A Naturally Occurring Defective DNA Satellite Associated with a Monopartite Begomovirus: Evidence for Recombination between Alphasatellite and Betasatellite


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
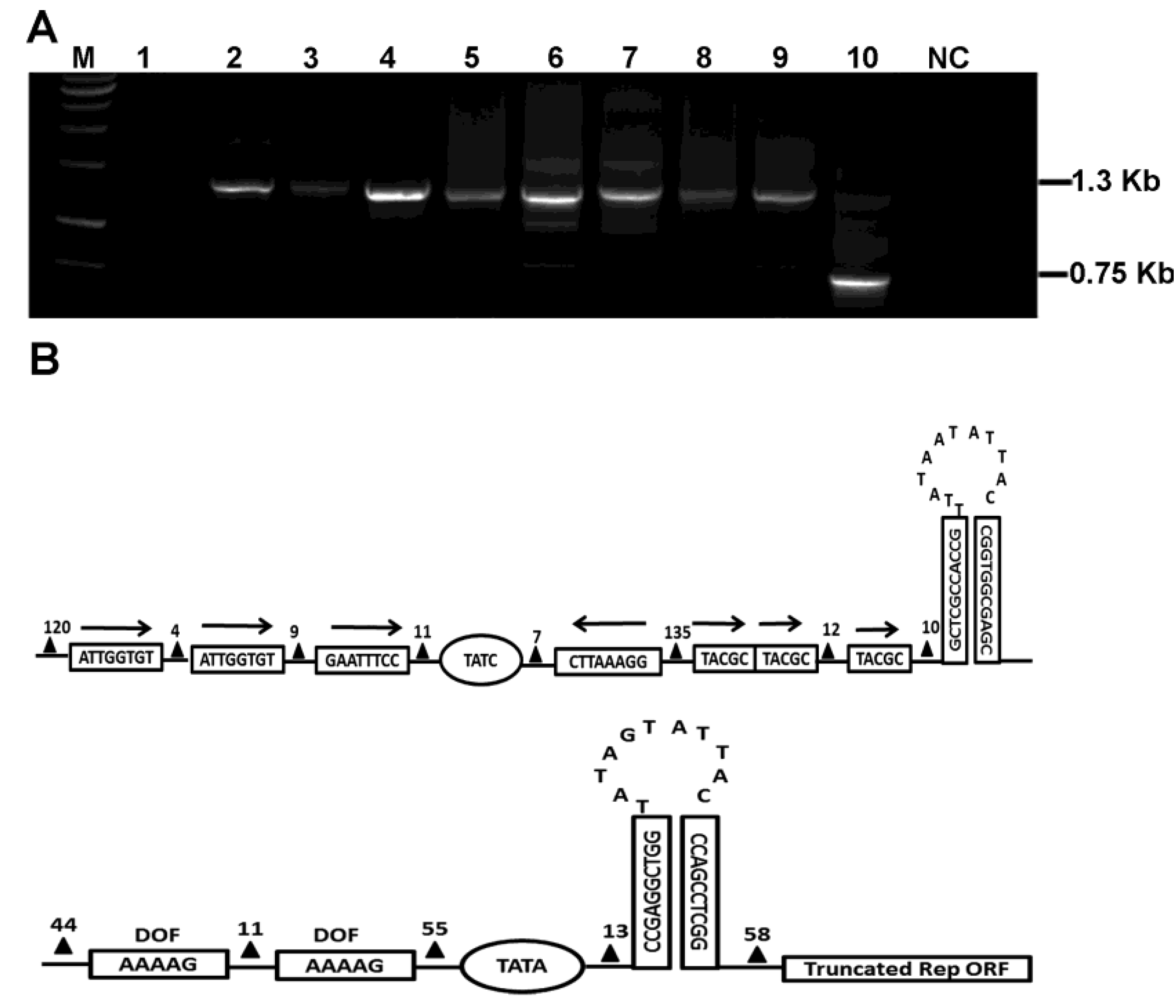
2.1.1. Identification of a Defective Betasatellite Molecule in Association with TbLCYNV-Infected Tobacco Plant
2.1.2. Molecular Characterization of RecSat

2.1.3. Recombination Analysis of RecSat

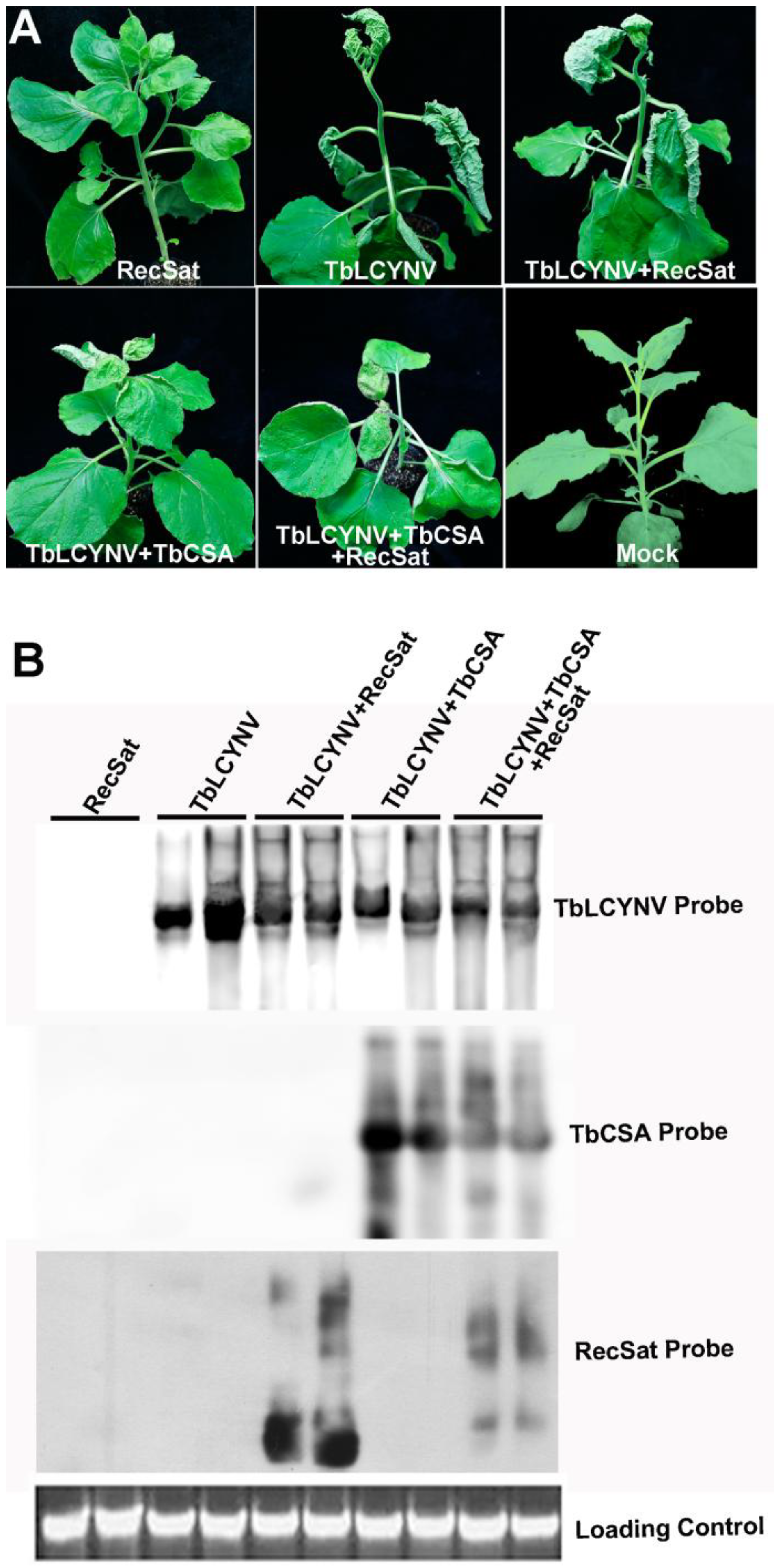
2.1.4. Transreplication of RecSat by TbLCYNV in Nicotiana benthamiana

2.2. Discussion
3. Experimental
3.1. Sources of Virus Isolates
3.2. PCR Amplification and Cloning of RecSat
3.3. Sequence Analysis
3.4. Construction of RecSat Infectious Clone
3.5. Agroinoculation of Plants
3.6. Analysis of Virus and Satellite DNA Accumulation
4. Conclusions
Acknowledgments
Conflicts of Interest
Referencess and Notes
- Harrison, B.; Robinson, D. Natural genomic and antigenic variation in whitefly-transmitted geminiviruses (begomoviruses). Annu. Rev. Phytopathol. 1999, 37, 369–398. [Google Scholar] [CrossRef]
- Mansoor, S.; Briddon, R.W.; Zafar, Y.; Stanley, J. Geminivirus disease complexes: An emerging threat. Trends Plant Sci. 2003, 8, 128–134. [Google Scholar] [CrossRef]
- Mansoor, S.; Zafar, Y.; Briddon, R.W. Geminivirus disease complexes: The threat is spreading. Trends Plant Sci. 2006, 11, 209–212. [Google Scholar] [CrossRef]
- Jeske, H. Geminiviruses. Curr. Top. Microbiol. Immunol. 2009, 331, 185–226. [Google Scholar] [CrossRef]
- Cui, X.F.; Li, G.X.; Wang, D.W.; Hu, D.W.; Zhou, X.P. A begomovirus DNA beta-encoded protein binds DNA, functions as a suppressor of RNA silencing, and targets the cell nucleus. J. Virol. 2005, 79, 10764–10775. [Google Scholar] [CrossRef]
- Cui, X.F.; Tao, X.R.; Xie, Y.; Fauquet, C.M.; Zhou, X.P. A DNA beta associated with Tomato yellow leaf curl China Virus is required for symptom induction. J. Virol. 2004, 78, 13966–13974. [Google Scholar] [CrossRef]
- Eini, O.; Dogra, S.; Selth, L.A.; Dry, I.B.; Randles, J.W.; Rezaian, M.A. Interaction with a host ubiquitin-conjugating enzyme is required for the pathogenicity of a geminiviral DNA beta satellite. Mol. Plant Microbe Interact. 2009, 22, 737–746. [Google Scholar] [CrossRef]
- Shen, Q.; Liu, Z.; Song, F.; Xie, Q.; Hanley-Bowdoin, L.; Zhou, X. Tomato SlSnRK1 protein interacts with and phosphorylates betaC1, a pathogenesis protein encoded by a geminivirus beta-satellite. Plant Physiol. 2011, 157, 1394–1406. [Google Scholar] [CrossRef]
- Yang, X.; Xie, Y.; Raja, P.; Li, S.; Wolf, J.N.; Shen, Q.; Bisaro, D.M.; Zhou, X. Suppression of methylation-mediated transcriptional gene silencing by betaC1-SAHH protein interaction during geminivirus-betasatellite infection. PLoS Pathog. 2011, 7, e1002329. [Google Scholar] [CrossRef]
- Zhou, X. Advances in understanding begomovirus satellites. Annu. Rev. Phytopathol. 2013, 51, 357–381. [Google Scholar] [CrossRef]
- Xie, Y.; Wu, P.; Liu, P.; Gong, H.; Zhou, X. Characterization of alphasatellites associated with monopartite begomovirus/betasatellite complexes in Yunnan, China. Virol. J. 2010, 7, 178. [Google Scholar] [CrossRef]
- Briddon, R.W.; Bull, S.E.; Amin, I.; Mansoor, S.; Bedford, I.D.; Rishi, N.; Siwatch, S.S.; Zafar, Y.; Abdel-Salam, A.M.; Markham, P.G. Diversity of DNA 1: A satellite-like molecule associated with monopartite begomovirus-DNA beta complexes. Virology 2004, 324, 462–474. [Google Scholar] [CrossRef]
- Idris, A.M.; Shahid, M.S.; Briddon, R.W.; Khan, A.J.; Zhu, J.K.; Brown, J.K. An unusual alphasatellite associated with monopartite begomoviruses attenuates symptoms and reduces betasatellite accumulation. J. Gen. Virol. 2011, 92, 706–717. [Google Scholar] [CrossRef]
- Paprotka, T.; Metzler, V.; Jeske, H. The first DNA 1-like alphasatellites in association with New World begomoviruses in natural infections. Virology 2010, 404, 148–157. [Google Scholar] [CrossRef]
- Dry, I.B.; Krake, L.R.; Rigden, J.E.; Rezaian, M.A. A novel subviral agent associated with a geminivirus: The first report of a DNA satellite. Proc. Natl. Acad. Sci. USA 1997, 94, 7088–7093. [Google Scholar] [CrossRef]
- Fiallo-Olive, E.; Martinez-Zubiaur, Y.; Moriones, E.; Navas-Castillo, J. A novel class of DNA satellites associated with New World begomoviruses. Virology 2012, 426, 1–6. [Google Scholar] [CrossRef]
- Ng, T.F.; Duffy, S.; Polston, J.E.; Bixby, E.; Vallad, G.E.; Breitbart, M. Exploring the diversity of plant DNA viruses and their satellites using vector-enabled metagenomics on whiteflies. PLoS One 2011, 6, e19050. [Google Scholar]
- Xie, Y.; Zhou, X.; Li, Z.; Zhang, Z.; Li, G. Identification of a novel DNA molecule associated with Tobacco leaf curl virus. Chinese Sci. Bull. 2002, 47, 1273–1276. [Google Scholar] [CrossRef]
- Zhou, X.P.; Xie, Y.; Peng, Y.; Zhang, Z.K. Malvastrum yellow vein virus, a new Begomovirus species associated with satellite DNA molecule. Chinese Sci. Bull. 2003, 48, 2205–2209. [Google Scholar] [CrossRef]
- Wu, P.J.; Zhou, X.P. Interaction between a nanovirus-like component and the Tobacco curly shoot virus/satellite complex. Acta Bioch. Bioph. Sin. 2005, 37, 25–31. [Google Scholar] [CrossRef]
- Cui, X.F.; Xie, Y.; Zhou, X.P. Molecular characterization of DNAβ molecules associated with Tobacco leaf curl Yunnan Virus. J. Phytopathol. 2004, 152, 647–650. [Google Scholar] [CrossRef]
- Xie, Y.; Jiang, T.; Zhou, X. Agroinoculation Shows Tobacco leaf curl Yunnan virus is a Monopartite Begomovirus. Eur. J.Plant Pathol. 2006, 115, 369–375. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Zhou, X.P.; Liu, Y.L.; Calvert, L.; Munoz, C.; OtimNape, G.W.; Robinson, D.J.; Harrison, B.D. Evidence that DNA-A of a geminivirus associated with severe cassava mosaic disease in Uganda has arisen by interspecific recombination. J. Gen. Virol. 1997, 78, 2101–2111. [Google Scholar]
- Rojas, M.R.; Hagen, C.; Lucas, W.J.; Gilbertson, R.L. Exploiting chinks in the plant's armor: Evolution and emergence of geminiviruses. Annu. Rev. Phytopathol. 2005, 43, 361–394. [Google Scholar] [CrossRef]
- Martin, D.P.; Lefeuvre, P.; Varsani, A.; Hoareau, M.; Semegni, J.Y.; Dijoux, B.; Vincent, C.; Reynaud, B.; Lett, J.M. Complex recombination patterns arising during geminivirus coinfections preserve and demarcate biologically important intra-genome interaction networks. PLoS Pathog. 2011, 7, e1002203. [Google Scholar] [CrossRef] [Green Version]
- Saunders, K.; Bedford, I.D.; Stanley, J. Pathogenicity of a natural recombinant associated with Ageratum yellow vein disease: Implications for geminivirus evolution and disease aetiology. Virology 2001, 282, 38–47. [Google Scholar] [CrossRef]
- Tao, X.; Zhou, X. Pathogenicity of a naturally occurring recombinant DNA satellite associated with tomato yellow leaf curl China virus. J. Gen. Virol. 2008, 89, 306–311. [Google Scholar] [CrossRef]
- Stanley, J.; Saunders, K.; Pinner, M.S.; Wong, S.M. Novel defective interfering DNAs associated with ageratum yellow vein geminivirus infection of Ageratum conyzoides. Virology 1997, 239, 87–96. [Google Scholar] [CrossRef]
- Saunders, K.; Stanley, J. A nanovirus-like DNA component associated with yellow vein disease of Ageratum conyzoides: Evidence for interfamilial recombination between plant DNA viruses. Virology 1999, 264, 142–152. [Google Scholar] [CrossRef]
- Zhou, X.; Xie, Y.; Tao, X.; Zhang, Z.; Li, Z.; Fauquet, C.M. Characterization of DNAbeta associated with begomoviruses in China and evidence for co-evolution with their cognate viral DNA-A. J. Gen. Virol. 2003, 84, 237–247. [Google Scholar] [CrossRef]
- Jiao, X.; Gong, H.; Liu, X.; Xie, Y.; Zhou, X. Etiology of Ageratum yellow vein diseases in South China. Plant Dis. 2013, in press. [Google Scholar]
- Saunders, K.; Briddon, R.W.; Stanley, J. Replication promiscuity of DNA-beta satellites associated with monopartite begomoviruses; deletion mutagenesis of the Ageratum yellow vein virus DNA-beta satellite localizes sequences involved in replication. J. Gen. Virol. 2008, 89, 3165–3172. [Google Scholar] [CrossRef]
- Zulfiqar, A.; Zhang, J.; Cui, X.; Qian, Y.; Zhou, X.; Xie, Y. A new begomovirus associated with alpha- and betasatellite molecules isolated from Vernonia cinerea in China. Arch Virol. 2012, 157, 189–191. [Google Scholar] [CrossRef]
- NCBI. Available online: http://blast.ncbi.nlm.nih.gov (accessed on 4 September 2013).
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Martin, D.P.; Lemey, P.; Lott, M.; Moulton, V.; Posada, D.; Lefeuvre, P. RDP3: A flexible and fast computer program for analyzing recombination. Bioinformatics 2010, 26, 2462–2463. [Google Scholar] [CrossRef]
- van Engelen, F.A.; Molthoff, J.W.; Conner, A.J.; Nap, J.P.; Pereira, A.; Stiekema, W.J. pBINPLUS: An improved plant transformation vector based on pBIN19. Transgenic Res. 1995, 4, 288–290. [Google Scholar] [CrossRef]
- Huang, C.J.; Xie, Y.; Zhou, X.P. Efficient virus-induced gene silencing in plants using a modified geminivirus DNA1 component. Plant Biotechnol. J. 2009, 7, 254–265. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, C.; Xie, Y.; Zhao, L.; Ren, H.; Li, Z. A Naturally Occurring Defective DNA Satellite Associated with a Monopartite Begomovirus: Evidence for Recombination between Alphasatellite and Betasatellite. Viruses 2013, 5, 2116-2128. https://0-doi-org.brum.beds.ac.uk/10.3390/v5092116
Huang C, Xie Y, Zhao L, Ren H, Li Z. A Naturally Occurring Defective DNA Satellite Associated with a Monopartite Begomovirus: Evidence for Recombination between Alphasatellite and Betasatellite. Viruses. 2013; 5(9):2116-2128. https://0-doi-org.brum.beds.ac.uk/10.3390/v5092116
Chicago/Turabian StyleHuang, Changjun, Yan Xie, Liling Zhao, He Ren, and Zhenghe Li. 2013. "A Naturally Occurring Defective DNA Satellite Associated with a Monopartite Begomovirus: Evidence for Recombination between Alphasatellite and Betasatellite" Viruses 5, no. 9: 2116-2128. https://0-doi-org.brum.beds.ac.uk/10.3390/v5092116