET28a-HolR | CCCAAGCTTCTTAGCAGACTCGAGTGCG | |
| pCH-HolF | GCAAGCTTATGCAATCCATGAATGTCGAAACC | 336 bp |
| pCH-HolR | CGGAATTCCTTAGCAGACTCGAGTGCG | |
| pKNT-LysF | CGGGATCCATGAAAACCTCTGATCGCGGAC | 684 bp |
| pKNT-LysR | TATAGAATTCTGACCACCCCTCTCGCCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Wang, Y.; Chen, J.; Hong, X.; Xu, X.; Wu, Z.; Ahmed, T.; Loh, B.; Leptihn, S.; Hassan, S.; et al. Identification and Characterization of a New Type of Holin-Endolysin Lysis Cassette in Acidovorax oryzae Phage AP1. Viruses 2022, 14, 167. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020167
Zhang M, Wang Y, Chen J, Hong X, Xu X, Wu Z, Ahmed T, Loh B, Leptihn S, Hassan S, et al. Identification and Characterization of a New Type of Holin-Endolysin Lysis Cassette in Acidovorax oryzae Phage AP1. Viruses. 2022; 14(2):167. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020167
Chicago/Turabian StyleZhang, Muchen, Yanli Wang, Jie Chen, Xianxian Hong, Xinyan Xu, Zhifeng Wu, Temoor Ahmed, Belinda Loh, Sebastian Leptihn, Sabry Hassan, and et al. 2022. "Identification and Characterization of a New Type of Holin-Endolysin Lysis Cassette in Acidovorax oryzae Phage AP1" Viruses 14, no. 2: 167. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020167