
Host Recovery and Reduced Virus Level in the Upper Leaves after Potato virus Y Infection Occur in Tobacco and Tomato but not in Potato Plants

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
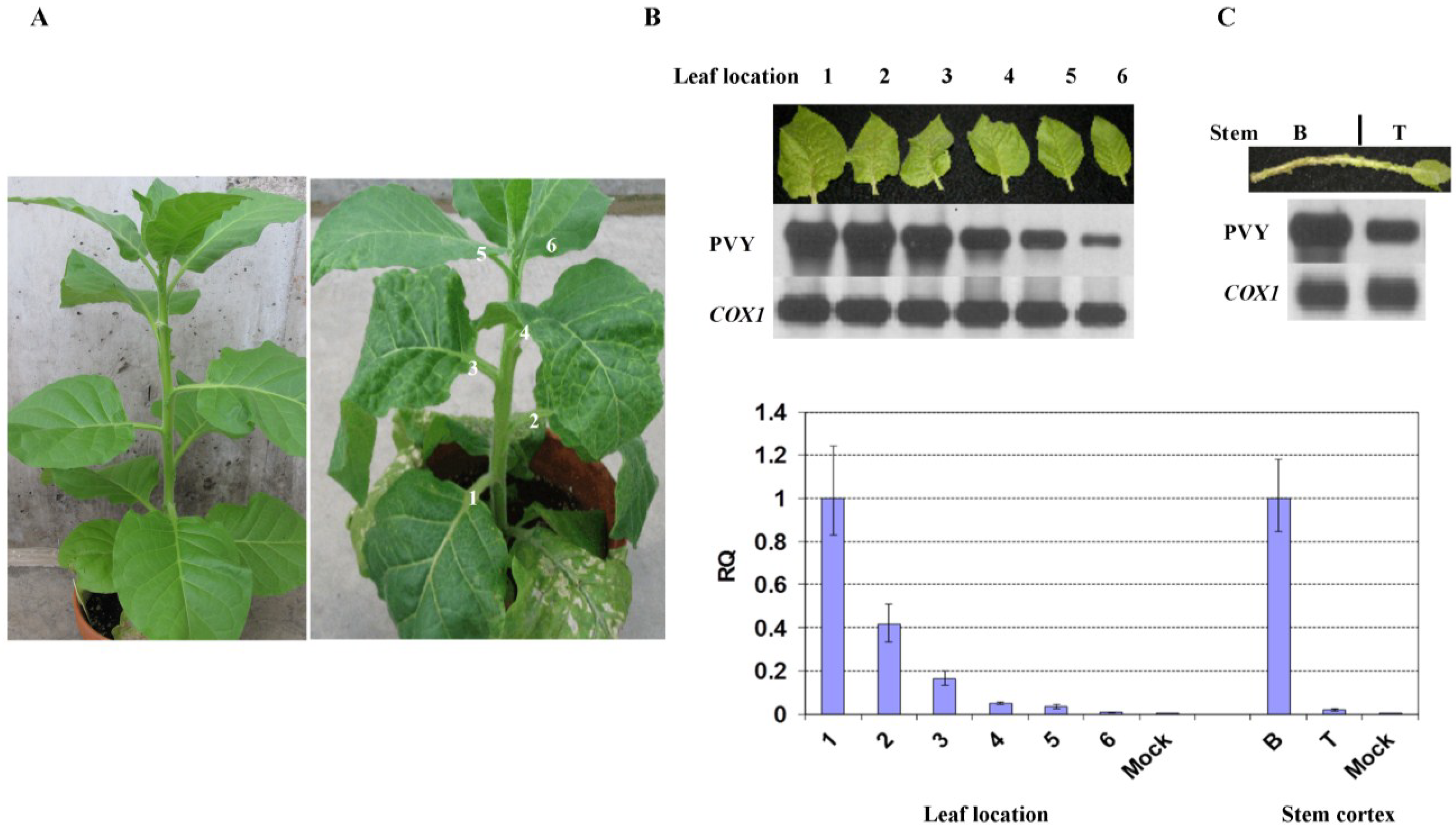
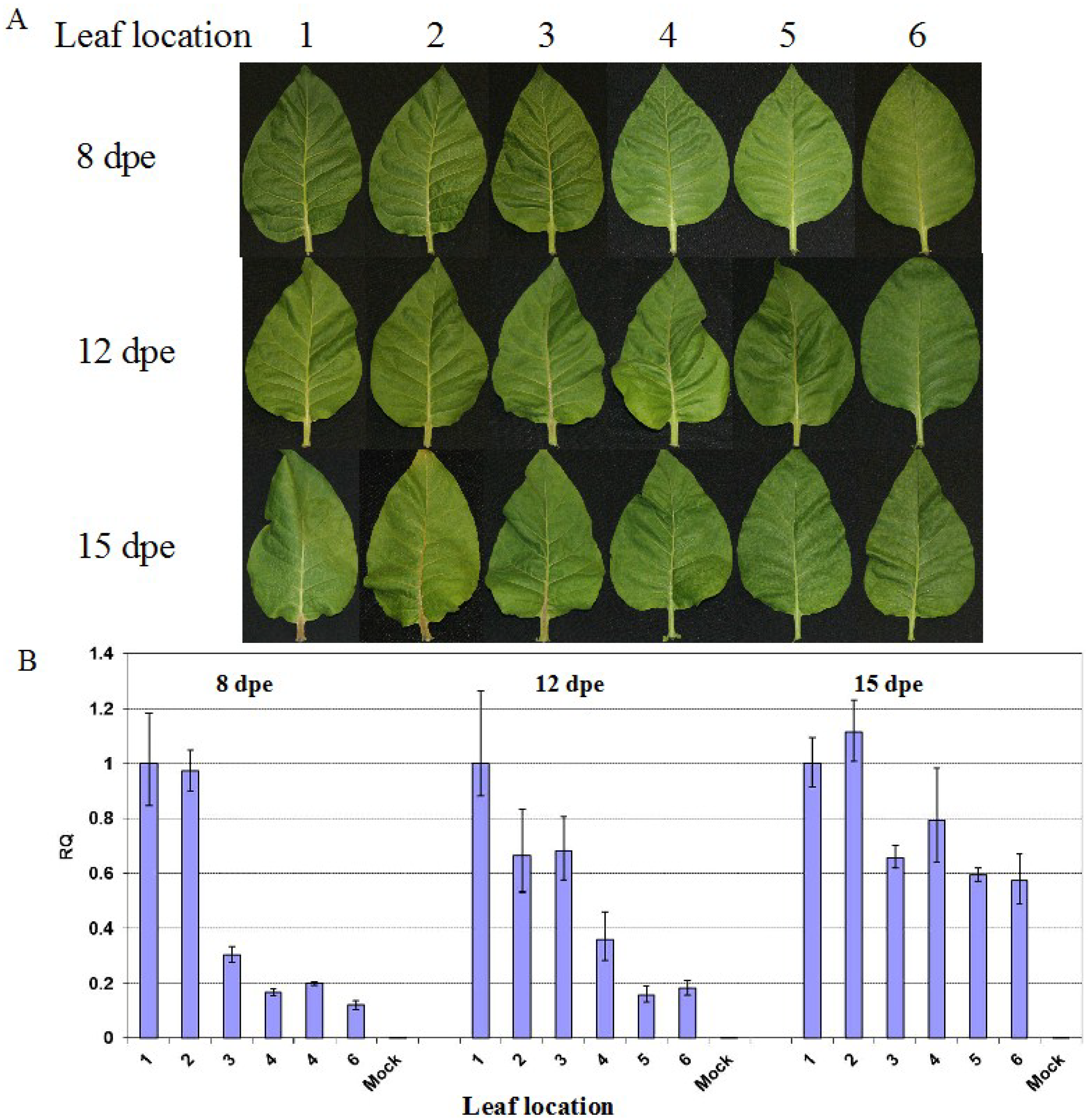
2.1. Reduction in PVY Symptom Severities is Associated with Lower Viral RNA Level


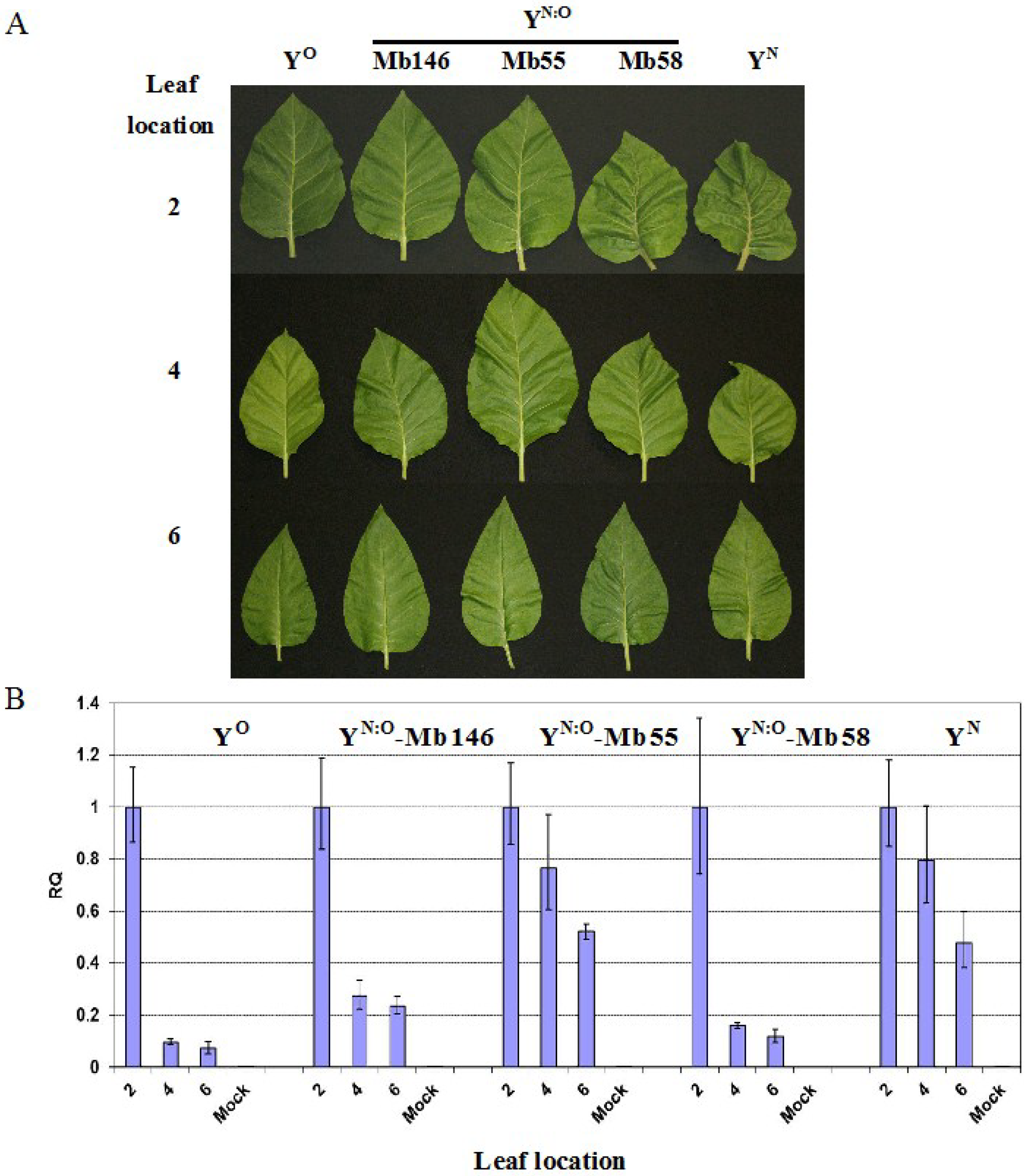
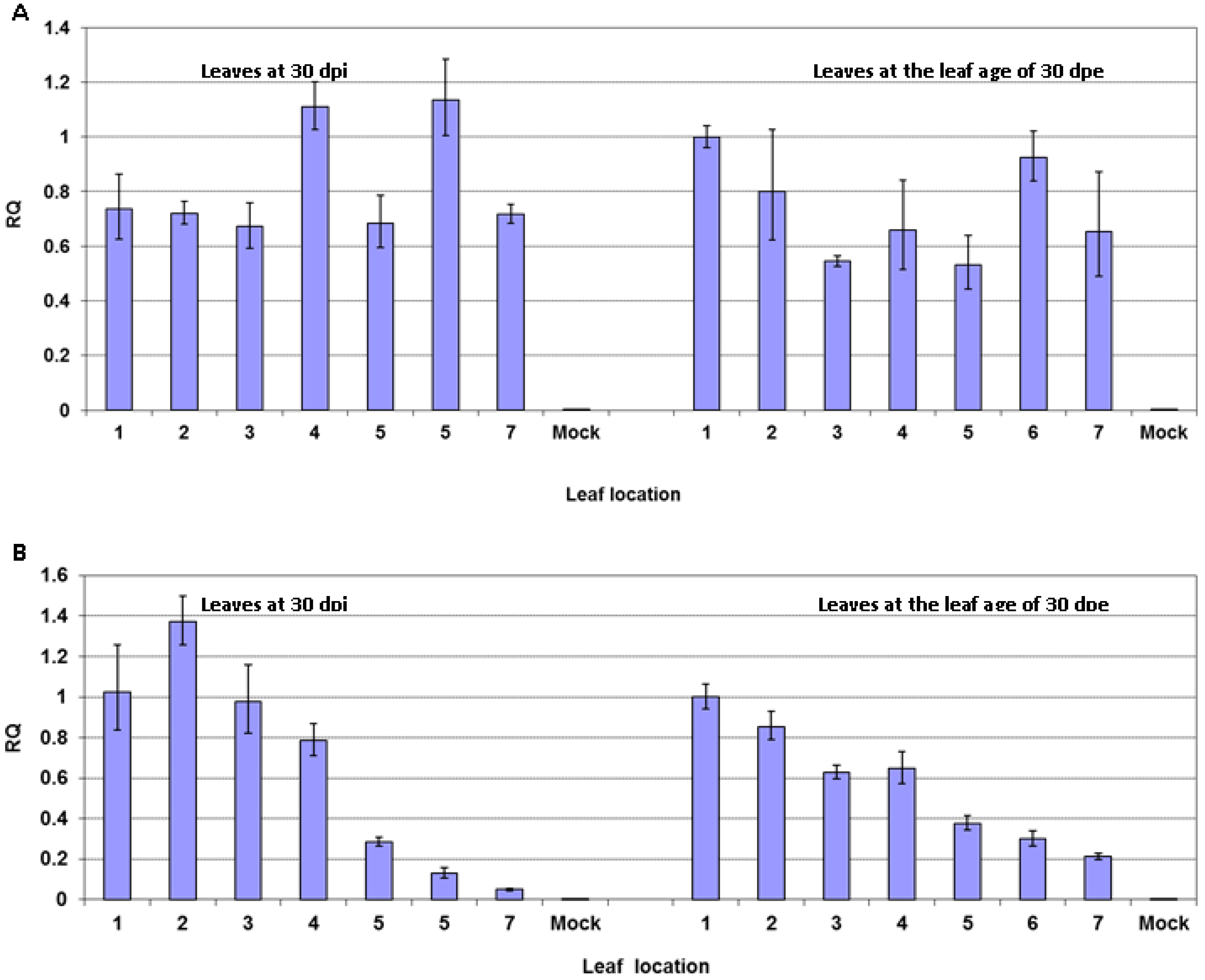
2.2. Reduced Virus Level in Upper Leaves Occurs in Tobacco upon Infection with Various PVY Strains

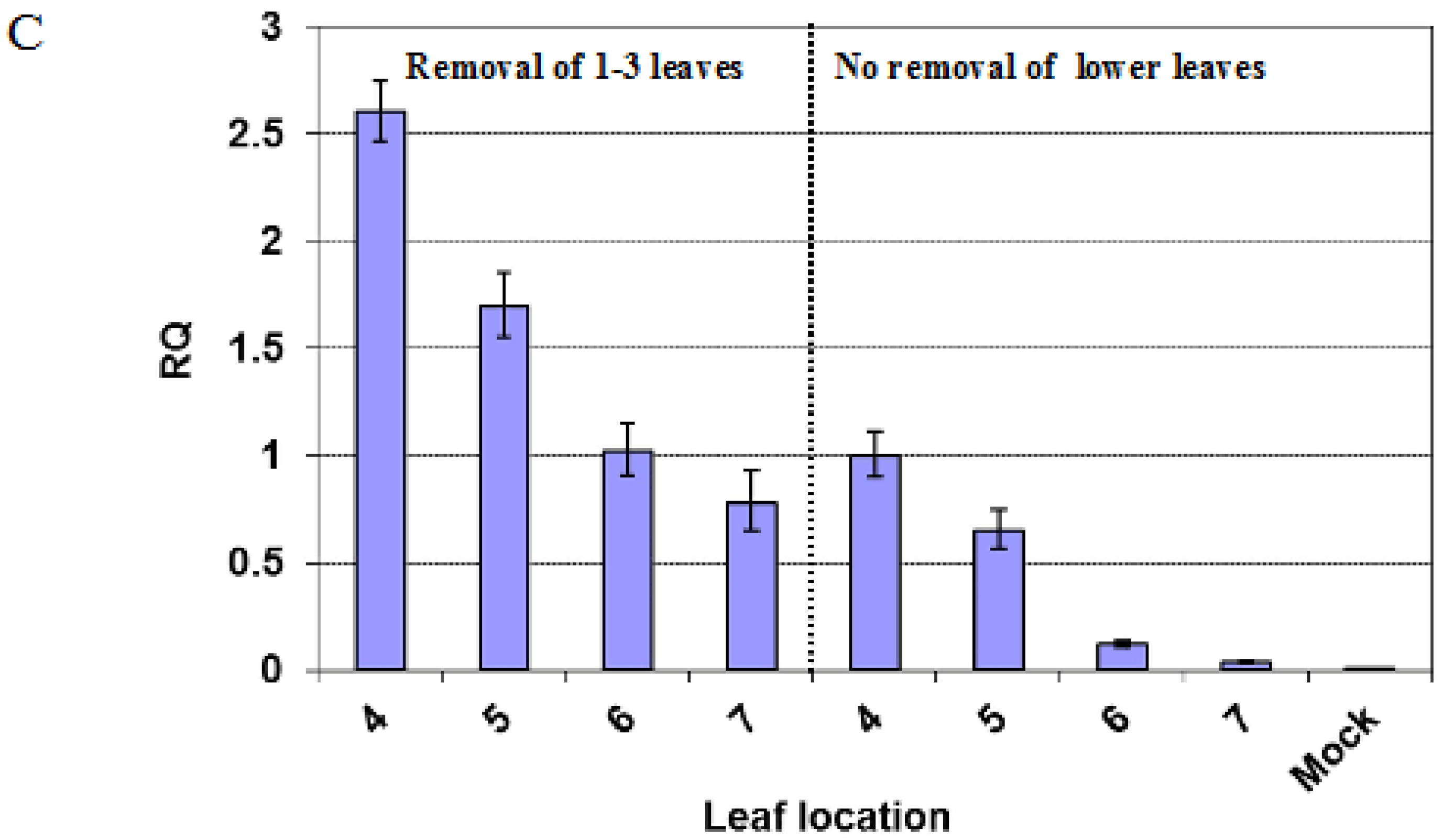
2.3. Removal of the Lower Leaves Affects the Occurrence of Recovery in Upper Leaves


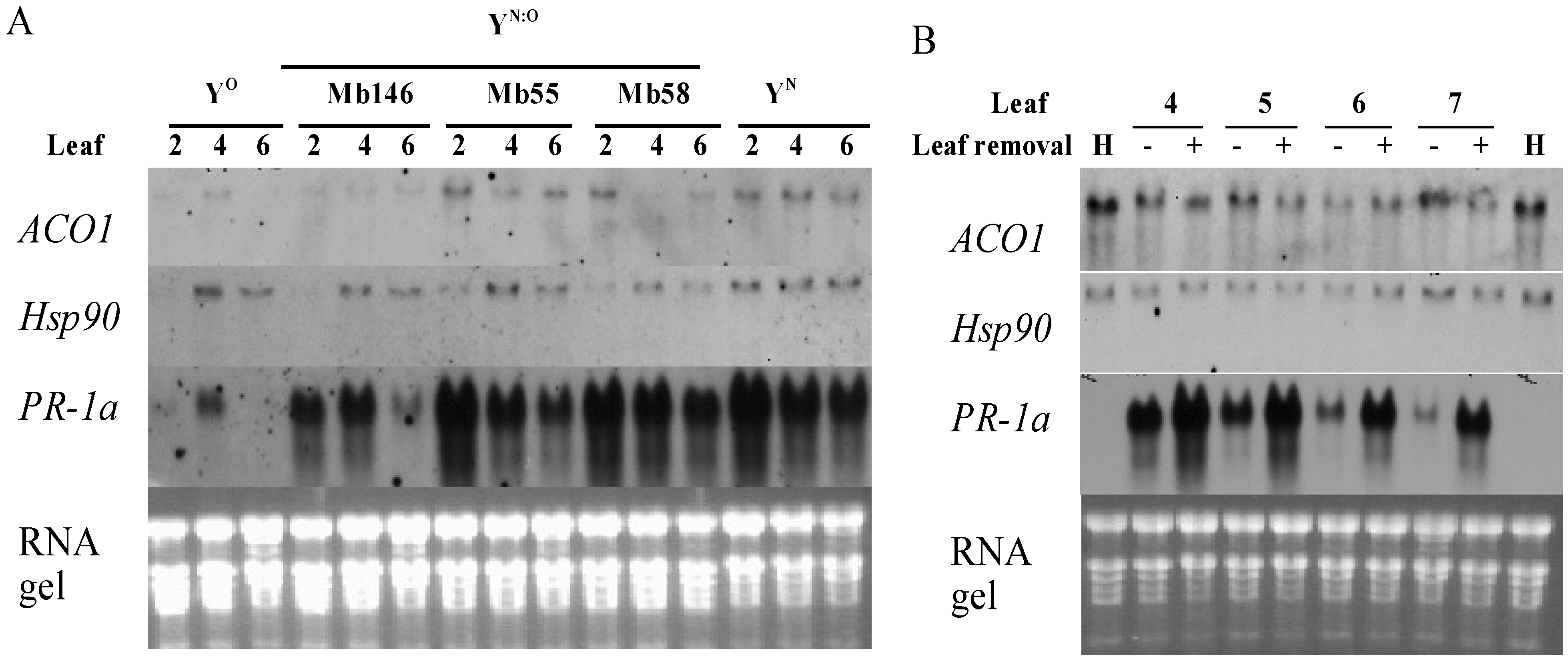
2.4. Recovery is Independent from PR-1a Expression

2.5. Reduced Virus Level in Upper Leaves after PVY Infection Occurs in Tomato but not in Potato Plants

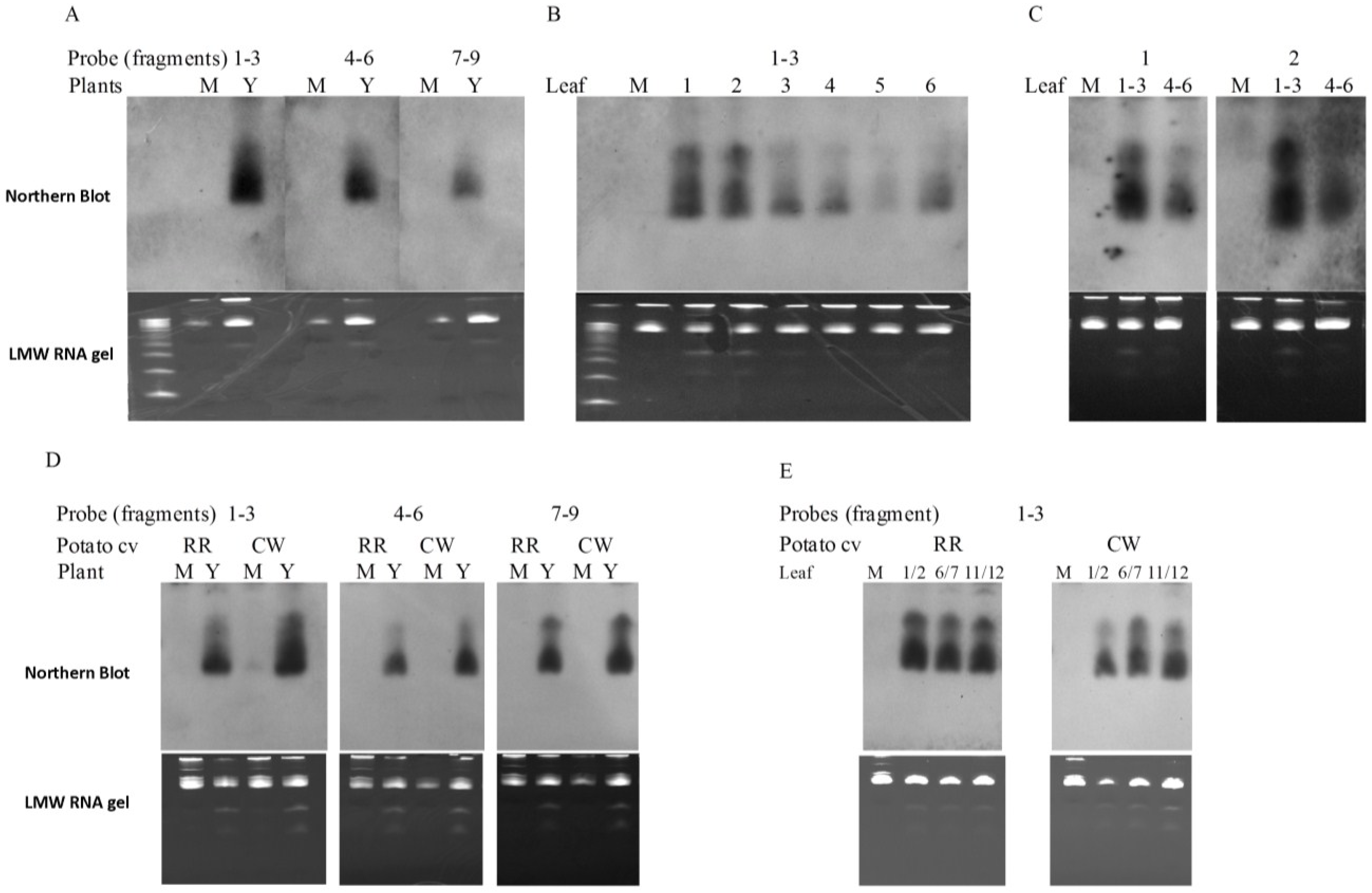
2.6. PVY-Derived siRNAs are Generated in both Tobacco and Potato Plants

3. Discussion
4. Materials and Methods
4.1. Virus Cultures and Plant Materials
4.2. Plant Inoculation and Symptom Observations
4.3. Northern Blot
4.4. Probe Preparation
4.5. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) and Real-Time RT-PCR
4.6. RT-PCR-Southern Blot
4.7. Isolation of Low Molecular Weight (LMW) RNA
4.8. Northern Blot Analysis of Small RNA
5. Conclusions
Acknowledgments
Author contributions
Conflict of Interest
References
- Kang, B.C.; Yeam, I.; Jahn, M.M. Genetics of plant virus resistance. Ann. Rev. Phytopathol. 2005, 43, 581–621. [Google Scholar] [CrossRef]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Gilliland, A.; Murphy, A.M.; Wong, C.E.; Carson, R.A.; Carr, J.P. Mechanisms involved in induced resistance to plant viruses. In Multigenic and Induced Systemic Resistance in Plants; Tuzun, S., Bent, E., Eds.; Springer: New York, NY, USA, 2006; pp. 335–359. [Google Scholar]
- Lewsey, M.G.; Carr, J.P. Effects of DICER-like proteins 2, 3 and 4 on cucumber mosaic virus and tobacco mosaic virus infections in salicylic acid-treated plants. J. Gen. Virol. 2009, 90, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Nie, X. Salicylic acid suppresses Potato virus Y isolate N:O-induced symptoms in tobacco plants. Phytopathology 2006, 96, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Hamilton, A.; Voinnet, O.; Chappell, L.; Baulcombe, D. Two classes of short interfering RNA in RNA silencing. EMBO J. 2002, 21, 4671–4679. [Google Scholar] [CrossRef] [PubMed]
- Kundu, J.K.; Briard, P.; Hily, J.M.; Ravelonandro, M.; Scorza, R. Role of the 25–26 nt siRNA in the resistance of transgenic Prunus domestica graft inoculated with plum pox virus. Virus Genes 2008, 36, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ferrer, V.; Voinnet, O. Roles of plant small RNAs in biotic stress responses. Annu. Rev. Plant Biol. 2009, 60, 485–510. [Google Scholar] [CrossRef] [PubMed]
- Moazed, D. Small RNAs in transcriptional gene silencing and genome defence. Nature 2009, 457, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Wingard, S.A. Hosts and symptoms of ring spot, a virus disease of plants. J. Agric. Res. 1928, 37, 127–153. [Google Scholar]
- Chellappan, P.; Vanitharani, R.; Fauquet, C.M. Short interfering RNA accumulation correlates with host recovery in DNA virus-infected hosts, and gene silencing targets specific viral sequences. J. Virol. 2004, 78, 7465–7477. [Google Scholar] [CrossRef] [PubMed]
- Jovel, J.; Walker, M.; Sanfaçon, H. Recovery of Nicotiana benthamiana plants from a necrotic response induced by a nepovirus is associated with RNA silencing but not with reduced virus titer. J. Virol. 2007, 81, 12285–12297. [Google Scholar] [CrossRef] [PubMed]
- Jovel, J.; Walker, M.; Sanfaçon, H. Salicylic acid-dependent restriction of Tomato ringspot virus spread in tobacco is accompanied by a hypersensitive response, local RNA silencing, and moderate systemic resistance. Mol. Plant-Microbe Interact. 2011, 24, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Negrete, E.A.; Carrillo-Tripp, J.; Rivera-Bustamante, R.F. RNA silencing against geminivirus: Complementary action of posttranscriptional gene silencing and transcriptional gene silencing in host recovery. J. Virol. 2009, 83, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.; de Boer, S.; Lorenzen, J.; Karasev, A.; Whitworth, J.; Nolte, P.; Singh, R.; Boucher, A.; Xu, H. Potato virus Y: An evolving concern for potato crops in the United States and Canada. Plant Dis. 2010, 94, 1384–1397. [Google Scholar] [CrossRef]
- Karasev, A.V.; Gray, S.M. Continuous and emerging challenges of Potato virus Y in potato. Annu. Rev. Phytopathol. 2013, 51, 571–586. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Singh, M.; Pelletier, Y.; McLaren, D. Recent advances on Potato virus Y research in Canada. Am. J. Potato Res. 2013, 90, 14–20. [Google Scholar] [CrossRef]
- Nie, X.; Singh, R.P.; Singh, M. Molecular and pathological characterization of N:O isolates of the Potato virus Y from Manitoba, Canada. Can. J. Plant Pathol. 2004, 26, 573–583. [Google Scholar] [CrossRef]
- Hu, X.; Nie, X.; He, C.; Xiong, X. Differential pathogenicity of two different recombinant PVYNTN isolates in Physalis floridana is determined by the coat protein gene. Virol. J. 2011, 8, 207. [Google Scholar] [CrossRef] [PubMed]
- Nie, B.; Singh, M.; Sullivan, A.; Singh, R.P.; Xie, C.; Nie, X. Recognition and molecular discrimination of severe and mild PVY° variants of Potato virus Y in potatoes in New Brunswick, Canada. Plant Dis. 2011, 95, 113–119. [Google Scholar] [CrossRef]
- Nie, B.; Singh, M.; Sullivan, A.; Murphy, A.; Xie, C.; Nie, X. Response of potato cultivars to five isolates belonging to four strains of Potato virus Y. Plant Dis. 2012, 96, 1422–1429. [Google Scholar] [CrossRef]
- Nie, X.; Singh, R.P. Evolution of North American PVYNTN strain Tu 660 from local PVYN by mutation rather than recombination. Virus Genes 2003, 26, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Chellappan, P.; Vanitharani, R.; Ogbe, F.; Fauquet, CM. Effect of temperature on geminivirus-induced RNA silencing in plants. Plant Physiol. 2005, 138, 1828–1841. [Google Scholar] [CrossRef] [PubMed]
- De Koeyer, D.; Nie, X.; Douglass, K.; Lagüe, M.; Gustafson, V.; Albert, L.; Singh, M. Next generation sequencing for detection of viruses infecting potato. In Proceedings of the 96th Annual Meeting of the Potato Association of America, Denver, CO, USA, 12–16 August 2012.
- Nolte, P.; Whitworth, J.L.; Thornton, M.K.; McIntosh, C.S. Effect of seedborne Potato virus Y on performance of Russet Burbank, Russet Norkotah, and Shepody potato. Plant Dis. 2004, 88, 248–252. [Google Scholar] [CrossRef]
- Carr, J.P.; Beachy, R.N.; Klessig, D.F. Are the PR1 proteins of tobacco involved in genetically engineered resistance to TMV? Virology 1989, 169, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Singh, R.P.; Tai, G.C.C. Molecular characterization and expression analysis of 1-aminocylcopropane-1-carboxylate oxidase homologs from potato under abiotic and biotic stresses. Genome 2002, 45, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Alamillo, J.M.; Saenz, P.; Garcia, J.A. Salicylic acid-mediated and RNA-silencing defense mechanisms cooperate in the restriction of systemic spread of plum pox virus in tobacco. Plant J. 2006, 48, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, V.; Scholthof, H.B. Plant responses against invasive nucleic acids: RNA silencing and its suppression by plant viral pathogens. Semin. Cell Dev. Biol. 2009, 20, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Dunoyer, P.; Schott, G.; Himber, C.; Meyer, D.; Takeda, A.; Carrington, J.C.; Voinnet, O. Small RNA duplexes function as mobile silencing signals between plant cells. Science 2010, 328, 912–916. [Google Scholar] [CrossRef] [PubMed]
- Mlotshwa, S.; Pruss, G.J.; Peragine, A.; Endres, M.W.; Li, J.; Chen, X.; Poethig, R.S.; Bowman, L.H.; Vance, V. DICER-LIKE2 plays a primary role in transitive silencing of transgenes in Arabidopsis. PLoS One 2008, 3, e1755. [Google Scholar] [CrossRef] [PubMed]
- Molnar, A.; Melnyk, C.W.; Bassett, A.; Hardcastle, T.J.; Dunn, R.; Baulcombe, D.C. Small silencing RNAs in plants are mobile and direct epigenetic modification in recipient cells. Science 2010, 328, 872–875. [Google Scholar] [CrossRef] [PubMed]
- Szittya, G.; Molnár, A.; Silhavy, D.; Hornyik, C.; Burgyán, J. Short defective interfering RNAs of tombusviruses are not targeted but trigger post-transcriptional gene silencing against their helper virus. Plant Cell 2002, 14, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.W. RNA-based antiviral immunity. Nat. Rev. Immunol. 2010, 10, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed]
- Schuck, J.; Gursinsky, T.; Pantaleo, V.; Burgyán, J.; Behrens, S.E. AGO/RISC-mediated antiviral RNA silencing in a plant in vitro system. Nucleic Acids Res. 2013, 41, 5090–5103. [Google Scholar] [CrossRef] [PubMed]
- Palauqui, J.C.; Elmayan, T.; Pollien, J.M.; Vaucheret, H. Systemic acquired silencing: Transgene-specific post-transcriptional silencing is transmitted by grafting from silenced stocks to non-silenced scions. EMBO J. 1997, 16, 4738–4745. [Google Scholar] [CrossRef] [PubMed]
- Donaire, L.; Wang, Y.; Gonzalez-Ibeas, D.; Mayer, K.F.; Aranda, M.A.; Llave, C. Deep-sequencing of plant viral small RNAs reveals effective and widespread targeting of viral genomes. Virology 2009, 392, 203–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llave, C. Virus-derived small interfering RNAs at the core of plant–virus interactions. Trends Plant Sci. 2010, 15, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Bao, F.S.; Xie, Z. Small RNA deep sequencing reveals role for Arabidopsis thaliana RNA-dependent RNA polymerases in viral siRNA biogenesis. PLoS One 2009, 4, e4971. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Wu, Q.; Ito, T.; Cillo, F.; Li, W.X.; Chen, X.; Yu, J.L.; Ding, S.W. RNAi-mediated viral immunity requires amplification of virus-derived siRNAs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Luo, Y.; Lu, R.; Lau, N.; Lai, E.C.; Li, W.X.; Ding, S.W. Virus discovery by deep sequencing and assembly of virus-derived small silencing RNAs. Proc. Natl. Acad. Sci. USA 2010, 107, 1606–1611. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Singh, R.P. A new approach for the simultaneous differentiation of biological and geographical strains of Potato virus Y by uniplex and multiplex RT-PCR. J. Virol. Methods 2002, 104, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Singh, R.P. Specific differentiation of recombinant PVYN:° and PVYNTN strains by multiplex RT-PCR. J. Virol. Methods 2003, 113, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, S.S.; Poole, R.J.; Dhindsa, R.S. Changes in protein patterns and translatable messenger RNA populations during cold acclimation of alfalfa. Plant Physiol. 1987, 84, 1172–1176. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular cloning: A laboratory manual, 2nd ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Balme-Sinibaldi, V.; Tribodet, M.; Croizat, F.; Lefeuvre, P.; Kerlan, C.; Jacquot, E. Improvement of Potato virus Y (PVY) detection and quantitation using PVYN-and PVY°-specific real-time RT-PCR assays. J. Virol. Methods 2006, 134, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Singh, R.P. Differential accumulation of Potato virus A and expression of pathogenesis-related genes in resistant potato cv. Shepody upon graft inoculation. Phytopathology 2001, 91, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Bookout, A.L.; Mangelsdorf, D.J. Quantitative real-time PCR protocol for analysis of nuclear receptor signaling pathways. Nucl. Recept. Signal. 2003, 1, e012. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, X.; Molen, T.A. Host Recovery and Reduced Virus Level in the Upper Leaves after Potato virus Y Infection Occur in Tobacco and Tomato but not in Potato Plants. Viruses 2015, 7, 680-698. https://0-doi-org.brum.beds.ac.uk/10.3390/v7020680
Nie X, Molen TA. Host Recovery and Reduced Virus Level in the Upper Leaves after Potato virus Y Infection Occur in Tobacco and Tomato but not in Potato Plants. Viruses. 2015; 7(2):680-698. https://0-doi-org.brum.beds.ac.uk/10.3390/v7020680
Chicago/Turabian StyleNie, Xianzhou, and Teresa A. Molen. 2015. "Host Recovery and Reduced Virus Level in the Upper Leaves after Potato virus Y Infection Occur in Tobacco and Tomato but not in Potato Plants" Viruses 7, no. 2: 680-698. https://0-doi-org.brum.beds.ac.uk/10.3390/v7020680