
Protective Immune Responses Induced by an mRNA-LNP Vaccine Encoding prM-E Proteins against Japanese Encephalitis Virus Infection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses, Cells and Plasmids
2.2. Generation of mRNA and mRNA-LNP
2.3. Denaturing Formaldehyde Gels
2.4. mRNA Transfections
2.5. Western Blotting
2.6. Animal Experiments
2.7. Flow Cytometry
2.8. Plaque Reduction Neutralization Test
2.9. Enzyme-Linked Immunosorbent Assay
2.10. RNA Extraction and Quantitative Real-Time PCR
2.11. Data Analysis and Statistics
3. Results
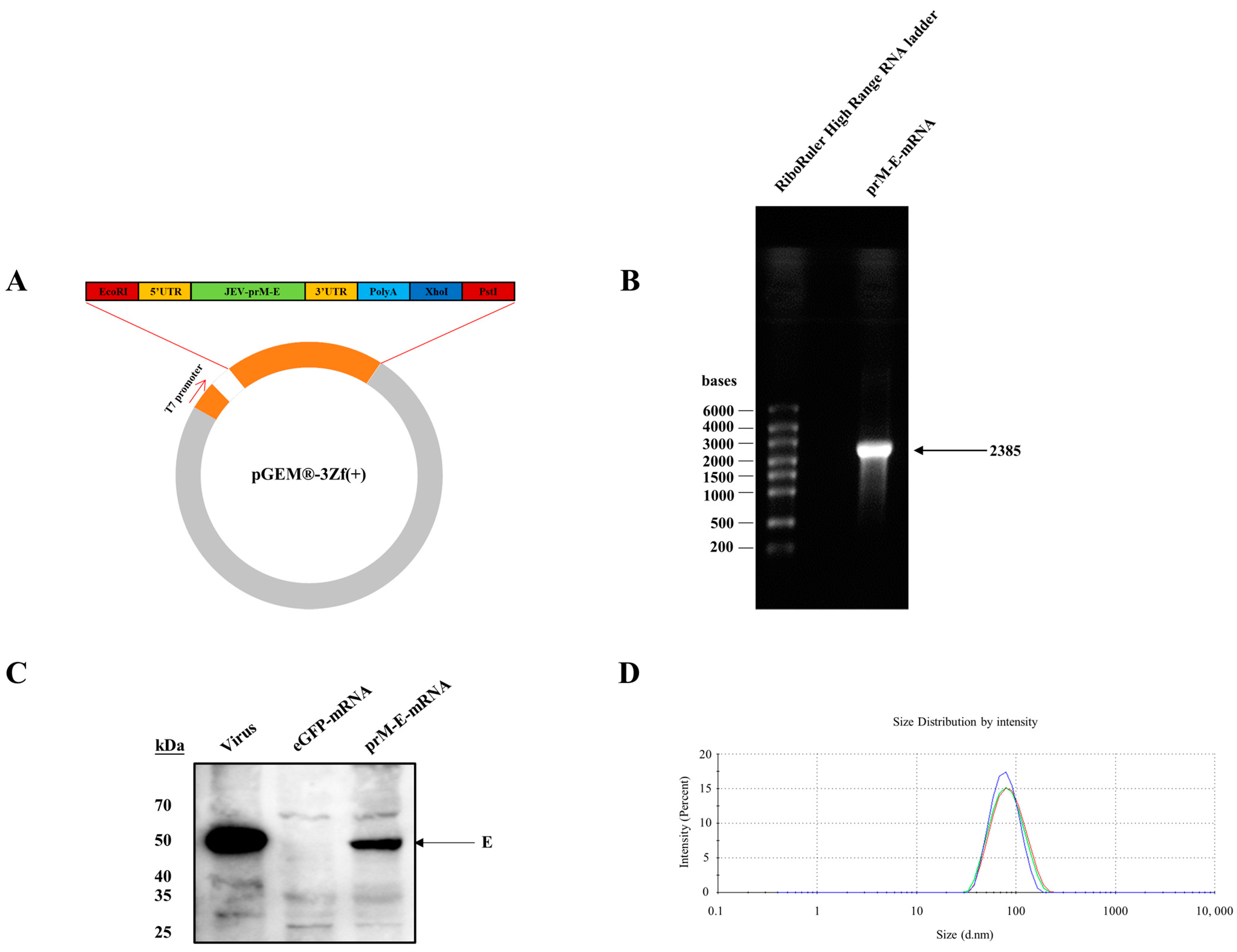
3.1. Construction of the mRNA Vaccine Encoding JEV prM-E Proteins
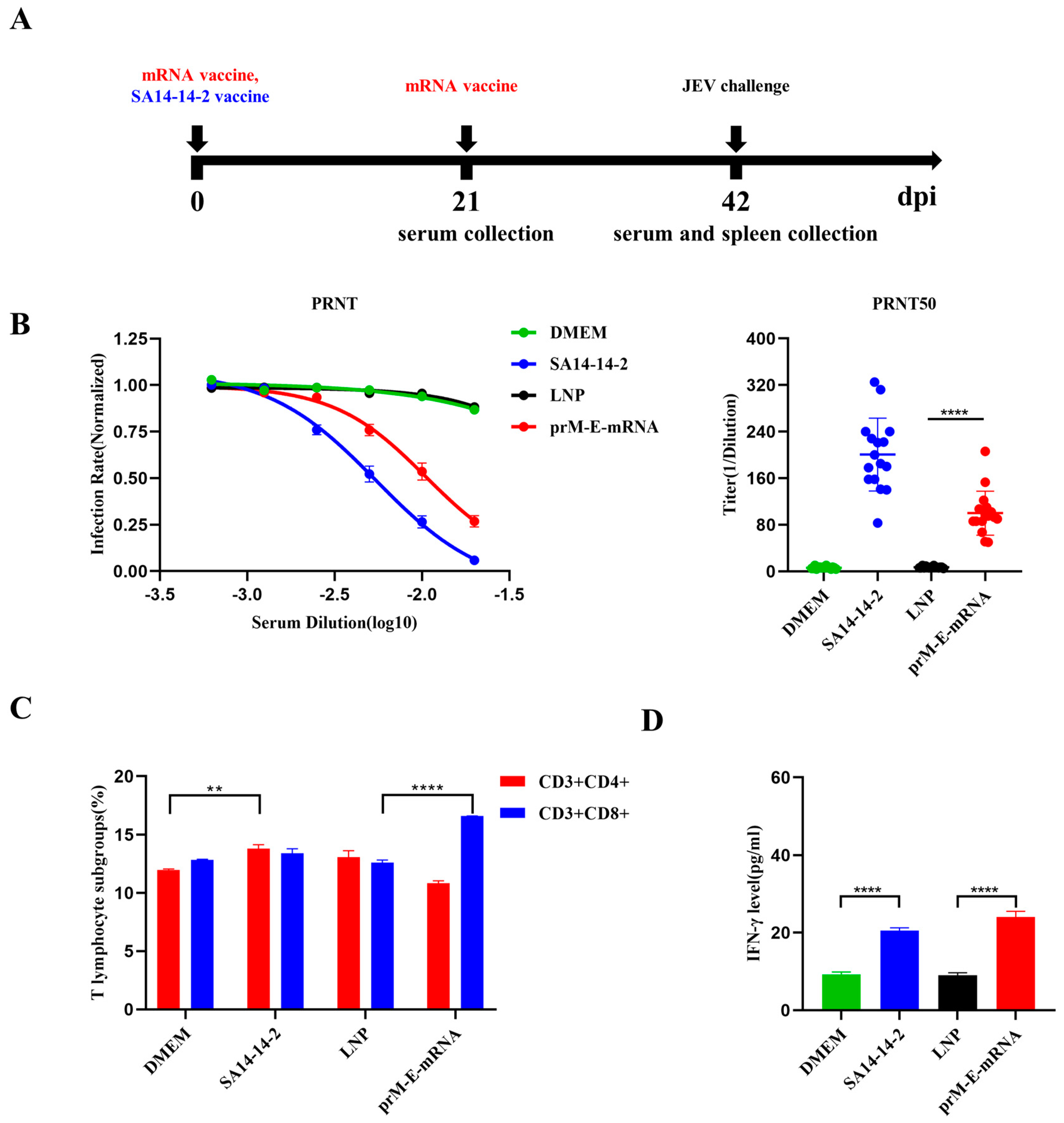
3.2. Immunogenicity of the JEV prM-E-mRNA Vaccine
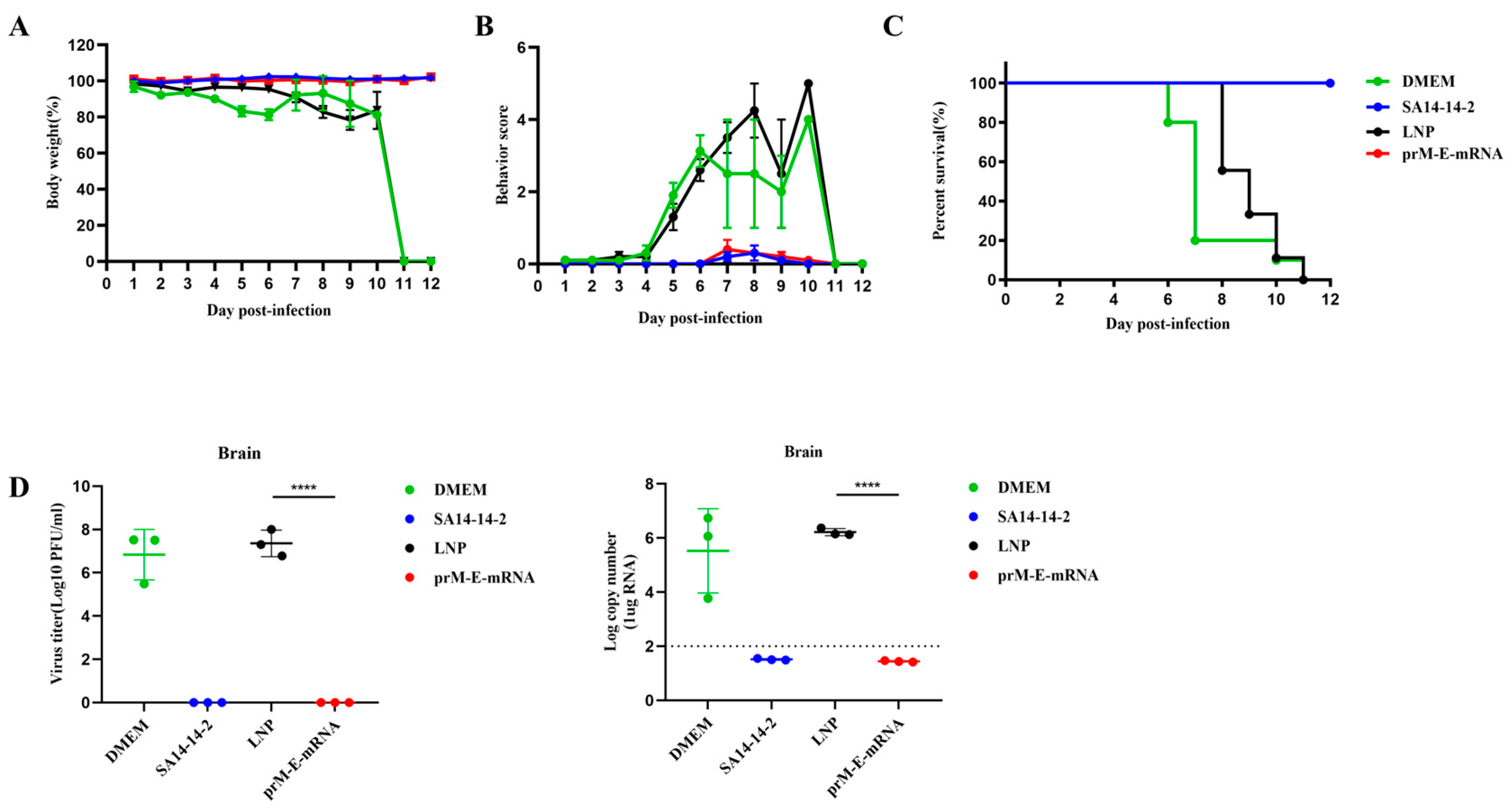
3.3. Protective Effect of the mRNA Vaccine against JEV
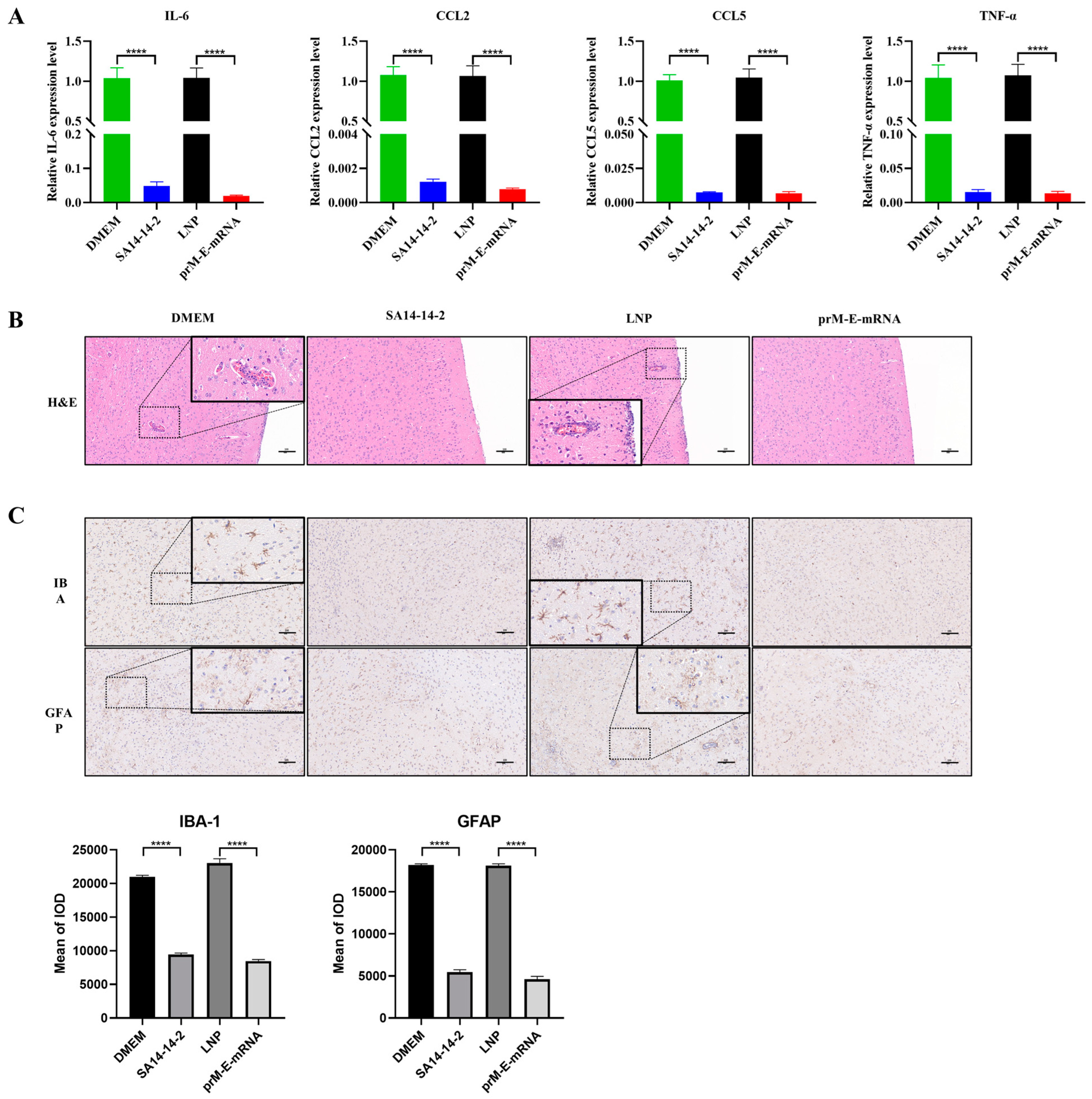
3.4. The mRNA Vaccine Immunization Attenuates JEV-Caused Neuroinflammation Response in Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mackenzie, J.S.; Gubler, D.J.; Petersen, L.R. Emerging flaviviruses: The spread and resurgence of Japanese encephalitis, West Nile and dengue viruses. Nat. Med. 2004, 10 (Suppl. S12), S98–S109. [Google Scholar] [CrossRef] [PubMed]
- Quan, T.M.; Thao, T.T.N.; Duy, N.M.; Nhat, T.M.; Clapham, H. Estimates of the global burden of Japanese encephalitis and the impact of vaccination from 2000–2015. eLife 2020, 9, e51027. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.L.; Lee, P.I. Safety of Japanese encephalitis vaccines. Hum. Vaccin. Immunother. 2021, 17, 4259–4264. [Google Scholar] [CrossRef]
- Wang, H.; Liang, G. Epidemiology of Japanese encephalitis: Past, present, and future prospects. Ther. Clin. Risk Manag. 2015, 11, 435–448. [Google Scholar] [PubMed] [Green Version]
- Wu, D.; Chen, X.; Liu, W.; Fu, S.; Li, F.; Liang, G.; Yang, G.; Zheng, H.; Li, J.; Yin, Z.; et al. Emergence of Japanese encephalitis among adults 40 years of age or older in northern China: Epidemiological and clinical characteristics. Transbound. Emerg. Dis. 2021, 68, 3415–3423. [Google Scholar] [CrossRef]
- Chen, H.L.; Chang, J.K.; Tang, R.B. Current recommendations for the Japanese encephalitis vaccine. J. Chin. Med. Assoc. 2015, 78, 271–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, J.; Wang, T.; Xu, J.; Ouyang, T.; Wang, Q.; Zhang, Y.; Weng, S.; Li, Y.; Wang, Y.; Xin, X.; et al. Novel Japanese encephalitis virus NS1-based vaccine: Truncated NS1 fused with E. coli heat labile enterotoxin B subunit. EBioMedicine 2021, 67, 103353. [Google Scholar] [CrossRef]
- Zhao, F.; Zhai, Y.; Zhu, J.; Xiao, P.; Feng, G. Enhancement of autophagy as a strategy for development of new DNA vaccine candidates against Japanese encephalitis. Vaccine 2019, 37, 5588–5595. [Google Scholar] [CrossRef]
- Kojima, A.; Yasuda, A.; Asanuma, H.; Ishikawa, T.; Takamizawa, A.; Yasui, K.; Kurata, T. Stable high-producer cell clone expressing virus-like particles of the Japanese encephalitis virus e protein for a second-generation subunit vaccine. J. Virol. 2003, 77, 8745–8755. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Wakita, T.; Yasui, K. Inoculation of plasmids encoding Japanese encephalitis virus PrM-E proteins with colloidal gold elicits a protective immune response in BALB/c mice. J. Virol. 2003, 77, 4248–4260. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Yu, X.; Wang, Y.; Turtle, L.; Cui, M.; Wang, R.; Yin, C. Complete protection for mice conferred by a DNA vaccine based on the Japanese encephalitis virus P3 strain used to prepare the inactivated vaccine in China. Virol. J. 2020, 17, 126. [Google Scholar] [CrossRef] [PubMed]
- Pushko, P.; Lukashevich, I.S.; Weaver, S.C.; Tretyakova, I. DNA-launched live-attenuated vaccines for biodefense applications. Expert Rev. Vaccines 2016, 15, 1223–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickols, B.; Tretyakova, I.; Tibbens, A.; Klyushnenkova, E.; Pushko, P. Plasmid DNA launches live-attenuated Japanese encephalitis virus and elicits virus-neutralizing antibodies in BALB/c mice. Virology 2017, 512, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Tombácz, I.; Weissman, D.; Pardi, N. Vaccination with Messenger RNA: A Promising Alternative to DNA Vaccination. Methods Mol. Biol. 2021, 2197, 13–31. [Google Scholar] [PubMed]
- Richner, J.M.; Himansu, S.; Dowd, K.A.; Butler, S.L.; Salazar, V.; Fox, J.M.; Julander, J.G.; Tang, W.W.; Shresta, S.; Pierson, T.C.; et al. Modified mRNA Vaccines Protect against Zika Virus Infection. Cell 2017, 168, 1114–1125.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cafri, G.; Gartner, J.J.; Zaks, T.; Hopson, K.; Levin, N.; Paria, B.C.; Parkhurst, M.R.; Yossef, R.; Lowery, F.J.; Jafferji, M.S.; et al. mRNA vaccine-induced neoantigen-specific T cell immunity in patients with gastrointestinal cancer. J. Clin. Investig. 2020, 130, 5976–5988. [Google Scholar] [CrossRef] [PubMed]
- Krienke, C.; Kolb, L.; Diken, E.; Streuber, M.; Kirchhoff, S.; Bukur, T.; Akilli-Öztürk, Ö.; Kranz, L.M.; Berger, H.; Petschenka, J.; et al. A noninflammatory mRNA vaccine for treatment of experimental autoimmune encephalomyelitis. Science 2021, 371, 145–153. [Google Scholar] [CrossRef]
- Davis, B.S.; Chang, G.J.; Cropp, B.; Roehrig, J.T.; Martin, D.A.; Mitchell, C.J.; Bowen, R.; Bunning, M.L. West Nile virus recombinant DNA vaccine protects mouse and horse from virus challenge and expresses in vitro a noninfectious recombinant antigen that can be used in enzyme-linked immunosorbent assays. J. Virol. 2001, 75, 4040–4047. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, X.; Qi, Y.; Wang, M.; Yu, N.; Nan, F.; Zhang, H.; Tian, M.; Li, C.; Lu, H.; Jin, N. mRNA Vaccines Encoding the HA Protein of Influenza A H1N1 Virus Delivered by Cationic Lipid Nanoparticles Induce Protective Immune Responses in Mice. Vaccines 2020, 8, 123. [Google Scholar] [CrossRef] [Green Version]
- Kling, K.; Harder, T.; Younger, Z.; Burchard, G.; Schmidt-Chanasit, J.; Wichmann, O. Vaccination against Japanese encephalitis with IC51: Systematic review on immunogenicity, duration of protection and safety. J. Travel Med. 2020, 27, taaa016. [Google Scholar] [CrossRef]
- Filgueira, L.; Lannes, N. Review of Emerging Japanese Encephalitis Virus: New Aspects and Concepts about Entry into the Brain and Inter-Cellular Spreading. Pathogens 2019, 8, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladreyt, H.; Durand, B.; Dussart, P.; Chevalier, V. How Central Is the Domestic Pig in the Epidemiological Cycle of Japanese Encephalitis Virus? A Review of Scientific Evidence and Implications for Disease Control. Viruses 2019, 11, 949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.L.; Huang, Y.S.; Lyons, A.C.; Ayers, V.B.; Hettenbach, S.M.; McVey, D.S.; Burton, K.R.; Higgs, S.; Vanlandingham, D.L. North American domestic pigs are susceptible to experimental infection with Japanese encephalitis virus. Sci. Rep. 2018, 8, 7951. [Google Scholar] [CrossRef] [PubMed]
- Ricklin, M.E.; Garcìa-Nicolàs, O.; Brechbühl, D.; Python, S.; Zumkehr, B.; Posthaus, H.; Oevermann, A.; Summerfield, A. Japanese encephalitis virus tropism in experimentally infected pigs. Vet. Res. 2016, 47, 34. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.J.; Lee, S.Y.; Kim, K.H.; Kim, D.S.; Cha, S.H.; Jo, D.S.; Kang, J.H. The Immunogenicity and Safety of the Live-attenuated SA 14-14-2 Japanese Encephalitis Vaccine Given with a Two-dose Primary Schedule in Children. J. Korean Med. Sci. 2015, 30, 612–616. [Google Scholar] [CrossRef] [Green Version]
- Lütticken, D.; Segers, R.P.; Visser, N. Veterinary vaccines for public health and prevention of viral and bacterial zoonotic diseases. Rev. Sci. Tech. 2007, 26, 165–177. [Google Scholar] [CrossRef]
- Firbas, C.; Jilma, B. Product review on the JE vaccine IXIARO. Hum. Vaccin. Immunother. 2015, 11, 411–420. [Google Scholar] [CrossRef]
- Anderson, E.J.; Rouphael, N.G.; Widge, A.T.; Jackson, L.A.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Safety and Immunogenicity of SARS-CoV-2 mRNA-1273 Vaccine in Older Adults. N. Engl. J. Med. 2020, 383, 2427–2438. [Google Scholar] [CrossRef]
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Publisher Correction: Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 2021, 590, E26. [Google Scholar] [CrossRef]
- Li, L.; Lok, S.M.; Yu, I.M.; Zhang, Y.; Kuhn, R.J.; Chen, J.; Rossmann, M.G. The flavivirus precursor membrane-envelope protein complex: Structure and maturation. Science 2008, 319, 1830–1834. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Kuhn, R.J.; Rossmann, M.G. A structural perspective of the flavivirus life cycle. Nat. Rev. Microbiol. 2005, 3, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Rey, F.A.; Heinz, F.X.; Mandl, C.; Kunz, C.; Harrison, S.C. The envelope glycoprotein from tick-borne encephalitis virus at 2 A resolution. Nature 1995, 375, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Konishi, E.; Ajiro, N.; Nukuzuma, C.; Mason, P.W.; Kurane, I. Comparison of protective efficacies of plasmid DNAs encoding Japanese encephalitis virus proteins that induce neutralizing antibody or cytotoxic T lymphocytes in mice. Vaccine 2003, 21, 3675–3683. [Google Scholar] [CrossRef]
- Wu, C.J.; Li, T.L.; Huang, H.W.; Tao, M.H.; Chan, Y.L. Development of an effective Japanese encephalitis virus-specific DNA vaccine. Microbes Infect. 2006, 8, 2578–2586. [Google Scholar] [CrossRef] [PubMed]
- Kalia, A.; Agrawal, M.; Gupta, N. CD8+ T cells are crucial for humoral immunity establishment by SA14-14-2 live attenuated Japanese encephalitis vaccine in mice. Eur. J. Immunol. 2021, 51, 368–379. [Google Scholar] [CrossRef]
- Zhang, N.N.; Li, X.F.; Deng, Y.Q.; Zhao, H.; Huang, Y.J.; Yang, G.; Huang, W.J.; Gao, P.; Zhou, C.; Zhang, R.R.; et al. A Thermostable mRNA Vaccine against COVID-19. Cell 2020, 182, 1271–1283.e16. [Google Scholar] [CrossRef]
- Cao, L.; Fu, S.; Gao, X.; Li, M.; Cui, S.; Li, X.; Cao, Y.; Lei, W.; Lu, Z.; He, Y.; et al. Low Protective Efficacy of the Current Japanese Encephalitis Vaccine against the Emerging Genotype 5 Japanese Encephalitis Virus. PLoS Negl. Trop. Dis. 2016, 10, e0004686. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) |
|---|---|
| prME-F | AGAGCGGCCGCTTTTTCAGCAAGATTAAGCCGCCACCATGA GAGGAGGAAATGAAGGCTCAA |
| prME-R | GGGGTACCTCAAGCATGCACATTGGTCGCTAAGAAC |
| mTNF-α-F | TGTCTCAGCCTCTTCTCATTCC |
| mTNF-α-R | TTAGCCCACTTCTTTCCCTCAC |
| mIL-6-F | AATGAGGAGACTTGCCTGGT |
| mIL-6-R | GCAGGAACTGGATCAGGACT |
| mCCL2-F | CGGCGAGATCAGAACCTACAAC |
| mCCL2-R | GGCACTGTCACACTGGTCACTC |
| mCCL5-F | TGCCCACGTCAAGGAGTATTTC |
| mCCL5-R | AACCCACTTCTTCTCTGGGTTG |
| JEV-F | TGGTTTCATGACCTCGCTCTC |
| JEV-R | CCATGAGGAGTTCTCTGTTTCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Zhu, S.; Wei, N.; Zhao, Z.; Niu, J.; Si, Y.; Cao, S.; Ye, J. Protective Immune Responses Induced by an mRNA-LNP Vaccine Encoding prM-E Proteins against Japanese Encephalitis Virus Infection. Viruses 2022, 14, 1121. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061121
Chen T, Zhu S, Wei N, Zhao Z, Niu J, Si Y, Cao S, Ye J. Protective Immune Responses Induced by an mRNA-LNP Vaccine Encoding prM-E Proteins against Japanese Encephalitis Virus Infection. Viruses. 2022; 14(6):1121. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061121
Chicago/Turabian StyleChen, Tao, Shuo Zhu, Ning Wei, Zikai Zhao, Junjun Niu, Youhui Si, Shengbo Cao, and Jing Ye. 2022. "Protective Immune Responses Induced by an mRNA-LNP Vaccine Encoding prM-E Proteins against Japanese Encephalitis Virus Infection" Viruses 14, no. 6: 1121. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061121