
CRISPR/Cas9-Mediated Disruption of the lef8 and lef9 to Inhibit Nucleopolyhedrovirus Replication in Silkworms


 and
and 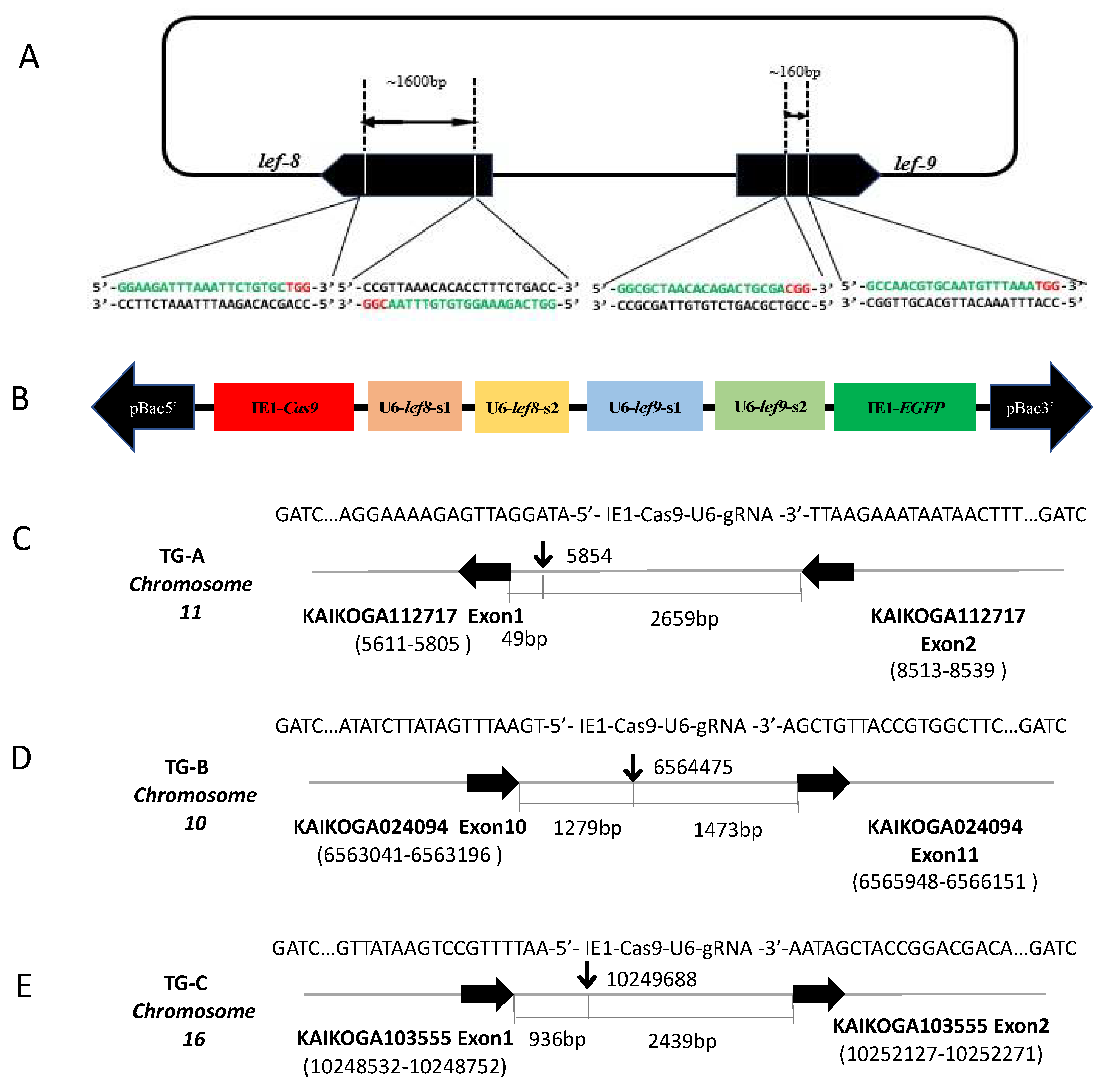{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Silkworm and Virus Inoculation
2.2. Vector Construction
2.3. Generating Transgenic Silkworms
2.4. Inverse Transcription-PCR and Sequencing of the Transgenic Silkworm
2.5. Viral Inoculation and Mortality Analyses
2.6. Mutagenesis Analysis of the Viral Target Genes
2.7. RNA Isolation and Quantitative Real-Time PCR (qPCR) of Viral Genes
2.8. Microscopy of Infected Midguts
2.9. Statistical Analysis
3. Results
3.1. Generation of Transgenic Silkworms
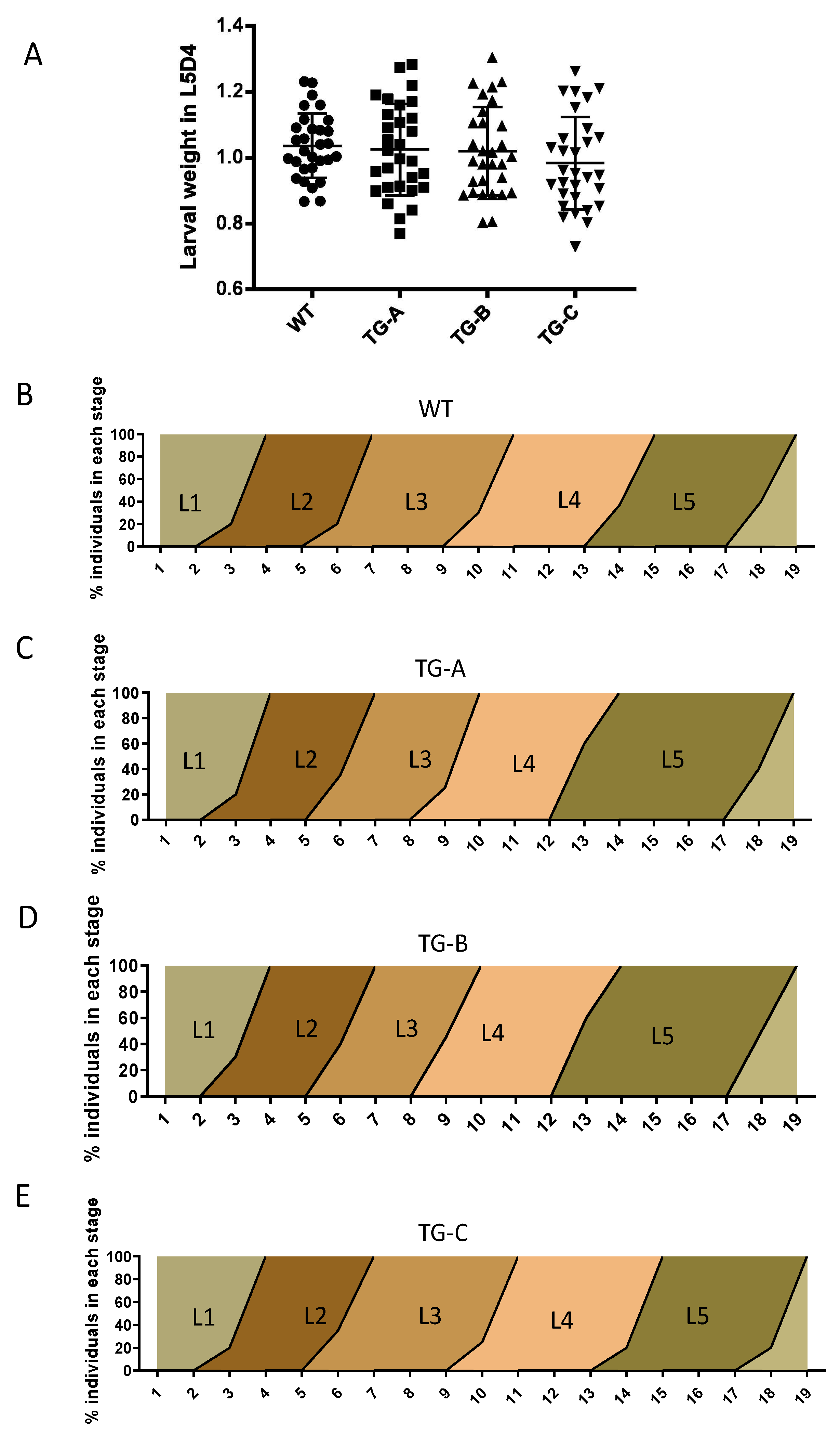
3.2. Normal Growth and Development in the Transgenic Lines
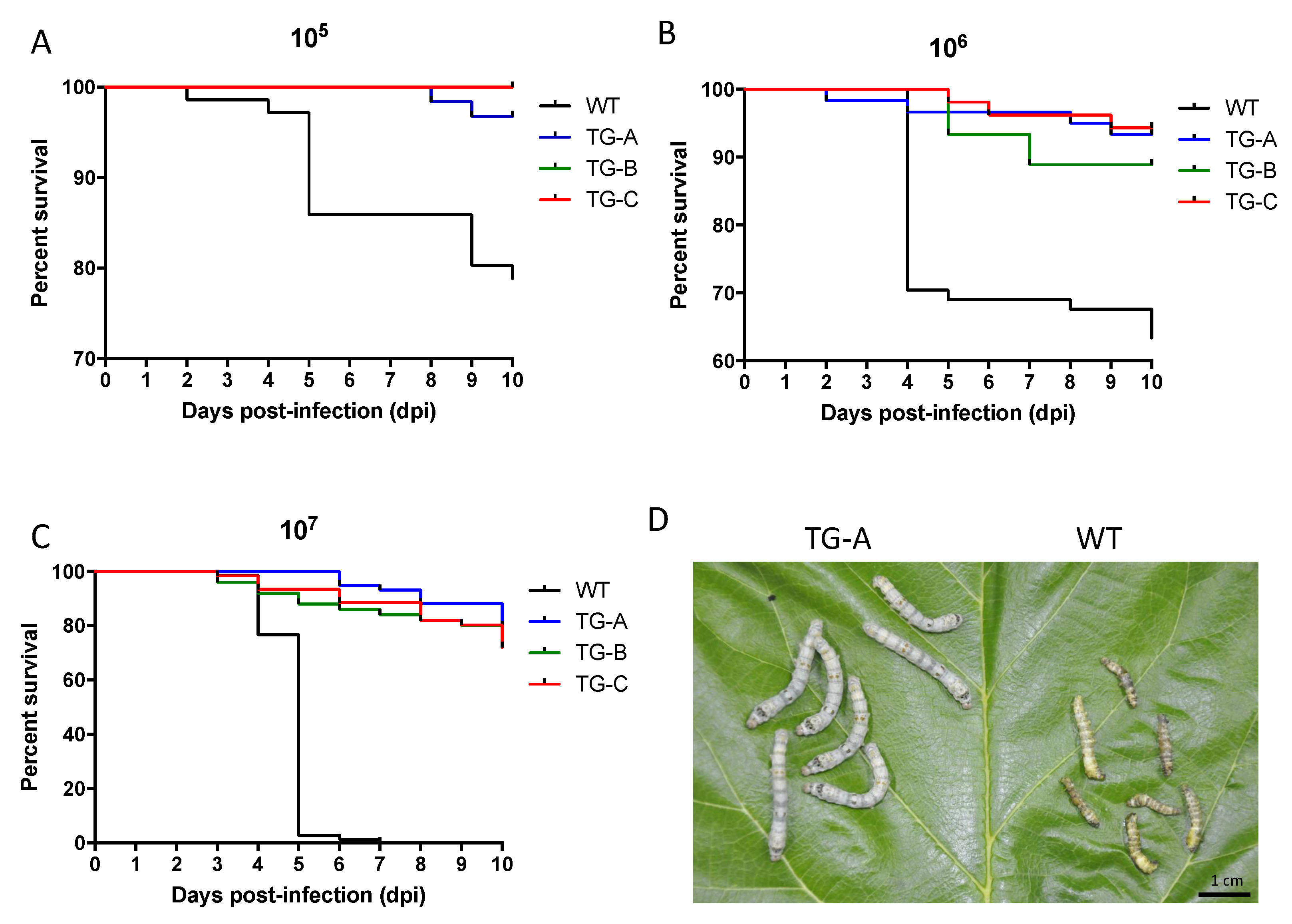
3.3. Transgenic Silkworms Showed Higher Survival Rate after BmNPV Infection
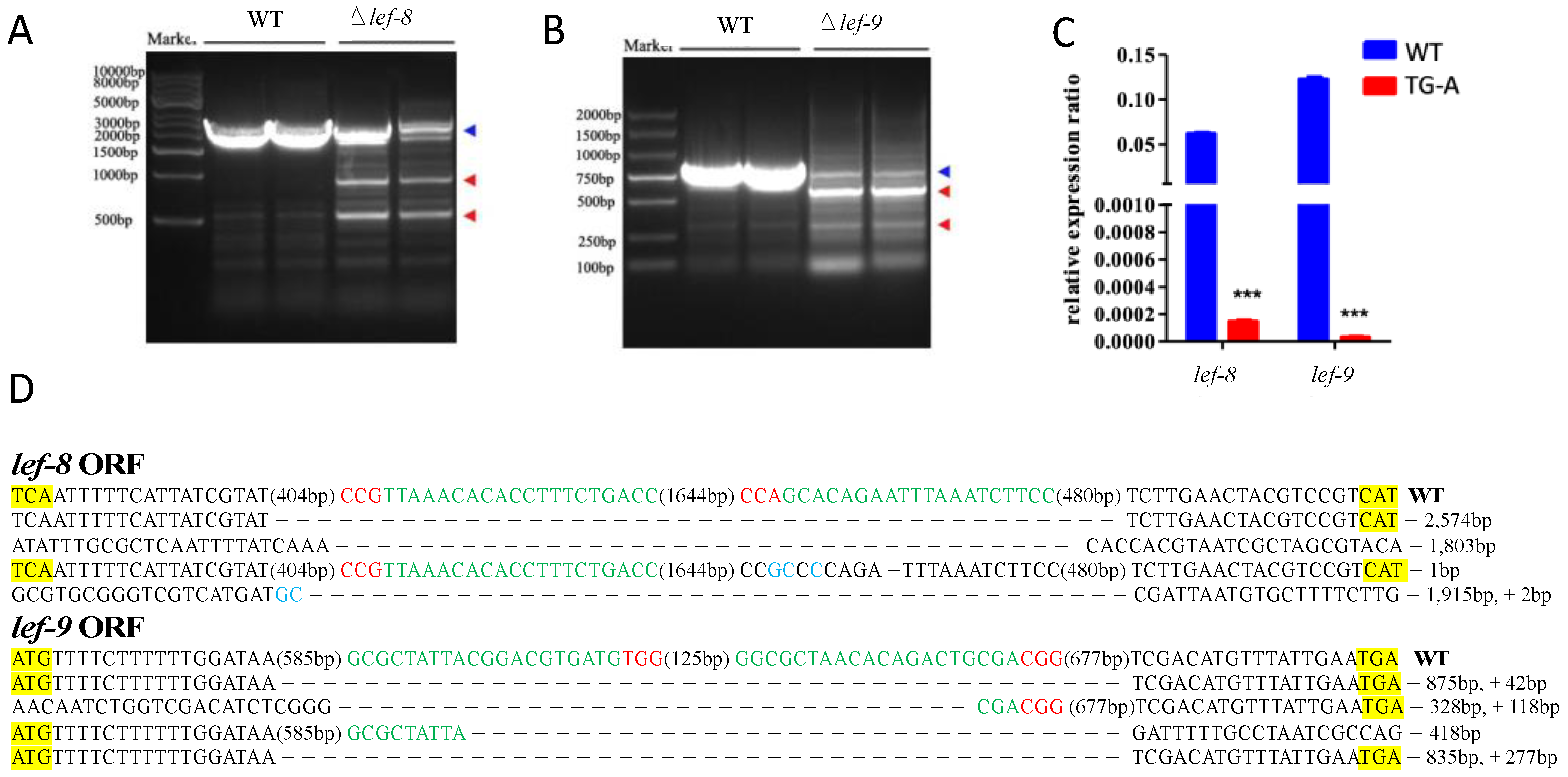
3.4. Targeted Mutations of BmNPV lef8 and lef9 in Infected Transgenic Silkworms
3.5. Microscopy Analysis of Midgut Cells in the Infected Transgenic and WT Silkworms
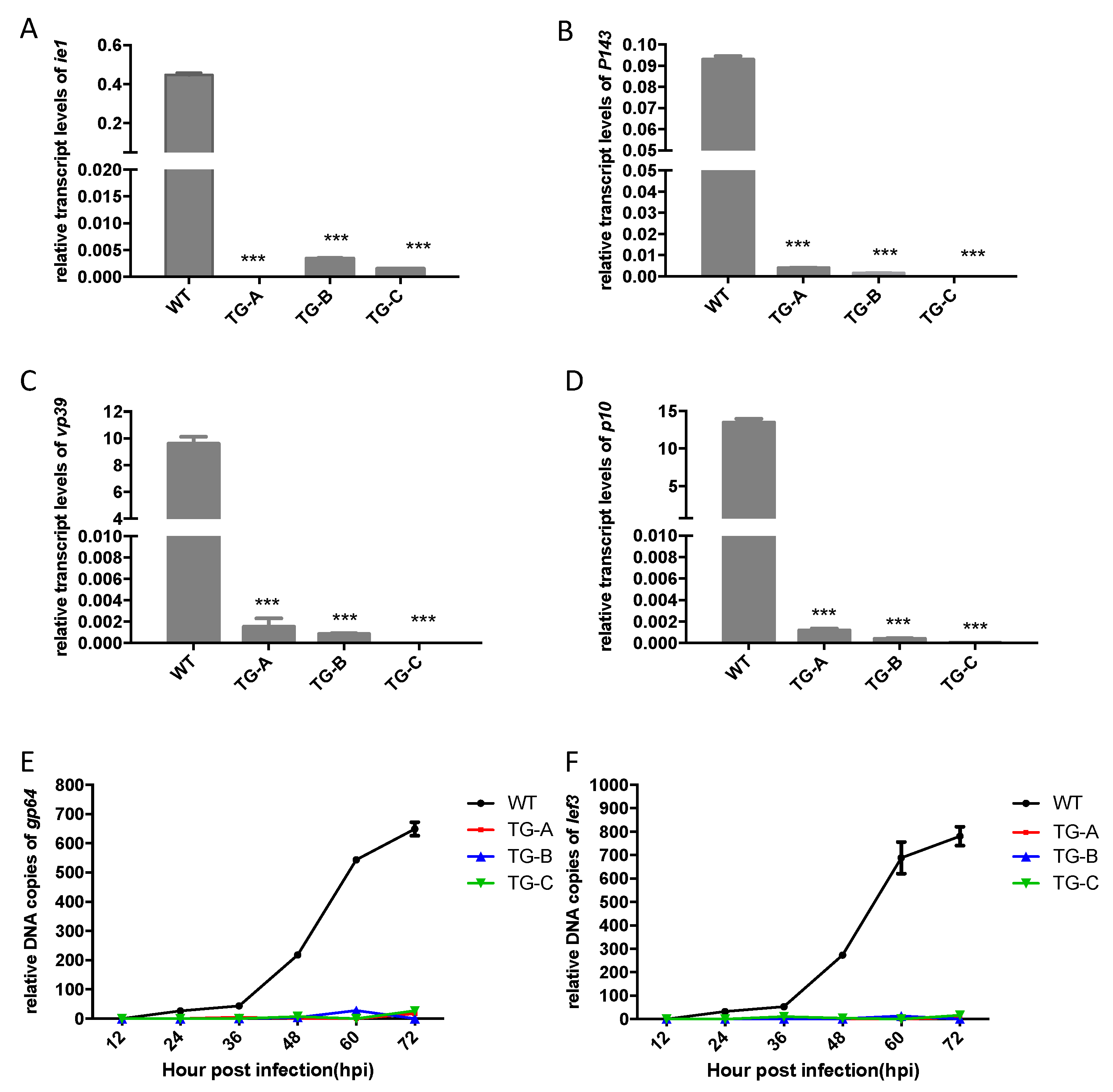
3.6. BmNPV Replication was Inhibited in Transgenic Silkworms
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, L.; Xia, Q. The progress and future of enhancing antiviral capacity by transgenic technology in the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2014, 48, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldsmith, M.R.; Shimada, T.; Abe, H. The genetics and genomics of the silkworm, Bombyx mori. Annu. Rev. Entomol. 2005, 50, 71–100. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Gopinathan, K.P. Systemic and in vitro infection process of Bombyx mori nucleopolyhedrovirus. Virus Res. 2004, 101, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Gomi, S.; Majima, K.; Maeda, S. Sequence analysis of the genome of Bombyx mori nucleopolyhedrovirus. J. Gen. Virol. 1999, 80, 1323–1337. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.L.; Xu, Y.P.; Zhang, C.X. Genome sequence of a Bombyx mori nucleopolyhedrovirus strain with cubic occlusion bodies. J. Virol. 2012, 86, 10245. [Google Scholar] [CrossRef] [Green Version]
- Rohrmann, G.F. Baculovirus Molecular Biology, 3rd ed.; National Center for Biotechnology Information: Bethesda, MD, USA, 2013.
- Lu, A.; Miller, L.K. The roles of eighteen baculovirus late expression factor genes in transcription and DNA replication. J. Virol. 1995, 69, 975–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapp, J.C.; Wilson, J.A.; Miller, L.K. Nineteen baculovirus open reading frames, including LEF-12, support late gene expression. J. Virol. 1998, 72, 10197–10206. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.D.; Carstens, E.B. Phenotypic characterization and physical mapping of a temperature-sensitive mutant of Autographa californica nuclear polyhedrosis virus defective in DNA synthesis. Virology 1984, 138, 69–81. [Google Scholar] [CrossRef]
- Guarino, L.A.; Xu, B.; Jin, J.; Dong, W. A virus-encoded RNA polymerase purified from baculovirus-infected cells. J. Virol. 1998, 72, 7985–7991. [Google Scholar] [CrossRef] [Green Version]
- Acharya, A.; Gopinathan, K.P. Characterization of late gene expression factors lef-9 and lef-8 from Bombyx mori nucleopolyhedrovirus. J. Gen. Virol. 2002, 83, 2015–2023. [Google Scholar] [CrossRef] [Green Version]
- Lu, A.; Miller, L.K. Identification of three late expression factor genes within the 33.8- to 43.4-map-unit region of Autographa californica nuclear polyhedrosis virus. J. Virol. 1994, 68, 6710–6718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passarelli, A.L.; Todd, J.W.; Miller, L.K. A baculovirus gene involved in late gene expression predicts a large polypeptide with a conserved motif of RNA polymerases. J. Virol. 1994, 68, 4673–4678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, C.; Kamagata, T.; Taka, H.; Sahara, K.; Asano, S.; Bando, H. Phenotypic grouping of 141 BmNPVs lacking viral gene sequences. Virus Res. 2012, 165, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ioannidis, K.; Swevers, L.; Iatrou, K. Bombyx mori nucleopolyhedrovirus lef-8 gene: Effects of deletion and implications for gene transduction applications. J. Gen. Virol. 2016, 97, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Crouch, E.A.; Cox, L.T.; Morales, K.G.; Passarelli, A.L. Inter-subunit interactions of the Autographa californica M nucleopolyhedrovirus RNA polymerase. Virology 2007, 367, 265–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isobe, R.; Kojima, K.; Matsuyama, T.; Quan, G.X.; Kanda, T.; Tamura, T.; Sahara, K.; Asano, S.I.; Bando, H. Use of RNAi technology to confer enhanced resistance to BmNPV on transgenic silkworms. Arch. Virol. 2004, 149, 1931–1940. [Google Scholar]
- Kanginakudru, S.; Royer, C.; Edupalli, S.V.; Jalabert, A.; Mauchamp, B.; Chandrashekaraiah; Prasad, S.V.; Chavancy, G.; Couble, P.; Nagaraju, J. Targeting ie-1 gene by RNAi induces baculoviral resistance in lepidopteran cell lines and in transgenic silkworms. Insect Mol. Biol. 2007, 16, 635–644. [Google Scholar] [CrossRef]
- Nishiyama, S.; Shitara, H.; Nakada, K.; Ono, T.; Sato, A.; Suzuki, H.; Ogawa, T.; Masaki, H.; Hayashi, J.; Yonekawa, H. Over-expression of Tfam improves the mitochondrial disease phenotypes in a mouse model system. Biochem. Biophys. Res. Commun. 2010, 401, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Wang, G.; Cheng, T.; Yang, Q.; Jin, S.; Lu, G.; Wu, F.; Xiao, Y.; Xu, H.; Xia, Q. Resistance to Bombyx mori nucleopolyhedrovirus via overexpression of an endogenous antiviral gene in transgenic silkworms. Arch. Virol. 2012, 157, 1323–1328. [Google Scholar] [CrossRef]
- de Bilbao, F.; Arsenijevic, D.; Moll, T.; Garcia-Gabay, I.; Vallet, P.; Langhans, W.; Giannakopoulos, P. In vivo over-expression of interleukin-10 increases resistance to focal brain ischemia in mice. J. Neurochem. 2009, 110, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Hou, C.; Bi, H.; Wang, Y.; Xu, J.; Li, M.; James, A.A.; Huang, Y.; Tan, A. Transgenic Clustered Regularly Interspaced Short Palindromic Repeat/Cas9-Mediated Viral Gene Targeting for Antiviral Therapy of Bombyx mori Nucleopolyhedrovirus. J. Virol. 2017, 91, e02465-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Chen, D.; Zhang, X.; Chen, S.; Yang, D.; Tang, L.; Yang, X.; Wang, Y.; Luo, X.; Wang, M.; et al. Construction of Baculovirus-Inducible CRISPR/Cas9 Antiviral System Targeting BmNPV in Bombyx mori. Viruses 2021, 14, 59. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.; Tanaka, H.; Tamura, T.; Shiotsuki, T. Precocious metamorphosis in transgenic silkworms overexpressing juvenile hormone esterase. Proc. Natl. Acad. Sci. USA 2005, 102, 11751–11756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.; Bhattacharya, A.; Debnath, N.; Datta, A.; Goswami, A. Nanoparticle-induced morphological transition of Bombyx mori nucleopolyhedrovirus: A novel method to treat silkworm grasserie disease. Appl. Microbiol. Biotechnol. 2013, 97, 6019–6030. [Google Scholar] [CrossRef]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Tsai, S.Q.; Sander, J.D.; Peterson, R.T.; Yeh, J.R.; Joung, J.K. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 227–229. [Google Scholar] [CrossRef]
- Tan, A.; Fu, G.; Jin, L.; Guo, Q.; Li, Z.; Niu, B.; Meng, Z.; Morrison, N.I.; Alphey, L.; Huang, Y. Transgene-based, female-specific lethality system for genetic sexing of the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2013, 110, 6766–6770. [Google Scholar] [CrossRef] [Green Version]
- Teng, X.; Zhang, Z.; He, G.; Yang, L.; Li, F. Validation of reference genes for quantitative expression analysis by real-time rt-PCR in four lepidopteran insects. J. Insect Sci. 2012, 12, 60. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Katsuma, S.; Kobayashi, J.; Koyano, Y.; Matsuda-Imai, N.; Kang, W.; Shimada, T. Baculovirus-encoded protein BV/ODV-E26 determines tissue tropism and virulence in lepidopteran insects. J. Virol. 2012, 86, 2545–2555. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Hu, Z.; Qin, Q.; Dong, F.; Huang, L.; Long, J.; Chen, P.; Lu, C.; Pan, M. CRISPR/Cas9-mediated disruption of the immediate early-0 and 2 as a therapeutic approach to Bombyx mori nucleopolyhedrovirus in transgenic silkworm. Insect Mol. Biol. 2019, 28, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Dong, Z.Q.; Dong, F.F.; Yu, X.B.; Hu, Z.G.; Liao, N.C.; Chen, P.; Lu, C.; Pan, M.H. Gene editing the BmNPV inhibitor of apoptosis protein 2 (iap2) as an antiviral strategy in transgenic silkworm. Int. J. Biol. Macromol. 2021, 166, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Yu, H.Z.; Geng, L.; Xu, J.P.; Yu, D.; Zhang, S.Z.; Ma, Y.; Fei, D.Q. Comparative Transcriptome Analysis of Bombyx mori (Lepidoptera) Larval Midgut Response to BmNPV in Susceptible and Near-Isogenic Resistant Strains. PLoS ONE 2016, 11, e0155341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Z.H.; Gao, Y.H.; Cheng, S.; Wen, Y.; Tang, X.D.; Li, M.W.; Wu, Y.C.; Wang, X.Y. Identification of the in vitro antiviral effect of BmNedd2-like caspase in response to Bombyx mori nucleopolyhedrovirus infection. J. Invertebr. Pathol. 2021, 183, 107625. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, Z.Q.; Huang, X.M.; Ding, X.Y.; Zhao, C.X.; Li, M.W.; Wu, Y.C.; Liu, Q.N.; Wang, X.Y. Bmcas-1 plays an important role in response against BmNPV infection in vitro. Arch. Insect Biochem. Physiol. 2021, 107, e21793. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Zhang, X.; Chen, D.; Yang, D.; Zhu, C.; Tang, L.; Yang, X.; Wang, Y.; Luo, X.; Wang, M.; et al. CRISPR/Cas9-Mediated Disruption of the lef8 and lef9 to Inhibit Nucleopolyhedrovirus Replication in Silkworms. Viruses 2022, 14, 1119. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061119
Liu Y, Zhang X, Chen D, Yang D, Zhu C, Tang L, Yang X, Wang Y, Luo X, Wang M, et al. CRISPR/Cas9-Mediated Disruption of the lef8 and lef9 to Inhibit Nucleopolyhedrovirus Replication in Silkworms. Viruses. 2022; 14(6):1119. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061119
Chicago/Turabian StyleLiu, Yujia, Xiaoqian Zhang, Dongbin Chen, Dehong Yang, Chenxu Zhu, Linmeng Tang, Xu Yang, Yaohui Wang, Xingyu Luo, Manli Wang, and et al. 2022. "CRISPR/Cas9-Mediated Disruption of the lef8 and lef9 to Inhibit Nucleopolyhedrovirus Replication in Silkworms" Viruses 14, no. 6: 1119. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061119