
Facilitation of Rice Stripe Virus Accumulation in the Insect Vector by Himetobi P Virus VP1


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Virus
2.2. Yeast Two-Hybrid Screen
2.3. Quantitative Detection of HiPV and RSV in a Single Insect
2.4. Synthesis of dsRNA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer and Purpose | Sequence (5’→3’) | Modification |
|---|---|---|
| Construction for Yeast Two-Hybrid Assay | ||
| VP1-F | CGAATTCGCTAACTTTGCGTCTACT | EcoR I |
| VP1-R | AGGATCCCTAAACTTTGTCAAAGG | BamH I |
| RNP-F | GCATATGATGGGTACCAACAAGCC | Nde I |
| RNP-R | TGGATCCCTAGTCATCTGCACCTT | BamH I |
| For qRT-PCR | ||
| qVP1-F | GGTCCAGGGTGCTTTGATTG | |
| qVP1-R | ACTGATGGTTGTGATGCGTG | |
| qRNP-F | TGCAGAAGGCAATCAATGACAT | |
| qRNP-R | TGTCACCACCTTTGTCCTTCAA | |
| qActin-F | TCTTGAGATTGGACTTGGC | |
| qActin-R | GTAGCACAGTTTCACCTTG | |
| For dsRNA Synthesis | ||
| T7-VP1-F | TAATACGACTCACTATAGGGATCCTTATCCTGTGTAGGAG | |
| T7-VP1-R | TAATACGACTCACTATAGGGAGTTAAATATGTGGGGCAAT | |
| T7-GFP-F | TAATACGACTCACTATAGGGAGTGGAGAGGGTGAAGG | |
| T7-GFP-R | TAATACGACTCACTATAGGGAGGGCAGATTGTGTGGAC | |
2.5. Feeding-Based RNA Interference (RNAi) Analysis
2.6. RSV Accumulation Analysis
2.7. Virus Acquisition Experiments on SBPH
3. Results
3.1. Interactions between HiPV-VP1 and RSV-RNP

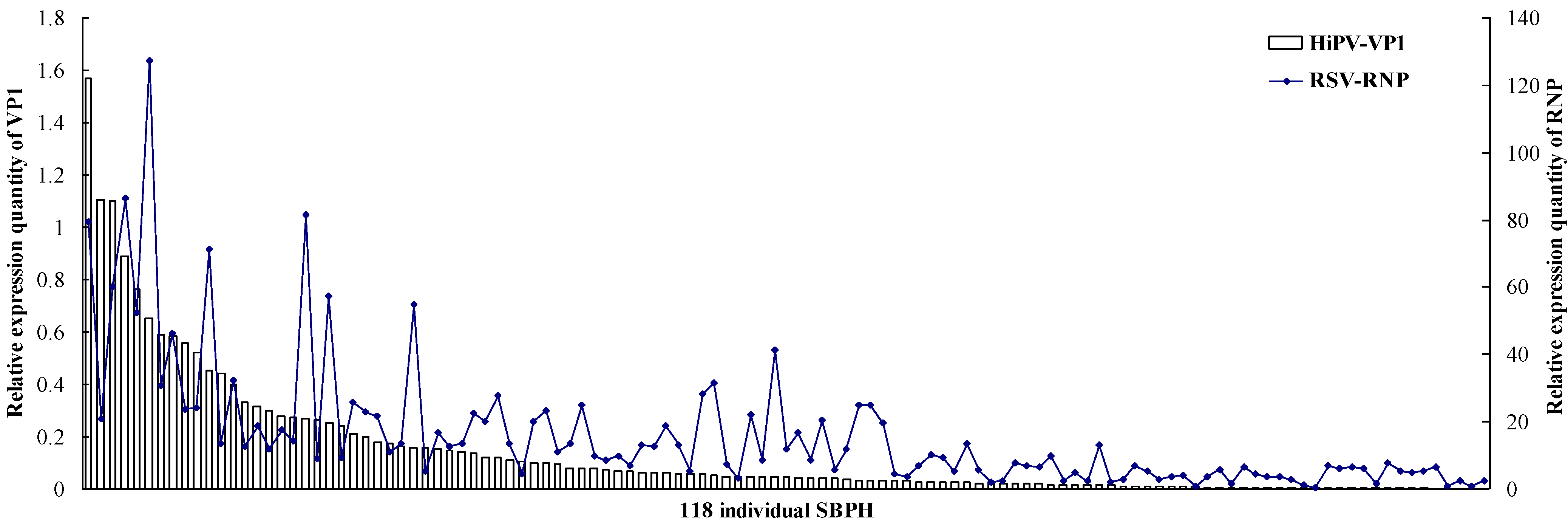
3.2. Titers of HiPV and RSV in a Single SBPH

| Group | The Number of Insects | |||
|---|---|---|---|---|
| Higher RSV Accumulation | Middle RSV Accumulation | Lower RSV Accumulation | Total | |
| First group | 33 (89.2%) | 3 (8.1%) | 1 (2.7%) | 37 |
| Second group | 23 (45.1%) | 19 (37.3%) | 9 (17.6%) | 51 |
| Third group | 0 | 11 (36.7%) | 19 (63.3%) | 30 |
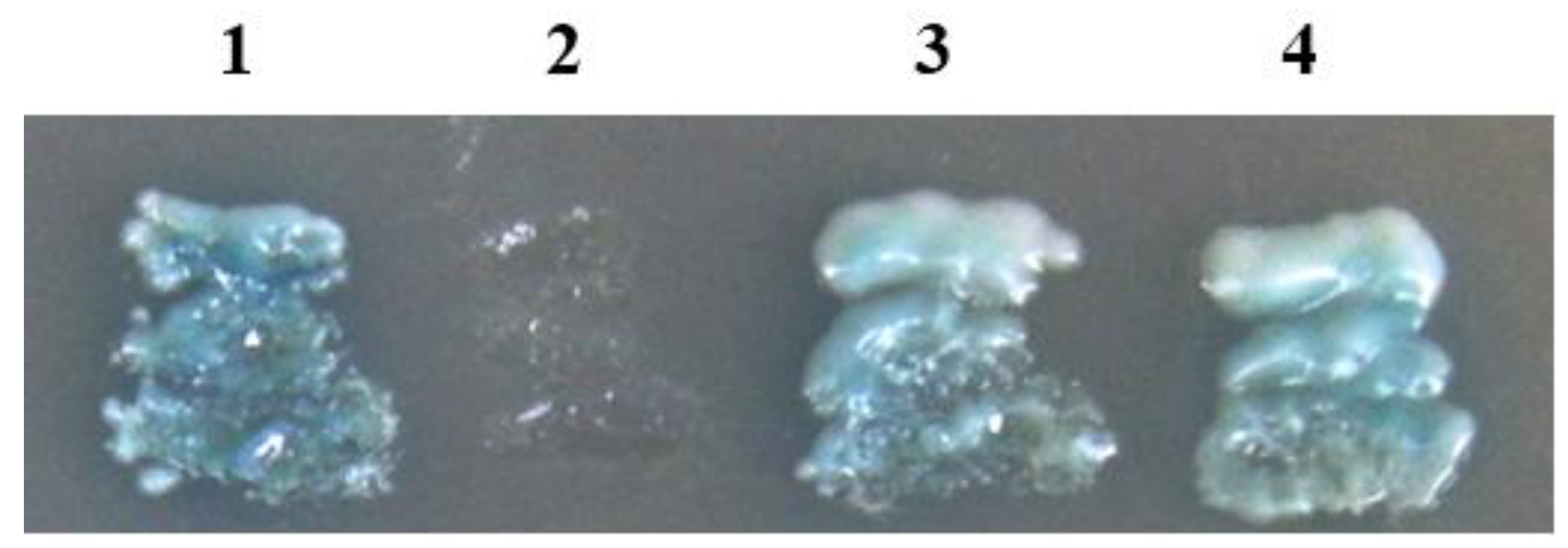
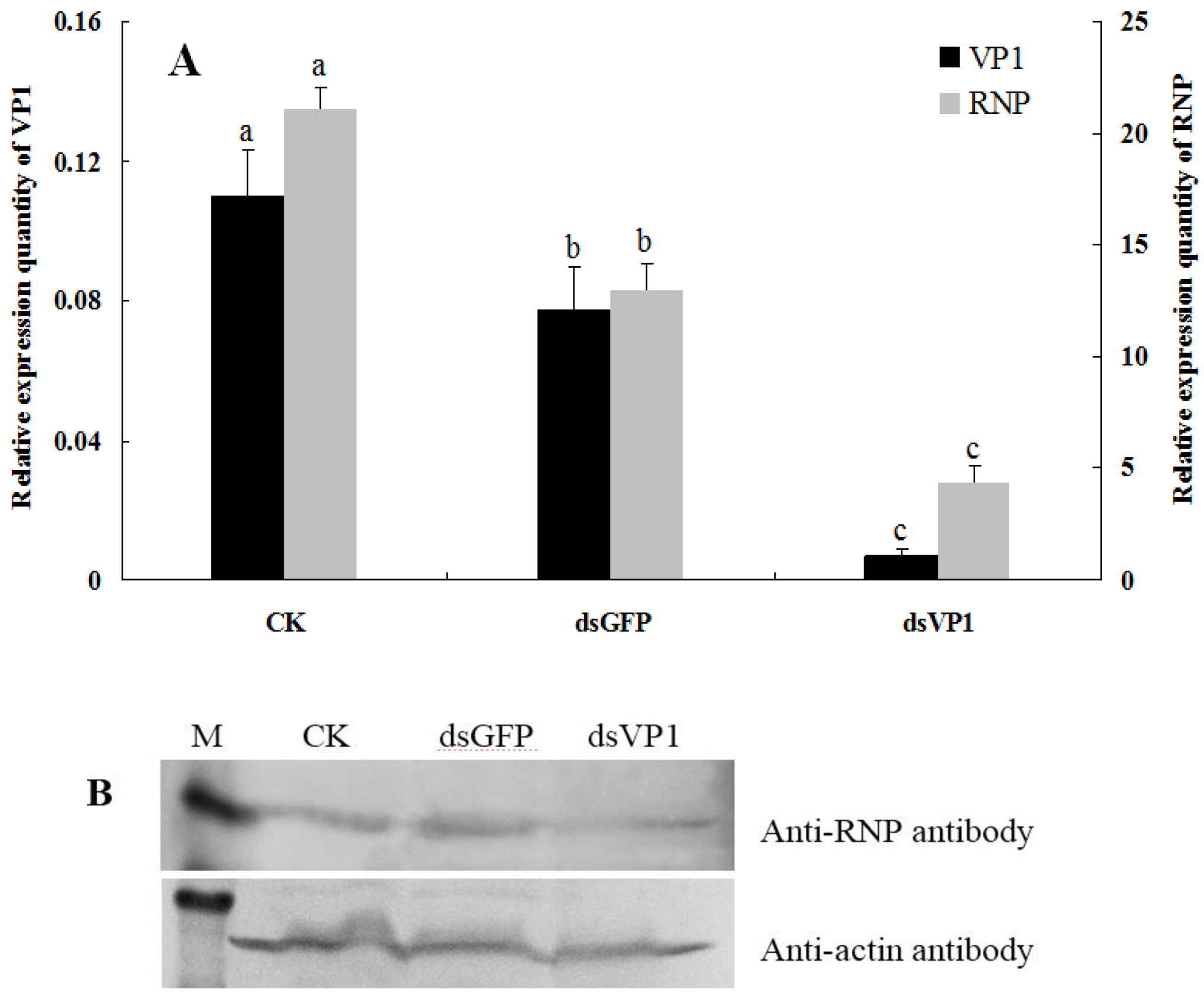
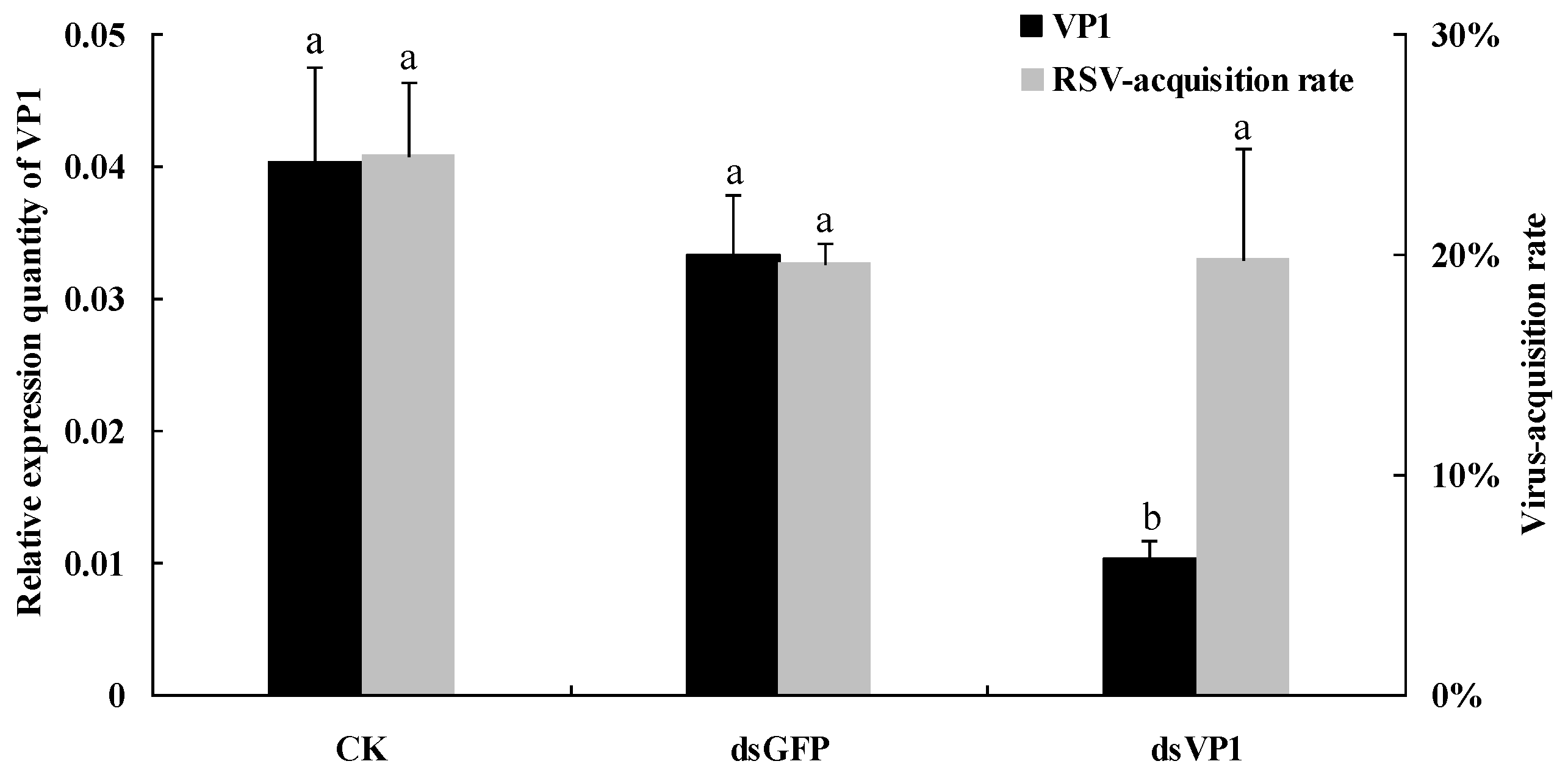
3.3. RSV Accumulation in SBPHs after Silencing of VP1

3.4. Virus-Acquisition Ability of SBPHs after RNAi

4. Discussion
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhou, Y.J.; Li, S.; Cheng, Z.B.; Zhou, T.; Fan, Y.J. Research advances in rice stripe disease in China. Jiangsu J. Agric. Sci. 2012, 28, 1007–1015. [Google Scholar]
- Falk, B.W.; Tsai, J.H. Biology and molecular biology of viruses in the genus Tenuivirus. Annu. Rev. Phytopathol. 1998, 36, 139–163. [Google Scholar] [CrossRef] [PubMed]
- Hibino, H. Biology and epidemiology of rice viruses. Annu. Rev. Phytopathol. 1996, 34, 249–274. [Google Scholar] [CrossRef] [PubMed]
- Otuka, A.; Zhou, Y.J.; Lee, G.S.; Matsumura, M.; Zhu, Y.Q.; Park, H.H.; Liu, Z.W.; Sanada-Morimura, S. Prediction of overseas migration of the small brown planthopper, Laodelphax striatellus (Hemiptera: Delphacidae) in East Asia. Appl. Entomol. Zool. 2012, 47, 379–388. [Google Scholar] [CrossRef]
- Guy, P.L.; Toriyama, S.; Fuji, S. Occurrence of a picorna-like virus in planthopper species and its transmission in Laodelphax striatellus. J. Invertebr. Pathol. 1992, 59, 161–164. [Google Scholar] [CrossRef]
- Toriyama, S.; Guy, P.L.; Fuji, S.; Takahashi, M. Characterization of a new picorna-like virus, himetobi P virus, in planthoppers. J. Gen. Virol. 1992, 73, 1021–1023. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Toriyama, S.; Matsuda, I.; Kojima, M. Detection of a picorna-like virus, himetobi P virus, in organs and tissues of Laodelphax striatellus by immunogold labeling and enzyme-linked immunosorbent assay. J. Invertebr. Pathol. 1993, 62, 99–104. [Google Scholar] [CrossRef]
- Nakashima, N.; Sasaki, J.; Toriyama, S. Determining the nucleotide sequence and capsid-coding region of himetobi P virus: A member of a novel group of RNA viruses that infect insects. Arch. Virol. 1999, 144, 2051–2058. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xiong, R.Y.; Wang, X.F.; Zhou, Y.J. Five proteins of Laodelphax striatellus are potentially involved in the interactions between rice stripe virus and vector. PLoS One 2011, 6, e26585. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.Z.; Zhou, Y.J.; Chen, Z.X.; Zhou, X.P. Production of monoclonal antibodies to rice stripe virus and application in virus detection. Acta Phytopathol. Sin. 2004, 34, 302–306. [Google Scholar]
- Li, S.; Sun, L.J.; Li, X.; Xiong, R.Y.; Xu, Q.F.; Zhou, Y.J. Construction of yeast two-hybrid cDNA library of high-viruliferous (RSV-infected) populations of the small brown planthopper, Laodelphax. striatellus (Hemiptera: Delphacidae). Acta Entomol. Sin. 2011, 54, 1324–1328. [Google Scholar]
- Li, S.; Li, X.; Sun, L.J.; Zhou, Y.J. Analysis of rice stripe virus whole-gene expression in rice and in the small brown planthopper by real-time quantitative PCR. Acta Virol. 2012, 56, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Zhang, Z.T.; Hu, C.; Lai, F.X.; Sun, Z.X. A chemically defined diet enables continuous rearing of the brown planthopper, Nilaparvata lugens (Stål) (Homoptera: Delphacidae). Appl. Entomol. Zool. 2001, 36, 111–116. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, D.; Yao, Q.; Zhang, J.; Dong, X.; Tian, H.; Chen, J.; Zhang, W. Feeding-based RNA interference of a trehalose phosphate synthase gene in the brown planthopper, Nilaparvata lugens. Insect Mol. Biol. 2010, 19, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, S.J.; Zi, J.Y.; Wang, X.; Zhou, Y.J. Rapid acquisition of rice stripe virus from infected leaves in vitro in small brown planthopper. Jiangsu J. Agric. Sci. 2014, 30, 449–451. [Google Scholar]
- Zhou, G.H.; Wen, J.J.; Cai, D.J.; Li, P.; Xu, D.L.; Zhang, S.G. Southern rice black-streaked dwarf virus: A new proposed Fijivirus species in the family Reoviridae. Chin. Sci. Bull. 2008, 53, 3677–3685. [Google Scholar] [CrossRef]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J. Insect Physiol. 2010, 56, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Araujo, R.N.; Santos, A.; Pinto, F.S.; Gontijo, N.F.; Lehane, M.J.; Pereira, M.H. RNA interference of the salivary gland nitrophorin 2 in the triatomine bug Rhodnius prolixus (Hemiptera: Reduviidae) by dsRNA ingestion or injection. Insect Biochem. Mol. Biol. 2006, 36, 683–693. [Google Scholar] [CrossRef]
- Turner, C.T.; Davy, M.W.; Macdiarmid, R.M.; Plummer, K.M.; Birch, N.P.; Newcomb, R.D. RNA interference in the light brown apple moth, Epiphyas postvittana (Walker) induced by double-stranded RNA feeding. Insect Mol. Biol. 2006, 15, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Walshe, D.P.; Lehane, S.M.; Lehane, M.J.; Haines, L.R. Prolonged gene knockdown in the tsetse fly Glossina. by feeding double stranded RNA. Insect Mol. Biol. 2009, 18, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Ammar, E.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, J.F.; Verbeek, M.; van der Wilk, F. Endosymbiotic bacteria associated with circulative transmission of potato leafroll virus by Myzus persicae. J. Gen. Virol. 1994, 75, 2559–2565. [Google Scholar] [CrossRef] [PubMed]
- Akad, F.; Dotan, N.; Czosnek, H. Trapping of tomato yellow leaf curl virus (TYLCV) and other plant viruses with a GroEL homologue from the whitefly Bemisia tabaci. Arch. Virol. 2004, 149, 1481–1497. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Kontsedalov, S.; Skaljac, M.; Brumin, M.; Sobol, I.; Czosnek, H.; Vavre, F.; Fleury, F.; et al. The transmission efficiency of tomato yellow leaf curl virus by the whitefly Bemisia tabaci is correlated with the presence of a specific symbiotic bacterium species. J. Virol. 2010, 84, 9310–9317. [Google Scholar] [CrossRef] [PubMed]
- Saridaki, A.; Bourtzis, K. Wolbachia: More than just a bug in insects genitals. Curr. Opin. Microbiol. 2010, 13, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia. symbiont in Aedes aegypti limits infection with dengue, Chikungunya and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef]
- Bian, G.; Xu, Y.; Lu, P.; Xie, Y.; Xi, Z.Y. The endosymbiotic bacterium Wolbachia. induces resistance to dengue virus in Aedes aegypti. PLoS Pathog. 2010, 6, e1000833. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.J.; Guo, H.Y.; Zheng, H.J.; Zhou, T.; Zhou, Y.J.; Wang, S.Y.; Fang, R.X.; Qian, W.; Chen, X.Y. Massively parallel pyrosequencing-based transcriptome analyses of small brown planthopper (Laodelphax striatellus), a vector insect transmitting rice stripe virus (RSV). BMC Genomics 2010, 11, 303. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Huang, L.Z.; Wang, Z.C.; Fu, S.; Che, J.; Qian, Y.J.; Zhou, X.P. Identification of Himetobi P virus in the small brown planthopper by deep sequencing and assembly of virus-derived small interfering RNAs. Virus Res. 2013, 179, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, V. Ribosome structure and the mechanism of translation. Cell 2002, 108, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Kaziro, Y. The role of guanosine 5'-triphosphate in polypeptide chain elongation. Biochim. Biophys. Acta 1978, 505, 95–127. [Google Scholar] [CrossRef] [PubMed]
- Horman, S.; Browne, G.J.; Krause, U.; Patel, J.V.; Vertommen, D.; Bertrand, L.; Lavoinne, A.; Hue, L.; Proud, C.G.; Rider, M.H. Activation of AMP-activated protein kinase leads to the phosphorylation of elongation factor 2 and an inhibition of protein synthesis. Curr. Biol. 2002, 12, 1419–1423. [Google Scholar] [CrossRef] [PubMed]
- Spahn, C.M.T.; Gomez-Lorenzo, M.G.; Grassucci, R.A.; Jørgensen, R.; Andersen, G.R.; Beckmann, R.; Penczek, P.A.; Ballesta, J.P.G.; Frank, J. Domain movements of elongation factor eEF2 and the eukaryotic 80S ribosome facilitate tRNA translocation. EMBO J. 2004, 23, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.; Liu, W.W.; Zhang, F.J.; Chen, X.Y.; Li, L.; Liu, Q.F.; Zhou, Y.J.; Wei, T.Y.; Fang, R.X.; Wang, X.F. Transovarial transmission of a plant virus is mediated by vitellogenin of its insect vector. PLoS Pathog. 2014, 10, e1003949. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.; Gildow, F.E. Luteovirus-aphid interactions. Annu. Rev. Phytopathol. 2003, 41, 539–566. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Pu, Y.Y.; Wei, T.Y.; Liu, H.J.; Deng, W.L.; Wei, C.H.; Ding, B.; Omura, T.; Li, Y. The P2 capsid protein of the nonenveloped rice dwarf phytoreovirus induces membrane fusion in insect host cells. Proc. Natl. Acad. Sci. USA 2007, 104, 19547–19552. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.Y.; Chen, H.Y.; Ichiki-Uehara, T.; Hibino, H.; Omura, T. Entry of rice dwarf virus into cultured cells of its insect vector involves clathrin-mediated endocytosis. J. Virol. 2007, 81, 7811–7815. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.Y.; Hibino, H.; Omura, T. Rice dwarf virus is engulfed into and released via vesicular compartments in cultured insect vector cells. J. Gen. Virol. 2008, 89, 2915–2920. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Ge, S.; Wang, X.; Sun, L.; Liu, Z.; Zhou, Y. Facilitation of Rice Stripe Virus Accumulation in the Insect Vector by Himetobi P Virus VP1. Viruses 2015, 7, 1492-1504. https://0-doi-org.brum.beds.ac.uk/10.3390/v7031492
Li S, Ge S, Wang X, Sun L, Liu Z, Zhou Y. Facilitation of Rice Stripe Virus Accumulation in the Insect Vector by Himetobi P Virus VP1. Viruses. 2015; 7(3):1492-1504. https://0-doi-org.brum.beds.ac.uk/10.3390/v7031492
Chicago/Turabian StyleLi, Shuo, Shangshu Ge, Xi Wang, Lijuan Sun, Zewen Liu, and Yijun Zhou. 2015. "Facilitation of Rice Stripe Virus Accumulation in the Insect Vector by Himetobi P Virus VP1" Viruses 7, no. 3: 1492-1504. https://0-doi-org.brum.beds.ac.uk/10.3390/v7031492