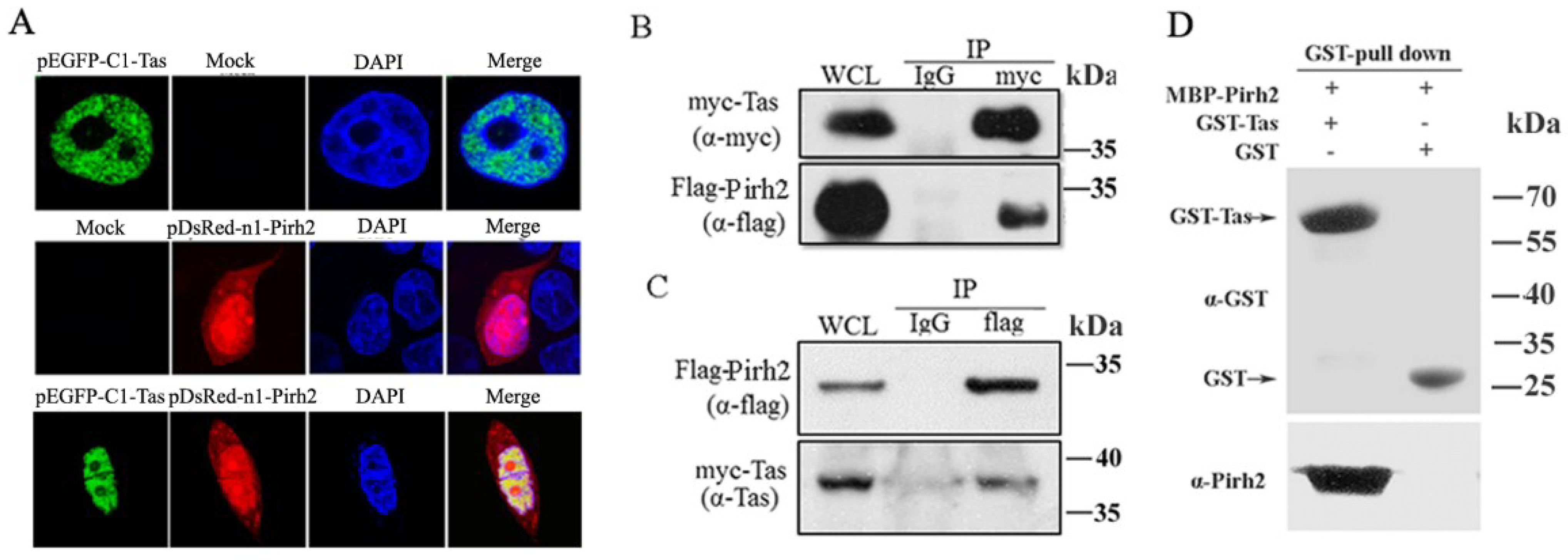
3.1. Tas Interacts with Pirh2
To screen proteins that may impact PFV replication through interaction with Tas, a human 293T cDNA library was screened in a yeast two-hybrid assay with Tas as the bait. We obtained 25 positive clones, and one of them contained a 786 bp cDNA fragment (GenBank accession No. GQ250944.1) encoding the full length protein Pirh2 and confirmed the interaction by yeast two-hybrid assay. To validate their interaction, we first detected the subcellular locations of Tas and Pirh2 by transfecting pEGFP-C1-Tas or pDsRed-N1-Pirh2 into 293T cells for 24 h, to investigate whether they would change the distribution by co-expressing with each other, 293T cells were also cotransfected with pEGFP-C1-Tas and pDsRed-N1-Pirh2 for 24 h, and examining the transfected cells with an Olympus confocal microscope. The distribution of Tas was almost completely nuclear, while, Pirh2 was distributed throughout both the nucleus and the cytoplasm, as reported previously [
29], and they colocalized predominantly in the nucleus and the co-expression did not change their distribution in the cells (
Figure 1A). To further confirm the interaction between Tas and Pirh2, coimmunoprecipitation was performed by co-transfecting 293T cells with myc-Tas and Flag-Pirh2. After 24 h, the cell lysate was divided into two equal portions and immunoprecipitated with anti-myc or with normal IgG as a control. Then, the precipitates were assessed by Western blot using anti-myc and anti-Flag antibodies. The data showed that Flag-Pirh2 could be coimmunoprecipitated with Tas (
Figure 1B). Tas was consistently coimmunoprecipitated with Flag-Pirh2 (
Figure 1C). In order to determine whether Tas binds directly to Pirh2, we expressed MBP-Pirh2 and GST-Tas in
E. coli (BL21) cells and purified the two proteins. Then we performed Glutathione S-transferase (GST) pull-down assays using purified GST-Tas and MBP-Pirh2. The results indicated Pirh2 was eluted together with GST-Tas but not GST, suggesting a direct association of Pirh2 and PFV Tas (
Figure 1D). Taken together, these results suggested that Pirh2 was able to interact with Tas.
Figure 1.
Tas interacts with Pirh2. (A) Tas colocalized with Pirh2 in nucleus. pEGFP-C1-Tas or/and pDsRed-N1-Pirh2 were transfected into 293T cells, 24 h after transfection, nuclei were visualized with DAPI staining; (B) Co-IP of Tas and Pirh2. 293T cells were cotransfected with myc-Tas and Flag-Pirh2, 24 h post transfection, the whole cell lysate was harvested and Tas was immunoprecipitated with anti-myc and normal IgG was used as negative control, then the immunoprecipitates were detected by Western blot using anti-myc and anti-Flag; (C) Reciprocal Co-IP of Tas and Pirh2. Immunoprecipitation (IP) was performed with anti-Flag, and analyzed by Western blot using anti- Flag and anti-Tas; (D) In vitro interaction between Tas and Pirh2. The purified Pirh2 protein was incubated with GST or GST-Tas agarose beads. GST pull down was then performed, and precipitates were detected by Western blot using an anti-Pirh2 antibody. Purified GST and GST-Tas used in the pull-down assay detected α-GST.
Figure 1.
Tas interacts with Pirh2. (A) Tas colocalized with Pirh2 in nucleus. pEGFP-C1-Tas or/and pDsRed-N1-Pirh2 were transfected into 293T cells, 24 h after transfection, nuclei were visualized with DAPI staining; (B) Co-IP of Tas and Pirh2. 293T cells were cotransfected with myc-Tas and Flag-Pirh2, 24 h post transfection, the whole cell lysate was harvested and Tas was immunoprecipitated with anti-myc and normal IgG was used as negative control, then the immunoprecipitates were detected by Western blot using anti-myc and anti-Flag; (C) Reciprocal Co-IP of Tas and Pirh2. Immunoprecipitation (IP) was performed with anti-Flag, and analyzed by Western blot using anti- Flag and anti-Tas; (D) In vitro interaction between Tas and Pirh2. The purified Pirh2 protein was incubated with GST or GST-Tas agarose beads. GST pull down was then performed, and precipitates were detected by Western blot using an anti-Pirh2 antibody. Purified GST and GST-Tas used in the pull-down assay detected α-GST.
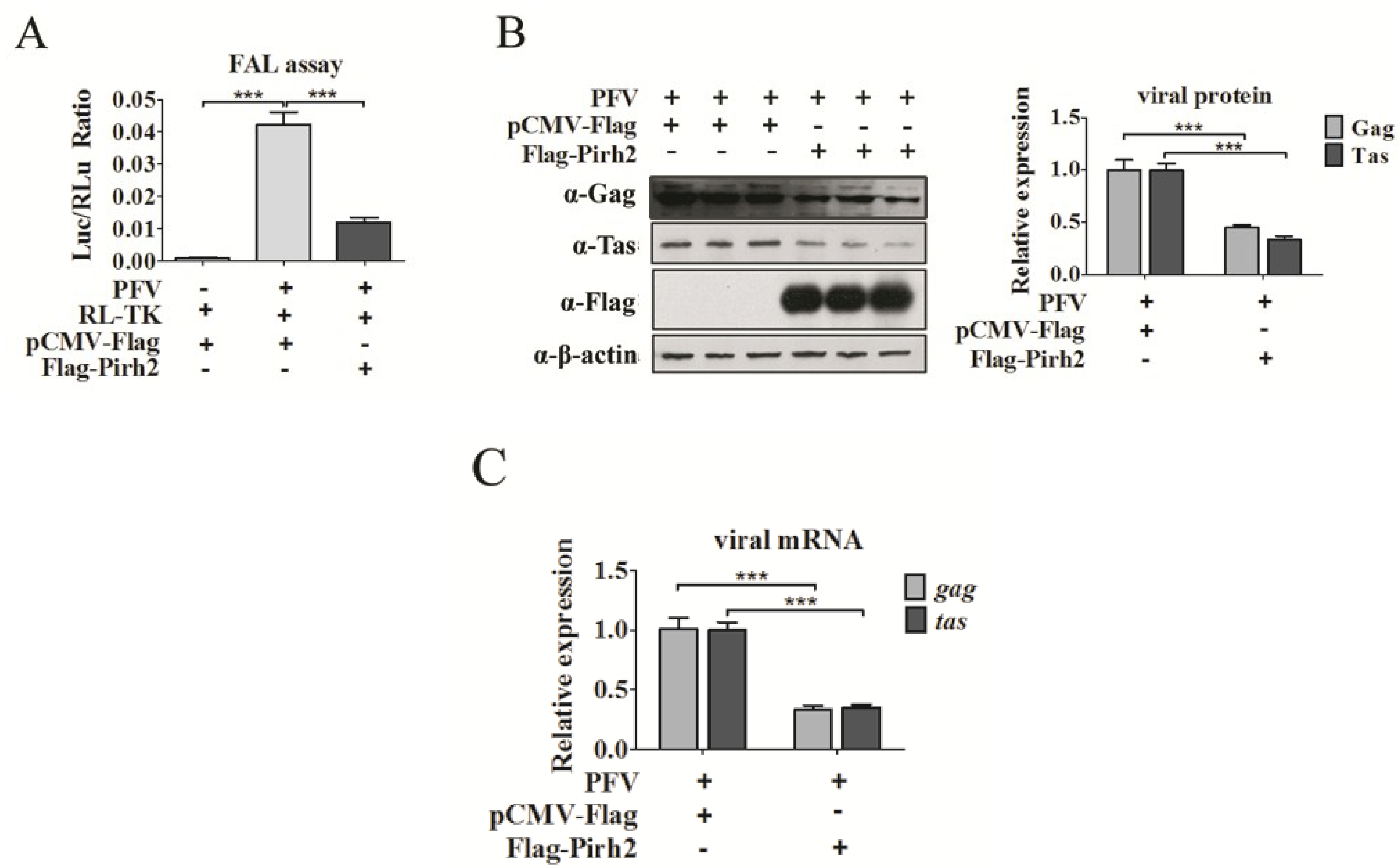
3.2. Pirh2 Inhibits PFV Replication
Because Pirh2 was able to interact with Tas, we evaluated whether Pirh2 was able to affect PFV replication. First, we determined whether the cellular PFV viral load would change with Pirh2 overexpression. For relative viral load detection, the PFV-infected 293T cells (1 × 10
4) were incubated with a PFV indicator cell line (PIC) (1 × 10
5) for 48 h [
19]. The PIC was established by stably transfecting baby hamster kidney-21 (BHK-21) cells with a luciferase gene driven by the PFV LTR and was more sensitive for quantifying PFV than TCID
50 [
23]. Twelve hours before incubating with the infected 293T cells, the PIC was transfected with RL-TK plasmid expressing Renilla (RLu) luciferase as an internal control. After 48 h, the luciferase activity was assessed. The FAL assay results (
Figure 2A) showed that PFV was able to more prominently activate the PIC compared to the control group, and the Luc/RLu ratio of Flag-Pirh2-transfected cells was significantly reduced compared to what was observed in pCMV-Flag-transfected cells (
Figure 2A), suggesting that overexpressed Pirh2 decreased virus production. Second, Western blot was performed to analyze the cellular viral proteins Gag and Tas in PFV-infected cells with or without Pirh2 overexpression. As shown in
Figure 2B, Pirh2 overexpression led to an evident reduction in the Gag and Tas proteins of over 60% compared to the control, demonstrating that Pirh2 overexpression inhibited PFV gene expression. Third, to investigate whether Pirh2 affects the viral transcription, RT-qPCR was performed to evaluate the relative mRNA levels of the PFV structural gene
gag and the regulatory gene
tas in PFV-infected 293T cells with or without Pirh2 overexpression. These results showed that with overexpression of Pirh2, the viral
gag and
tas mRNAs were markedly decreased compared to the control (
Figure 2C), suggesting that Pirh2 may have an influence on transcription during an early stage of viral replication.
To further confirm the inhibition of endogenous Pirh2 for PFV, we utilized a gene-specific siRNA to knockdown Pirh2 in 293T cells. 293T cells were transfected with either Pirh2-siRNA or NC (used as a negative control) for 24 h and challenged with PFV for another 48 h [
30]. The knockdown efficacy of endogenous Pirh2 protein by Pirh2-siRNA was confirmed by Western blot. As expected, siRNA down regulated Pirh2 protein level effectively and the loss of Pirh2 resulted in enhanced PFV replication as evidenced by facilitated production of the viral structural protein Gag (
Figure 2D) and the regulatory protein Tas (
Figure 2D), as assessed by the Western blot method. The mRNA of the viral genes
gag and
tas were consistently increased by over two-fold in the RT-PCR analysis when Pirh2 was knocked down (
Figure 2F). Collectively, our results suggest for the first time that host cell endogenous Pirh2 is a novel PFV replication inhibitor.
Figure 2.
Pirh2 inhibits the replication of PFV. (A) The relative viral load in the presence or absence of overexpressed Pirh2 was analyzed using the FAL assay with a PFV indicator cell line. RL-TK (5 μg) was transfected as an internal control; (B) 293T cells that seeded in 6-well plate and challenged with PFV (MOI 0.1) were transfected with Flag-Pirh2 (4 μg) or pCMV-Flag (4 μg) (as a control), the level of the viral protein Gag and Tas were analyzed by Western blot with β-actin used as a loading control. Quantitation of Gag and Tas protein levels from the Western blot by using Quantity one software (Bio-Rad); (C) 293T cells were seeded in 6-well plate and transfected with Flag-Pirh2 (4 μg) for 24 h (pCMV-Flag (4 μg) was used as a control) and then challenged with PFV for another 48 h. Then, the total RNA (2 μg) was reverse transcribed to cDNA. RT-qPCR was used to examine the relative expression (normalized to β-actin) of the viral structural gene gag and the regulatory gene tas with or without Pirh2 overexpression; (D) Specific siRNA used to knockdown Pirh2 (100 pmol) and NC (100 pmol) used as a negative control. The viral protein Gag and Tas was detected by Western blot, quantitation analysis of Gag and Tas intensity from the Western blot by using Quantity one software (Bio-Rad); (E) Relative mRNA expression (normalized to β-actin) of the viral structural gene gag and regulatory gene tas in cells that were transfected with Pirh2 siRNA or NC were assessed by RT-qPCR. All the data are representative of three independent experiments with triplicate samples. (Student’s t-test, ** p < 0.01; *** p < 0.001).
Figure 2.
Pirh2 inhibits the replication of PFV. (A) The relative viral load in the presence or absence of overexpressed Pirh2 was analyzed using the FAL assay with a PFV indicator cell line. RL-TK (5 μg) was transfected as an internal control; (B) 293T cells that seeded in 6-well plate and challenged with PFV (MOI 0.1) were transfected with Flag-Pirh2 (4 μg) or pCMV-Flag (4 μg) (as a control), the level of the viral protein Gag and Tas were analyzed by Western blot with β-actin used as a loading control. Quantitation of Gag and Tas protein levels from the Western blot by using Quantity one software (Bio-Rad); (C) 293T cells were seeded in 6-well plate and transfected with Flag-Pirh2 (4 μg) for 24 h (pCMV-Flag (4 μg) was used as a control) and then challenged with PFV for another 48 h. Then, the total RNA (2 μg) was reverse transcribed to cDNA. RT-qPCR was used to examine the relative expression (normalized to β-actin) of the viral structural gene gag and the regulatory gene tas with or without Pirh2 overexpression; (D) Specific siRNA used to knockdown Pirh2 (100 pmol) and NC (100 pmol) used as a negative control. The viral protein Gag and Tas was detected by Western blot, quantitation analysis of Gag and Tas intensity from the Western blot by using Quantity one software (Bio-Rad); (E) Relative mRNA expression (normalized to β-actin) of the viral structural gene gag and regulatory gene tas in cells that were transfected with Pirh2 siRNA or NC were assessed by RT-qPCR. All the data are representative of three independent experiments with triplicate samples. (Student’s t-test, ** p < 0.01; *** p < 0.001).
![Viruses 07 01668 g002a]()
![Viruses 07 01668 g002b]()
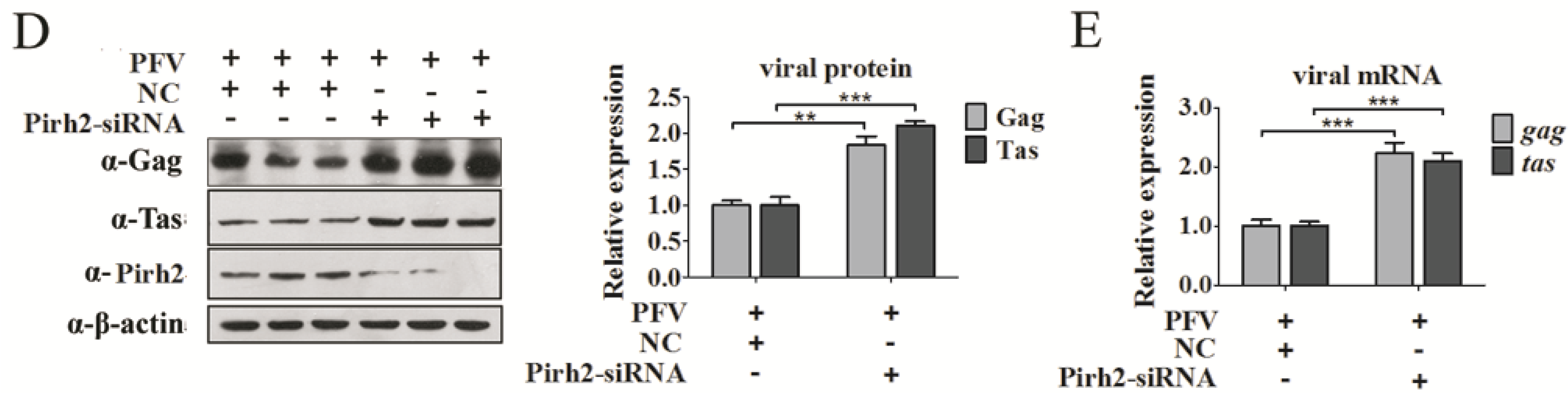
3.3. Pirh2 Negatively Regulates the Tas-Dependent Transcriptional Activation of PFV LTR and IP
Given that Pirh2 repressed PFV at both the protein and mRNA levels, this may imply that Pirh2 may function at the transcription stage or an early stage of the virus life cycle. Because many other factors that interact with Tas may influence the transactivity of Tas, we speculated that Pirh2 may suppress viral transcription. To investigate whether Pirh2 interfered with the basal or Tas-dependent transcriptional activation of both the conventional LTR promoter and IP, a luciferase reporter gene under the control of the PFV LTR (LTR-Luc) or the IP (IP-Luc) was utilized to assess the influence of Pirh2 on PFV gene transcription. We transfected 293T cells with PFV LTR-Luc or IP-Luc, with or without TK-Tas, together with Flag-Pirh2 plasmid or pCMV-Flag as a control, and RL-TK was used to normalize the transfection efficiency. Forty-eight hours post-transfection, Luc/RLu activity was measured. We observed that Tas dramatically activated theg LTR (
Figure 3A) and IP (
Figure 3B) as expected (
Figure 3A,B). Pirh2 did not impact the basal transcriptional activation of neither promoter (
Figure 3A,B). However, Pirh2 significantly impaired the Tas-dependent transcriptional activation of the LTR and IP. As shown in
Figure 3A,B, with overexpressed Pirh2, Tas transactivation of the LTR and IP was inhibited over 70% compared to the control. In contrast, Pirh2-specific siRNA transfection resulted in observably elevated Tas-dependent transcriptional activation of both the LTR (
Figure 3C) and the IP (
Figure 3D) compared to the control, the knockdown efficacy of Pirh2 protein by Pirh2-siRNA was confirmed by Western blot. These results indicated that Pirh2 could act as a repressor of Tas-dependent transactivation of the PFV LTR and IP. However, whether Pirh2 had negative effect on the PFV uncoating, entry or integration remained uncertain. Affecting one of those stages are sufficient to impact the viral protein and mRNA levels. Thus, we could not exclude that Pirh2 may influence the cellular uncoating-triggering signals, or Pirh2 may interfere the receptors on the cell membrane for PFV entry, and even affect the integration process. Further research is required.
Figure 3.
Pirh2 negatively regulates the Tas-dependent transcriptional activation of PFV LTR and IP. (A,B) Overexpressing Pirh2 down-regulates Tas-dependent transcription of the PFV LTR and IP. 293T cells were transfected with LTR-Luc (40 ng) (A) or IP-Luc (20 ng) (B) in the presence or absence TK-Tas (50 ng) combined with Flag-Pirh2 (400 ng) or pCMV-Flag (400 ng) (as a control); RL-TK was used to normalize transfection efficiency. At 48 h post transfection, luciferase activities were measured according to the manufacturer’s instructions; (C,D) Knockdown of Pirh2 with its specific siRNA up-regulates Tas-dependent transcription of the PFV LTR and IP. 293T cells were transfected with LTR-Luc (C) (40 ng) or IP-Luc (20 ng) (D) in the presence TK-Tas (50 ng) combined with Pirh2-siRNA (20 pmol) or nonsilencing siRNA NC (20 pmol) (as a negative control); RL-TK (50 ng) was used to normalize transfection efficiency. At 48 h post transfection, luciferase activities were measured according to the manufacturer’s instructions. (Student’s t-test, ** p < 0.01; *** p < 0.001).
Figure 3.
Pirh2 negatively regulates the Tas-dependent transcriptional activation of PFV LTR and IP. (A,B) Overexpressing Pirh2 down-regulates Tas-dependent transcription of the PFV LTR and IP. 293T cells were transfected with LTR-Luc (40 ng) (A) or IP-Luc (20 ng) (B) in the presence or absence TK-Tas (50 ng) combined with Flag-Pirh2 (400 ng) or pCMV-Flag (400 ng) (as a control); RL-TK was used to normalize transfection efficiency. At 48 h post transfection, luciferase activities were measured according to the manufacturer’s instructions; (C,D) Knockdown of Pirh2 with its specific siRNA up-regulates Tas-dependent transcription of the PFV LTR and IP. 293T cells were transfected with LTR-Luc (C) (40 ng) or IP-Luc (20 ng) (D) in the presence TK-Tas (50 ng) combined with Pirh2-siRNA (20 pmol) or nonsilencing siRNA NC (20 pmol) (as a negative control); RL-TK (50 ng) was used to normalize transfection efficiency. At 48 h post transfection, luciferase activities were measured according to the manufacturer’s instructions. (Student’s t-test, ** p < 0.01; *** p < 0.001).
![Viruses 07 01668 g003]()
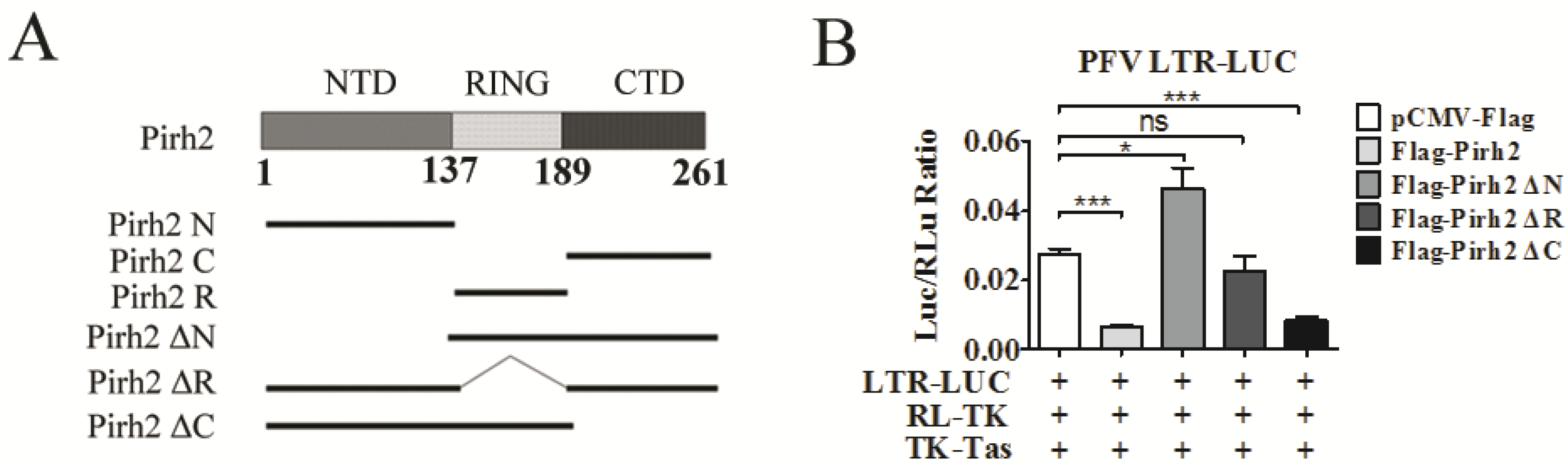
3.4. The Pirh2 N-Terminal Domain and RING Domain are Responsible for Its Inhibitory Effect on PFV Transcription and Replication
Pirh2 contains three domains, an N-terminal domain (NTD), a RING domain (RING), and a C-terminal domain (CTD), which fold independently of each other [
20]. To identify functional domains of Pirh2 that are essential for its inhibitory effect on PFV replication and transcription, we generated a series of Pirh2 expression constructs with various coding region truncations, including deletions of the NTD, RING, and CTD (
Figure 4A). The influences of each mutant Pirh2 on LTR transcriptional activation and PFV viral load were tested in a luciferase assay. We found that deletion of the NTD (Pirh2 ΔN) and RING (Pirh2 ΔR) showed no inhibitory effect on LTR transactivity (
Figure 4B), suggesting that the NTD plays an important role in the inhibitory effect of Pirh2. Interestingly, Pirh2 ΔN was able to enhance the LTR transcriptional activation by 1.5-fold (
Figure 4B). In addition, deletion of the CTD (Pirh2 ΔC) reduced LTR transcriptional activation by approximately 70%, which is similar to the complete Pirh2 (
Figure 4B), suggesting that the CTD may not be required for Pirh2 to suppress the LTR. Taken together, the NTD and/or RING of Pirh2 may be important for reducing LTR transcription and viral replication inhibition.
Subsequently, to confirm whether a single domain of Pirh2 was sufficient to repress PFV LTR transcription, Pirh2 N, Pirh2 R, and Pirh2 C were transfected into 293T cells with PFV LTR-Luc. These results indicated that both the NTD and the RING of Pirh2 are capable of decreasing LTR transcription by over 50% compared to the control (
Figure 4C). Because the RING domain of Pirh2 is associated with its E3 ubiquitin ligase activity, we speculated that ubiquitination may be involved in Pirh2-mediated inhibition. Interestingly, the Pirh2 CTD modestly promoted LTR transcription by 1.5-fold (
Figure 4C). Based on the previous results and the Pirh2 ΔN results (
Figure 4B), we hypothesized that the CTD may function as a transactivation domain.
Figure 4.
Identification of the key domain of Pirh2 that is responsible for the suppression of PFV. (A) Different domains of Pirh2 were cloned into the eukaryotic expression vector pCMV-3Flag; (B,C) The influences of Pirh2 ΔN, Pirh2 ΔR, Pirh2 ΔC (B) and Pirh2 N, Pirh2 R, Pirh2 C (C) on LTR transcription were determined by dual luciferase assay. 293T cells were transfected with LTR-Luc in the presence TK-Tas combined with different truncated Pirh2 plasmids. RL-TK was used to normalize transfection efficiency. At 48 h post transfection, luciferase activities were measured; (D) The relative viral load in the presence of different Pirh2 truncations was analyzed using the FAL assay with a PFV indicator cell line. All the data are representative of three independent experiments with triplicate samples. Asterisks denote significant differences of samples compared with the pCMV-Flag transfected group. (Student’s t-test, * p < 0.05; ** p < 0.01; *** p < 0.001, ns = not significant).
Figure 4.
Identification of the key domain of Pirh2 that is responsible for the suppression of PFV. (A) Different domains of Pirh2 were cloned into the eukaryotic expression vector pCMV-3Flag; (B,C) The influences of Pirh2 ΔN, Pirh2 ΔR, Pirh2 ΔC (B) and Pirh2 N, Pirh2 R, Pirh2 C (C) on LTR transcription were determined by dual luciferase assay. 293T cells were transfected with LTR-Luc in the presence TK-Tas combined with different truncated Pirh2 plasmids. RL-TK was used to normalize transfection efficiency. At 48 h post transfection, luciferase activities were measured; (D) The relative viral load in the presence of different Pirh2 truncations was analyzed using the FAL assay with a PFV indicator cell line. All the data are representative of three independent experiments with triplicate samples. Asterisks denote significant differences of samples compared with the pCMV-Flag transfected group. (Student’s t-test, * p < 0.05; ** p < 0.01; *** p < 0.001, ns = not significant).
Furthermore, the FAL assay showed similar effects of different Pirh2 mutants on PFV replication (
Figure 4D). Deletion of the NTD or RING of Pirh2 did not inhibit PFV replication, and separately, the NTD and RING were capable of impairing PFV replication. In contrast, Pirh2ΔN and Pirh2C, which both contain the CTD, modestly facilitated PFV replication.
3.5. The E3 Ubiquitin Ligase Activity of Pirh2 Contributes to Its Inhibitory Effect on PFV
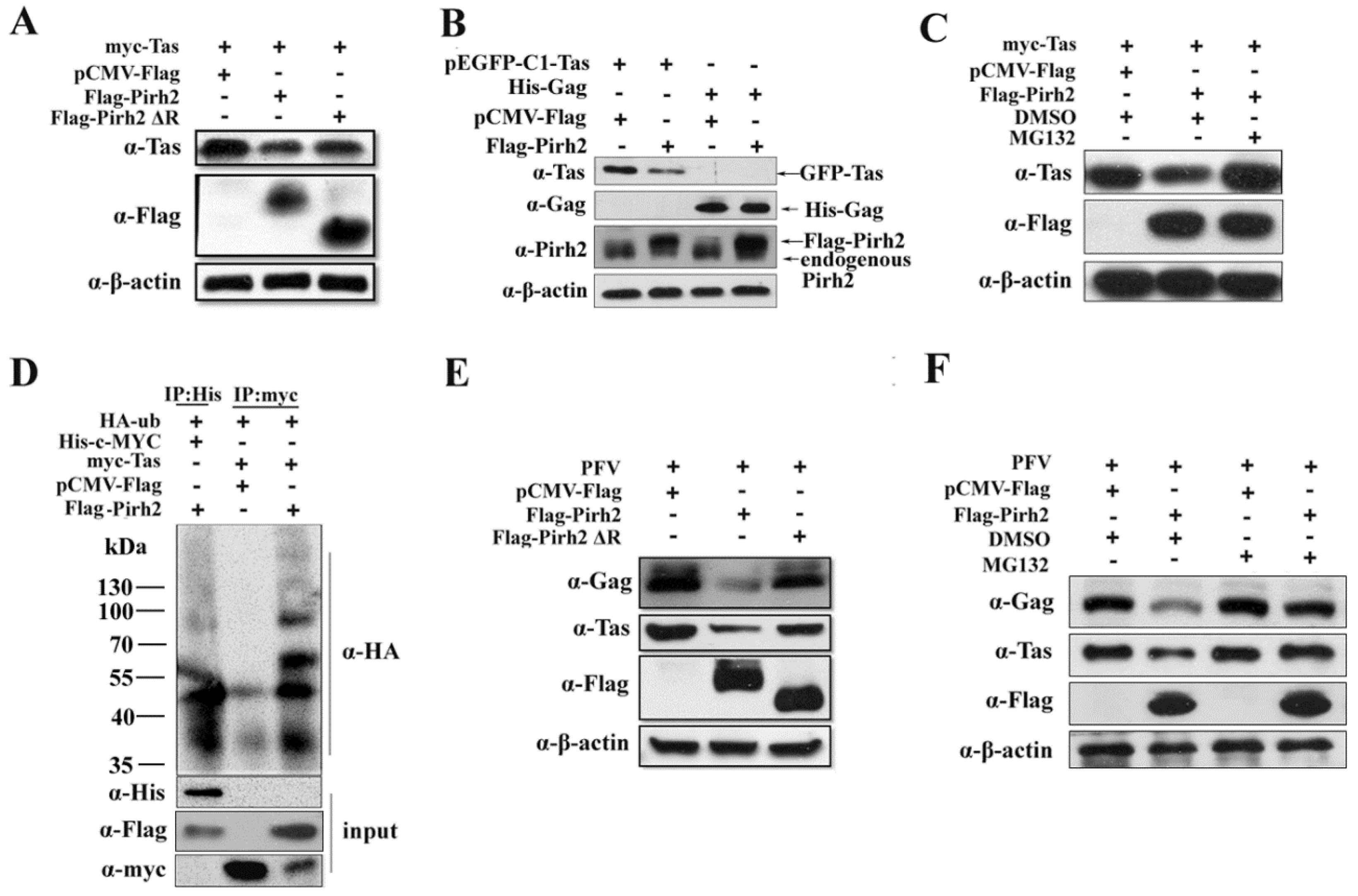
Because the RING domain of Pirh2 is related to the ubiquitin-proteasome, we next investigated whether Pirh2 could influence the stability of the Tas protein. myc-Tas was co-transfected with Flag-Pirh2 or Flag-Pirh2ΔR, and pCMV-Flag was used as control. As shown in
Figure 5A, overexpression of Pirh2 led to a decline in the level of Tas protein compared to the control. To demonstrate that Pirh2 overexpression are specific to down regulated Tas. 293T cells were transfected Flag-Pirh2 with pEGFP-C1-Tas or His-Gag, and Western blot was used to detect the expression of Tas or Gag. The result showed that overexpressed Pirh2 could down regulate pEGFP-C1-Tas but not the His-Gag (
Figure 5B). And Pirh2 lacking the RING domain had little effect on Tas expression, suggesting that the inhibitory effect of Pirh2 on PFV is partially attributable to its E3 ubiquitin ligase activity. To further confirm whether the molecular mechanism is related to the proteasome degradation pathway, Pirh2-overexpressing cells were treated with the proteasome inhibitor MG132 at 5 μM for 8 h before lysis, and we found that Tas expression was notably rescued by MG132 (
Figure 5C), indicating that Pirh2 may decrease Tas through the ubiquitin-proteasome pathway. Then, we performed an
in vivo ubiquitination assay. The 293T cells were transfected with HA-tagged ubiquitin (HA-Ub), myc-Tas or c-MYC combined with Pirh2, pCMV-Flag used as a negative control, or c-MYC used as a positive control [
28].
Immunoprecipitation was used with anti-myc or anti-His antibodies, and the precipitates were analyzed with anti-HA antibody. Overexpression of Pirh2 was found to significantly increased myc-Tas polyubiquitination, which was similar to c-MYC, indicating that Pirh2 polyubiquitinated Tas and mediated its proteasomal degradation (
Figure 5D). We further tested by Western blot whether deletion of the RING domain or treatment with MG132 would reverse the inhibitory effect of Pirh2 on PFV replication. As expected, the PFV structural protein Gag and the regulatory protein Tas were slightly decreased by Pirh2ΔR compared to Pirh2 (
Figure 5E), and MG132 was able to counteract the inhibitory effect of overexpressed Pirh2 (
Figure 5F), but could not increase the Tas and Gag levels by itself. All of these results indicate that Pirh2 negatively influences viral replication by promoting the instability of Tas through the ubiquitin pathway.
Figure 5.
Pirh2 decreases the expression of Tas protein and inhibits PFV replication through ubiquitin-proteasome pathway. (A,E) Tas expression or PFV replication are down regulated by overexpressed Pirh2 and deletion of RING domain abrogates its inhibitory effect; (B) Pirh2 down regulates Tas specifically. 293T cells were transfected Flag-Pirh2 with pEGFP-C1-Tas or His-Gag, and Western blot was used to detect the expression of Tas or Gag; (C) Pirh2 down regulates the expression of Tas protein and the inhibition was reversed by MG132. myc-Tas combined with Flag-Pirh2 or pCMV-Flag (as control) were transfected into 293T cells; 8 h before harvest, the cells were treated with DMSO or MG132 (5 μM). Western blot was performed to assess Tas protein levels; (D) Pirh2 promotes Tas polyubiquitination. HA-tagged ubiquitin (HA-Ub), myc-Tas or c-MYC combined with Pirh2 were transfected into 293T cells, pCMV-Flag used as negative control and c-MYC used as positive control; (F) MG132 rescues the suppression of PFV produced by Pirh2. 293T cells were transfected with Flag-Pirh2 or pCMV-Flag (as control) for 24 h and challenged with PFV for another 48 h; 8 h before harvest, the cells transfected with Flag-Pirh2 or not were treated with DMSO or MG132 (5 μM). Then, Western blot was performed to assess the viral protein Gag and Tas.
Figure 5.
Pirh2 decreases the expression of Tas protein and inhibits PFV replication through ubiquitin-proteasome pathway. (A,E) Tas expression or PFV replication are down regulated by overexpressed Pirh2 and deletion of RING domain abrogates its inhibitory effect; (B) Pirh2 down regulates Tas specifically. 293T cells were transfected Flag-Pirh2 with pEGFP-C1-Tas or His-Gag, and Western blot was used to detect the expression of Tas or Gag; (C) Pirh2 down regulates the expression of Tas protein and the inhibition was reversed by MG132. myc-Tas combined with Flag-Pirh2 or pCMV-Flag (as control) were transfected into 293T cells; 8 h before harvest, the cells were treated with DMSO or MG132 (5 μM). Western blot was performed to assess Tas protein levels; (D) Pirh2 promotes Tas polyubiquitination. HA-tagged ubiquitin (HA-Ub), myc-Tas or c-MYC combined with Pirh2 were transfected into 293T cells, pCMV-Flag used as negative control and c-MYC used as positive control; (F) MG132 rescues the suppression of PFV produced by Pirh2. 293T cells were transfected with Flag-Pirh2 or pCMV-Flag (as control) for 24 h and challenged with PFV for another 48 h; 8 h before harvest, the cells transfected with Flag-Pirh2 or not were treated with DMSO or MG132 (5 μM). Then, Western blot was performed to assess the viral protein Gag and Tas.
![Viruses 07 01668 g005]()


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






