
A Bacteriophage Cocktail Significantly Reduces Listeria monocytogenes without Deleterious Impact on the Commensal Gut Microbiota under Simulated Gastrointestinal Conditions

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteriophage Cocktail
2.2. Bacterial Strains
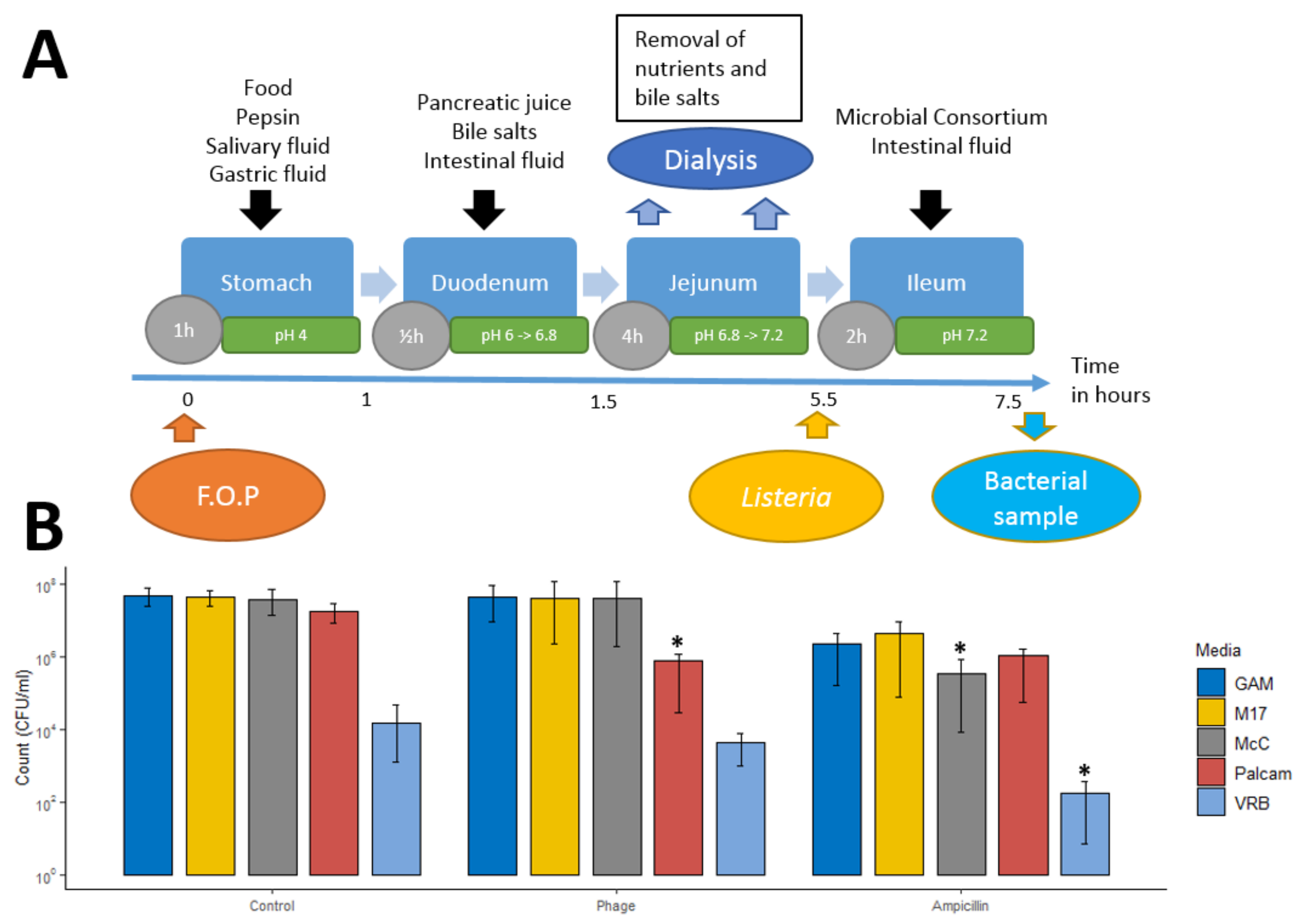
2.3. Small Intestinal Model System
2.3.1. Small Intestine In Vitro Simulation
2.3.2. Consortium of Small Intestinal Bacteria
2.4. Bacteriophage Impact on L. monocytogenes during Stomach and Small Intestine Passage
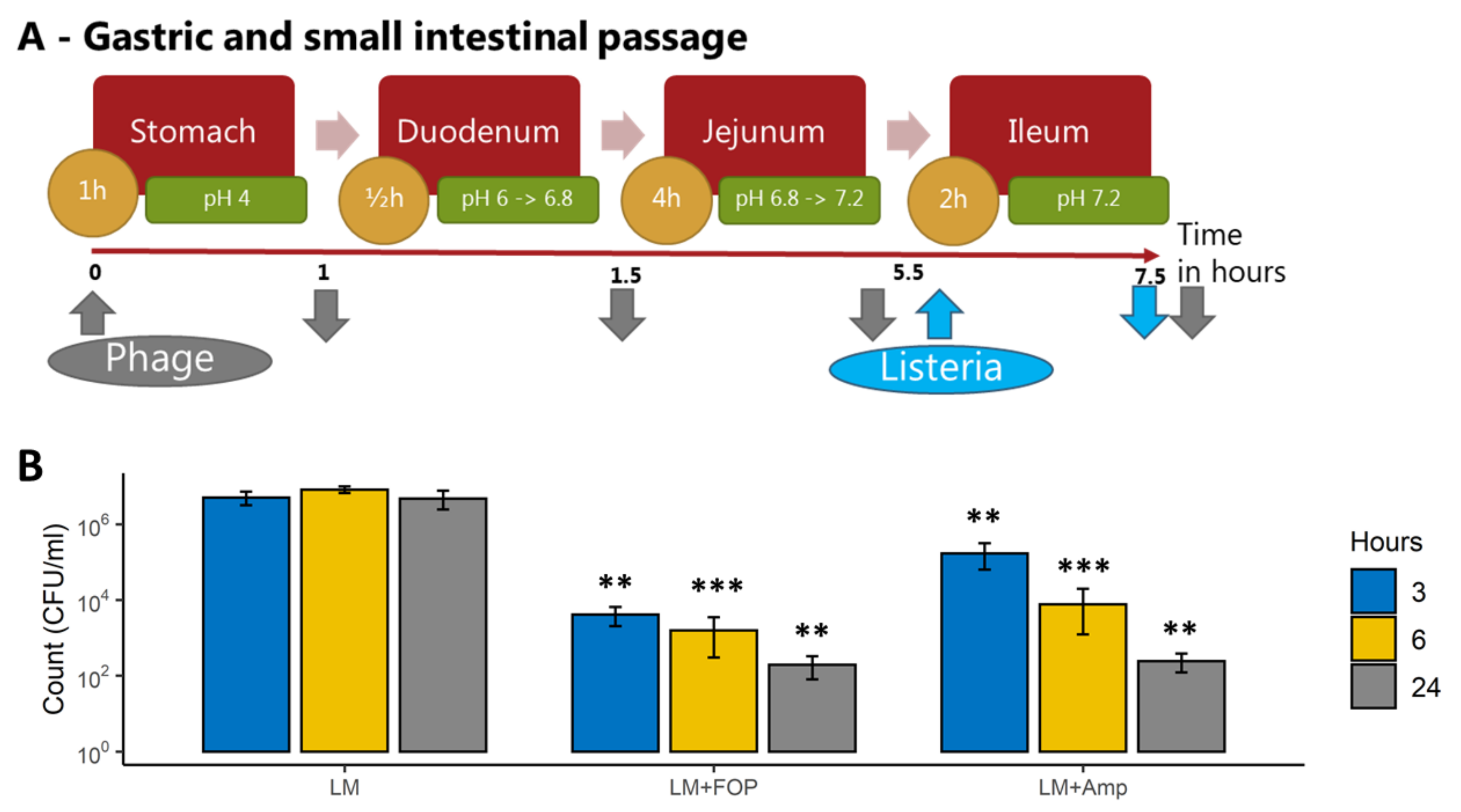
2.5. Colon Model System
2.5.1. Large Intestine Model System
2.5.2. Impact of the FOP Bacteriophage Preparation on L. monocytogenes and Colon Microbiome
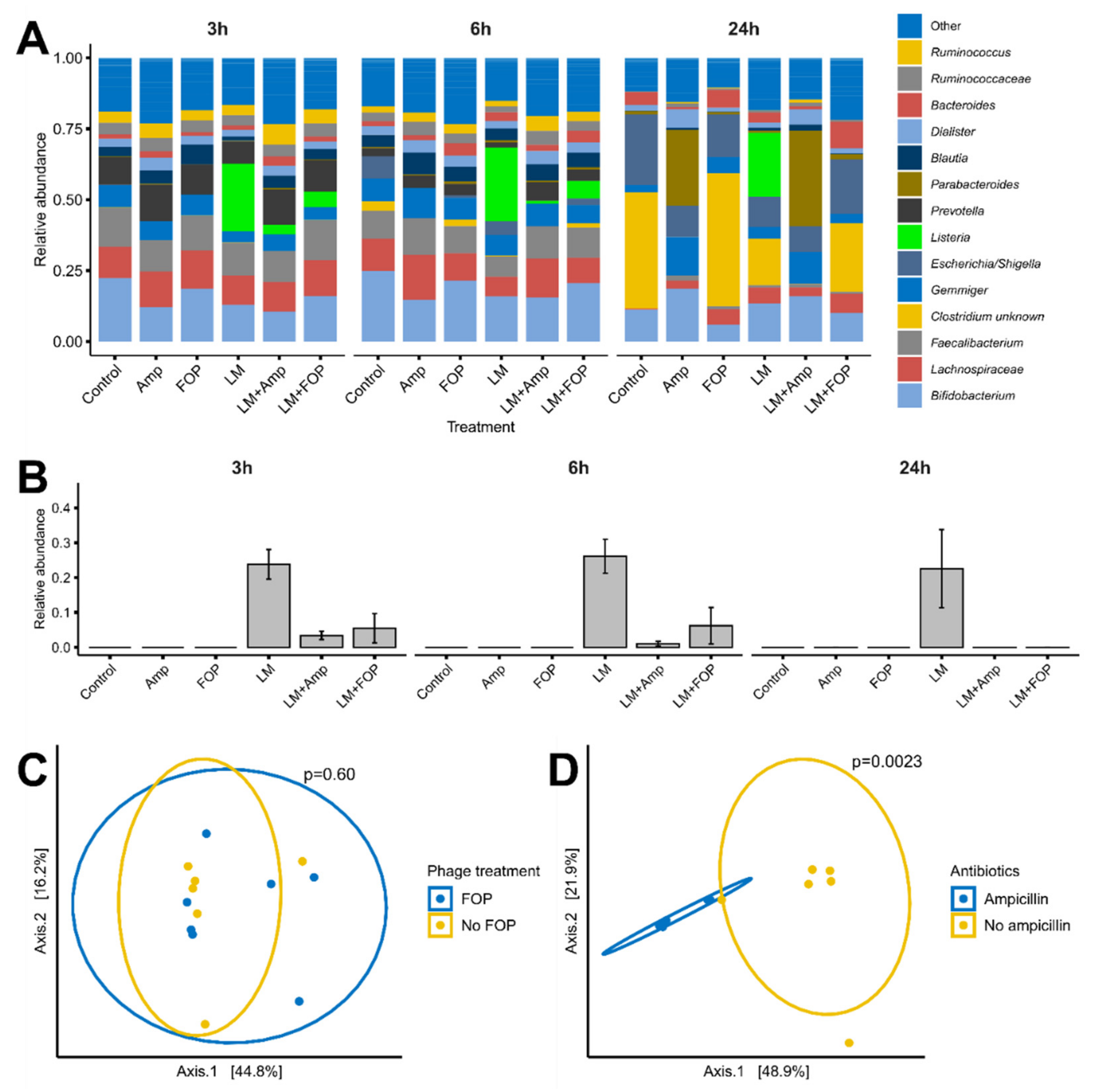
2.6. Sequencing of Bacterial Community
2.6.1. Library Preparation and Sequencing
2.6.2. Bioinformatic Analysis
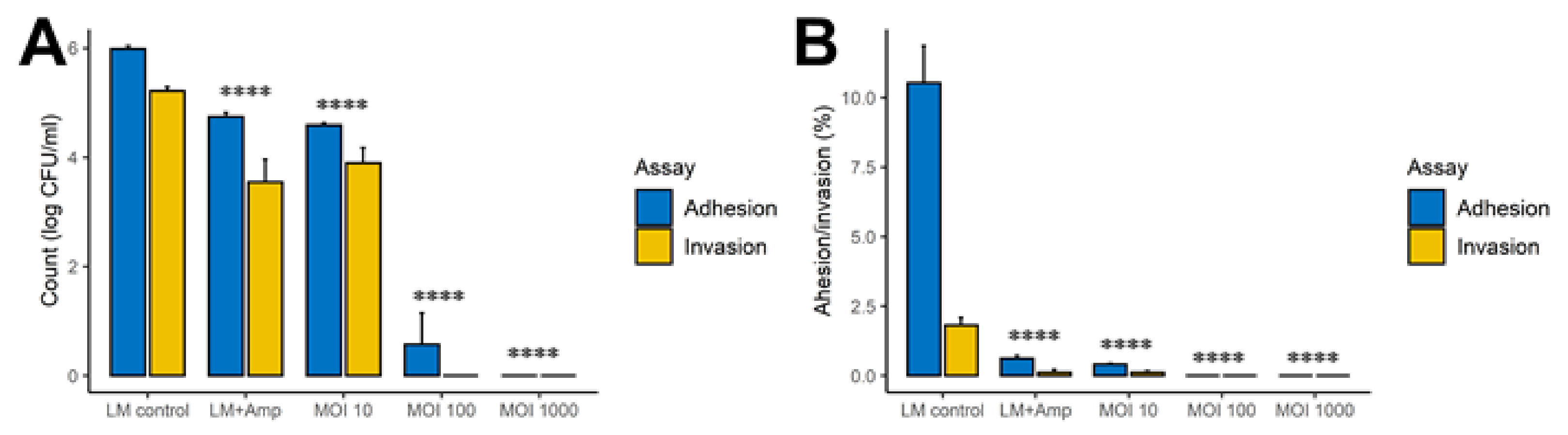
2.7. Caco-2 Intestinal Epithelial Model
2.7.1. Caco-2 Cell Culturing
2.7.2. Adhesion and Invasion Assay
2.7.3. Transepithelial Resistance (TER) Assay
2.7.4. Statistics
3. Results
3.1. The FOP Bacteriophage Cocktail Selectively Reduces Listeria monocytogenes in a Small Intestine In Vitro Model
3.2. The FOP Bacteriophage Cocktail Significantly Reduces L. monocytogenes in a Colon Model While Preserving Bacterial Community Structure
3.3. The FOP Bacteriophage Cocktail Significantly Reduces L. monocytogenes Adhesion and Invasion of Caco-2 Cells
3.4. Cytokine Production and Transepithelial Resistance in Caco-2 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- ECDC. Listeriosis, Annual Epidemiological Report for 2017; Surveill Rep 6; ECDC: Stockholm, Sweden, 2020.
- De Noordhout, C.M.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.; Kirk, M.; Havelaar, A.; Speybroeck, N. The global burden of listeriosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef] [Green Version]
- Rácz, P.; Tenner, K.; Mérö, E. Experimental Listeria enteritis. I. An electron microscopic study of the epithelial phase in experimental listeria infection. Lab. Investig. 1972, 26, 694–700. [Google Scholar] [PubMed]
- Cieplak, T.; Soffer, N.; Sulakvelidze, A.; Nielsen, D.S. A bacteriophage cocktail targeting Escherichia coli reduces E. coli in simulated gut conditions, while preserving a non-targeted representative commensal normal microbiota. Gut Microbes 2018, 9, 391–399. [Google Scholar]
- Domingo-Calap, P.; Delgado-Martínez, J. Bacteriophages: Protagonists of a post-antibiotic era. Antibiotics 2018, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Segall, A.M.; Roach, D.R.; Strathdee, S.A. Stronger together? Perspectives on phage-antibiotic synergy in clinical applications of phage therapy. Curr. Opin. Microbiol. 2019, 51, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Mahony, J.; McAuliffe, O.; Ross, R.P.; van Sinderen, D. Bacteriophages as biocontrol agents of food pathogens. Curr. Opin. Biotechnol. 2011, 22, 157–163. [Google Scholar] [CrossRef]
- United States Food and Drug Administration Center for Food Safety and Applied Nutrition. Agency Response Letter, GRAS Notice No. 000198. 2007. Available online: http://www.cfsan.fda.gov/~rdb/opa-g198.html (accessed on 5 December 2021).
- Endersen, L.; Coffey, A. The use of bacteriophages for food safety. Curr. Opin. Food Sci. 2020, 36, 1–8. [Google Scholar] [CrossRef]
- Hernandez-Milian, A.; Payeras-Cifre, A. What is new in Listeriosis? BioMed Res. Int. 2014, 2014, 358051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage Therapy: A Renewed Approach to Combat Antibiotic-Resistant Bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dąbrowska, K. Phage therapy: What factors shape phage pharmacokinetics and bioavailability? Systematic and critical review. Med. Res. Rev. 2019, 39, 2000–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moye, Z.D.; Woolston, J.; Van den Abbeele, P.; Duysburgh, C.; Verstrepen, L.; Marzorati, M.; Sulakvelidze, A. A Bacteriophage Cocktail Eliminates Salmonella Typhimurium from the Human Colonic Microbiome while Preserving Cytokine Signaling and Preventing Attachment to and Invasion of Human Cells by Salmonella In Vitro. J. Food Prot. 2019, 82, 1336–1349. [Google Scholar] [CrossRef]
- Lamy-Besnier, Q.; Chaffringeon, L.; Lourenço, M.; Payne, R.B.; Trinh, J.T.; Schwartz, J.A.; Sulakvelidze, A.; Debarbieux, L. Prophylactic Administration of a Bacteriophage Cocktail Is Safe and Effective in Reducing Salmonella enterica Serovar Typhimurium Burden in Vivo. Microbiol. Spectr. 2021, 9, e00497-21. [Google Scholar] [CrossRef]
- Dulbecco, R.; Vogt, M. Some problems of animal virology as studied by the plaque technique. Cold Spring Harb. Symp. Quant. Biol. 1953, 18, 273–279. [Google Scholar] [CrossRef]
- Cieplak, T.; Wiese, M.; Nielsen, S.; Van de Wiele, T.; van den Berg, F.; Nielsen, D.S. The Smallest Intestine (TSI)-a low volume in vitro model of the small intestine with increased throughput. FEMS Microbiol. Lett. 2018, 365, fny231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dlugosz, A.; Winckler, B.; Lundin, E.; Zakikhany, K.; Sandström, G.; Ye, W.; Engstrand, L.; Lindberg, G. No difference in small bowel microbiota between patients with irritable bowel syndrome and healthy controls. Sci. Rep. 2015, 5, 8508. [Google Scholar] [CrossRef]
- Chung, C.S.; Chang, P.F.; Liao, C.H.; Lee, T.H.; Chen, Y.; Lee, Y.C.; Wu, M.S.; Wang, H.P.; Ni, Y.H. Differences of microbiota in small bowel and faeces between irritable bowel syndrome patients and healthy subjects. Scand. J. Gastroenterol. 2016, 51, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Van Netten, P.; van Gaal, B.; Mossel, D.A.A. Selection, differentiation and counting of haemolytic Listeria spp. on PALCAM medium. Lett. Appl. Microbiol. 1991, 12, 20–22. [Google Scholar] [CrossRef]
- Wiese, M.; Khakimov, B.; Nielsen, S.; Sørensen, H.; van den Berg, F.; Nielsen, D.S. CoMiniGut—A small volume in vitro colon model for the screening of gut microbial fermentation processes. PeerJ 2018, 6, e4268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krych, Ł.; Kot, W.; Bendtsen, K.M.B.; Hansen, A.K.; Vogensen, F.K.; Nielsen, D.S. Have you tried spermine? A rapid and cost-effective method to eliminate dextran sodium sulfate inhibition of PCR and RT-PCR. J. Microbiol. Methods 2018, 144, 1–7. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. bioRxiv 2016, 081257. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R. SINTAX: A simple non-Bayesian taxonomy classifier for 16S and ITS sequences. bioRxiv 2016, 074161. [Google Scholar]
- Kim, O.S.; Cho, Y.J.; Lee, K.; Yoon, S.H.; Kim, M.; Na, H.; Park, S.C.; Jeon, Y.S.; Lee, J.H.; Yi, H.; et al. Introducing EzTaxon-e: A prokaryotic 16s rRNA gene sequence database with phylotypes that represent uncultured species. Int. J. Syst. Evol. Microbiol. 2012, 62, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Team RC. R: A Language and Environment for Statistical Title. 4.0.1; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Paulson, J.N.; Stine, O.C.; Bravo, H.C.; Pop, M. Differential abundance analysis for microbial marker-gene surveys. Nat. Methods 2013, 10, 1200–1202. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.; Xu, Z.Z.; Peddada, S.; Amir, A.; Bittinger, K.; Gonzalez, A.; Lozupone, C.; Zaneveld, J.R.; Vázquez-Baeza, Y.; Birmingham, A.; et al. Normalization and microbial differential abundance strategies depend upon data characteristics. Microbiome 2017, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Paulson, J. metagenomeSeq: Statistical analysis for sparse high-throughput sequencing. Bioconductor Package 2014, 1, 191. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Kassambara, A. “ggplot2” Based Publication Ready Plots [R Package Ggpubr VERSION 0.4.0]. Available online: https://rpkgs.datanovia.com/ggpubr/ (accessed on 5 December 2021).
- Wickham, H. ggplot2. Wiley Interdiscip. Rev. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Klingberg, T.D.; Pedersen, M.H.; Cencic, A.; Budde, B.B. Application of measurements of transepithelial electrical resistance of intestinal epithelial cell monolayers to evaluate probiotic activity. Appl. Environ. Microbiol. 2005, 71, 7528–7530. [Google Scholar] [CrossRef] [Green Version]
- Quereda, J.J.; Dussurget, O.; Nahori, M.A.; Ghozlane, A.; Volant, S.; Dillies, M.A.; Regnault, B.; Kennedy, S.; Mondot, S.; Villoing, B.; et al. Bacteriocin from epidemic Listeria strains alters the host intestinal microbiota to favor infection. Proc. Natl. Acad. Sci. USA 2016, 113, 5706–5711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becattini, S.; Pamer, E.G. Multifaceted defense against listeria monocytogenes in the gastro-intestinal lumen. Pathogens 2018, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, V.; Ukhanova, M.; Visone, L.; Abuladze, T.; Sulakvelidze, A. Bacteriophage administration reduces the concentration of Listeria monocytogenes in the gastrointestinal tract and its translocation to spleen and liver in experimentally infectedmice. Int. J. Microbiol. 2010, 2010, 624234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, T.S.; Koefoed, A.K.; Jakobsen, R.R.; Deng, L.; Castro-Mejía, J.L.; Brunse, A.; Neve, H.; Vogensen, F.K.; Nielsen, D.S. Bacteriophage-mediated manipulation of the gut microbiome—Promises and presents limitations. FEMS Microbiol. Rev. 2020, 44, 507–521. [Google Scholar] [CrossRef]
- García, R.; Latz, S.; Romero, J.; Higuera, G.; García, K.; Bastías, R. Bacteriophage production models: An overview. Front. Microbiol. 2019, 10, 01187. [Google Scholar] [CrossRef]
- Tsuchiya, K.; Kawamura, I.; Takahashi, A.; Nomura, T.; Kohda, C.; Mitsuyama, M. Listeriolysin O-induced membrane permeation mediates persistent interleukin-6 production in Caco-2 cells during Listeria monocytogenes infection in vitro. Infect. Immun. 2005, 73, 3869–3877. [Google Scholar] [CrossRef] [Green Version]
- Kutter, E.; Sulakvelidze, A. Bacteriophage therapy in humans. In Bacteriophages: Biology and Applications; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Fallingborg, J.; Christensen, L.A.; Ingeman-Nielsen, M.; Jacobsen, B.A.; Abildgaard, K.; Rasmussen, H.H.; Rasmussen, S.N. Measurement of gastrointestinal pH and regional transit times in normal children. J. Pediatr. Gastroenterol. Nutr. 1990, 11, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, R.; Matsumoto, T.; Sano, G.; Ishii, Y.; Tateda, K.; Sumiyama, Y.; Uchiyama, J.; Sakurai, S.; Matsuzaki, S.; Imai, S.; et al. Efficacy of bacteriophage therapy against gut-derived sepsis caused by Pseudomonas aeruginosa in mice. Antimicrob. Agents Chemother. 2007, 51, 446–452. [Google Scholar] [CrossRef] [Green Version]
- Ly-Chatain, M.H. The factors affecting effectiveness of treatment in phages therapy. Front. Microbiol. 2014, 5, 51. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, M.; Chaffringeon, L.; Lamy-Besnier, Q.; Pédron, T.; Campagne, P.; Eberl, C.; Bérard, M.; Stecher, B.; Debarbieux, L.; De Sordi, L. The Spatial Heterogeneity of the Gut Limits Predation and Fosters Coexistence of Bacteria and Bacteriophages. Cell Host Microbe 2020, 28, 390–401.e5. [Google Scholar] [CrossRef] [PubMed]
- Malik, D.J.; Sokolov, I.J.; Vinner, G.K.; Mancuso, F.; Cinquerrui, S.; Vladisavljevic, G.T.; Clokie, M.R.J.; Garton, N.J.; Stapley, A.G.F.; Kirpichnikova, A. Formulation, stabilisation and encapsulation of bacteriophage for phage therapy. Adv. Colloid Interface Sci. 2017, 249, 100–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dissanayake, U.; Ukhanova, M.; Moye, Z.D.; Sulakvelidze, A.; Mai, V. Bacteriophages Reduce Pathogenic Escherichia coli Counts in Mice Without Distorting Gut Microbiota. Front. Microbiol. 2019, 10, 01984. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain Number | Origin | Culture Time (h) | Culture Media |
|---|---|---|---|---|
| Escherichia coli | DSM 1058 | Human origin | 24 | GAM |
| Streptococcus salivarius | DSM 20560 | Blood | 6 | GAM |
| Streptococcus luteinensis | DSM 15350 | Human origin | 24 | GAM |
| Enterococcus faecalis | DSM 20478 | Human feces | 24 | GAM |
| Bacteroides fragilis | DSM 2151 | Appendix abscess | 24 | GAM |
| Veillonella parvula | DSM 2008 | Human intestine | 48 | GAM |
| Flavonifractor plautii | DSM 6740 | Human feces | 48 | GAM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakobsen, R.R.; Trinh, J.T.; Bomholtz, L.; Brok-Lauridsen, S.K.; Sulakvelidze, A.; Nielsen, D.S. A Bacteriophage Cocktail Significantly Reduces Listeria monocytogenes without Deleterious Impact on the Commensal Gut Microbiota under Simulated Gastrointestinal Conditions. Viruses 2022, 14, 190. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020190
Jakobsen RR, Trinh JT, Bomholtz L, Brok-Lauridsen SK, Sulakvelidze A, Nielsen DS. A Bacteriophage Cocktail Significantly Reduces Listeria monocytogenes without Deleterious Impact on the Commensal Gut Microbiota under Simulated Gastrointestinal Conditions. Viruses. 2022; 14(2):190. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020190
Chicago/Turabian StyleJakobsen, Rasmus Riemer, Jimmy T. Trinh, Louise Bomholtz, Signe Kristine Brok-Lauridsen, Alexander Sulakvelidze, and Dennis Sandris Nielsen. 2022. "A Bacteriophage Cocktail Significantly Reduces Listeria monocytogenes without Deleterious Impact on the Commensal Gut Microbiota under Simulated Gastrointestinal Conditions" Viruses 14, no. 2: 190. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020190