
Overview on Sobemoviruses and a Proposal for the Creation of the Family Sobemoviridae

Abstract
:1. Introduction

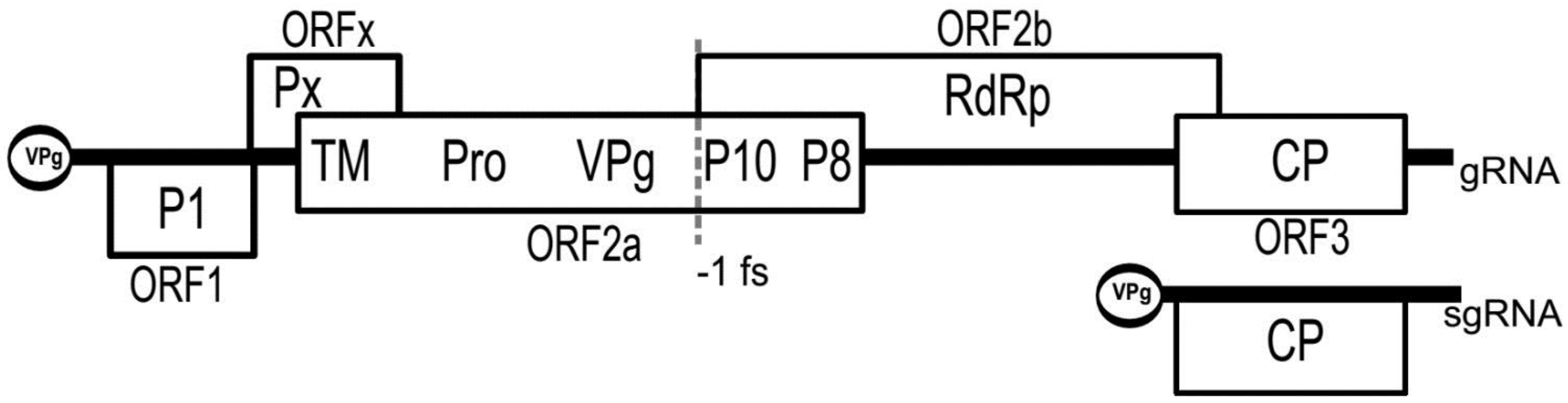{kind=link}
{kind=link}
{kind=link}
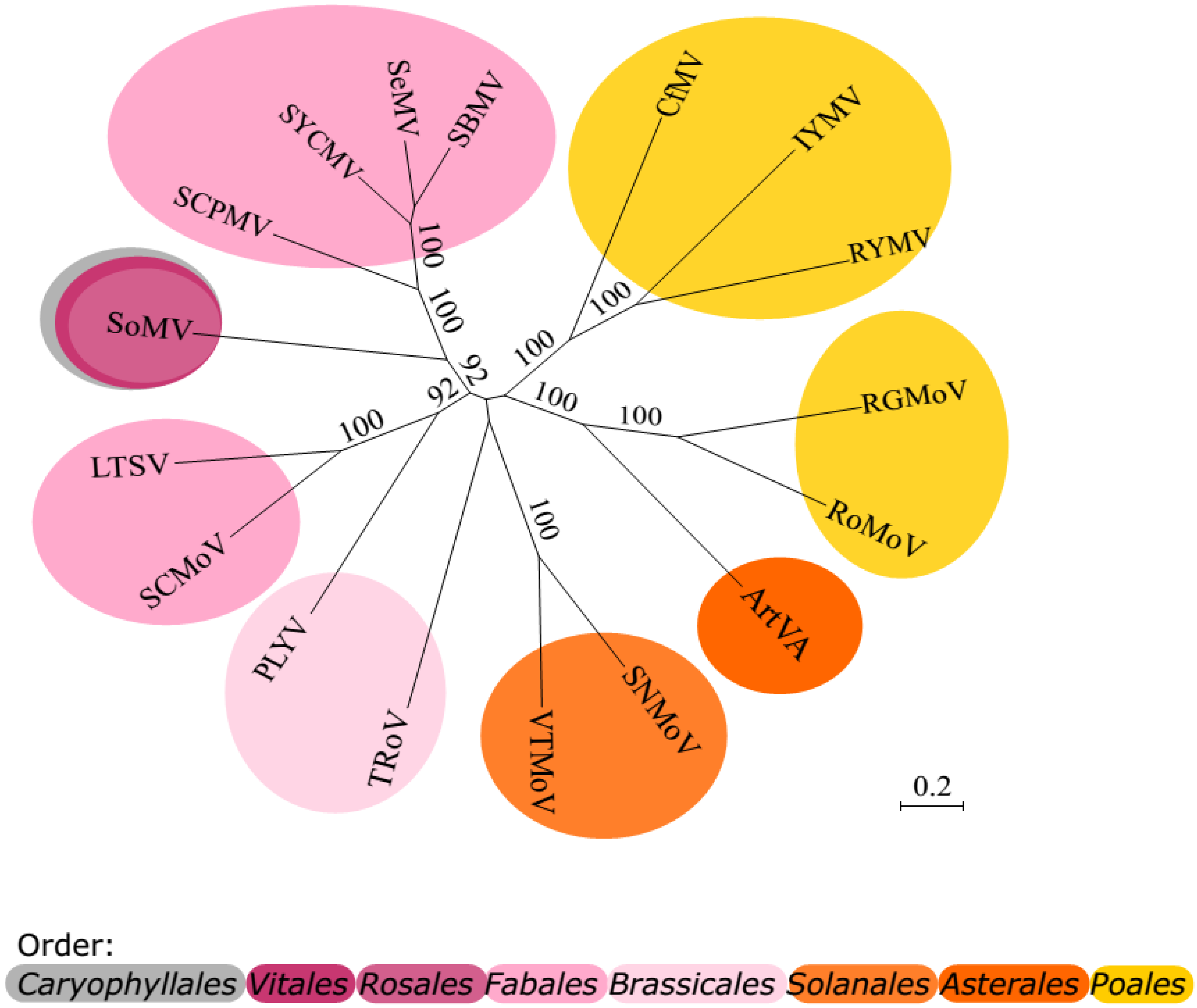{kind=link}
{kind=link}
| Species Recognized by ICTV | Abbreviation | Genome Sequence in GenBank (Acc. No) |
|---|---|---|
| Blueberry shoestring virus | BSSV | - |
| Cocksfoot mottle virus | CfMV | NC_002618 |
| Imperata yellow mottle virus | IYMV | NC_011536 |
| Lucerne transient streak virus | LTSV | NC_001696 |
| Rice yellow mottle virus | RYMV | NC_001575 |
| Ryegrass mottle virus | RGMoV | NC_003747 |
| Sesbania mosaic virus | SeMV | NC_002568 |
| Solanum nodiflorum mottle virus | SNMoV | KC577470 |
| Southern bean mosaic virus | SBMV | NC_004060 |
| Southern cowpea mosaic virus | SCPMV | NC_001625 |
| Sowbane mosaic virus | SoMV | NC_011187 |
| Subterranean clover mottle virus | SCMoV | NC_004346 |
| Turnip rosette virus | TRoV | NC_004553 |
| Velvet tobacco mottle virus | VTMoV | NC_014509 |
| Related Viruses not Approved by ICTV as Sobemovirus Species | ||
| Artemisia virus A | ArtVA | NC_017914 |
| Cynosurus mottle virus | CnMoV | - |
| Ginger chlorotic fleck virus | GCFV | - |
| Papaya lethal yellowing virus | PLYV | NC_018449 |
| Rottboellia yellow mottle virus | RoMoV | KC577469 |
| Snake melon asteroid mosaic virus | SMAMV | - |
| Soybean yellow common mosaic virus | SYCMV | NC_016033 |
2. Geographical Distribution, Host Range, Transmission
3. Genomic Organization

4. Satellite RNAs
5. P1 in Viral Movement and Suppression of RNA Silencing

6. Proteolytic Processing of Polyprotein
7. The Proteins VPg, P8, and P10
8. RNA-Dependent RNA Polymerase
9. Virion Topology

10. Virion Assembly and Disassembly
11. Localization of Virions in Cells and Tissues
12. Pathology and Resistance
13. Phylogenesis

14. A Proposal for the Creation of the Family Sobemoviridae

15. Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Walters, H.J. Beetle transmission of plant viruses. Adv. Virus Res. 1969, 15, 339–363. [Google Scholar] [PubMed]
- Hull, R. The grouping of small spherical plant viruses with single RNA components. J. Gen. Virol. 1977, 36, 289–295. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses. Virus Taxonomy: 2014 Release. Available online: http://www.ictvonline.org/virusTaxonomy.asp (accessed on 18 May 2015).
- Gratsia, M.E.; Kyriakopoulou, P.E.; Voloudakis, A.E.; Fasseas, C.; Tzanetakis, I.E. First report of Olive mild mosaic virus and Sowbane mosaic virus in spinach in Greece. Plant Dis. 2012, 96, 1230–1230. [Google Scholar] [CrossRef]
- Verhoeven, J.T.J.; Roenhorst, J.W.; Lesemann, D.E.; Segundo, E.; Velasco, L.; Ruiz, L.; Janssen, D.; Cuadrado, I.M. Southern bean mosaic virus the causal agent of a new disease of Phaseolus vulgaris beans in Spain. Eur. J. Plant Pathol. 2003, 109, 935–941. [Google Scholar] [CrossRef]
- Jones, R.A.C. Using epidemiological information to develop effective integrated virus disease management strategies. Virus Res. 2004, 100, 5–30. [Google Scholar] [CrossRef] [PubMed]
- Klimaszewski, J.; Watt, J.C. Coleoptera: Family-group review and keys to identification. Fauna N. Z. 1997, 37, 199. [Google Scholar]
- Campell, A.W.; Guy, P.L. Cocksfoot mottle virus spreads to the South Island of New Zealand. Australas. Plant Pathol. 2001, 30, 217–220. [Google Scholar] [CrossRef]
- Delmiglio, C.; Pearson, M.N.; Lister, R.A.; Guy, P.L. Incidence of cereal and pasture viruses in New Zealand’s native grasses. Ann. Appl. Biol. 2010, 157, 25–36. [Google Scholar] [CrossRef]
- Lima, R.C.A.; Lima, J.A.A.; Souza, M.T.; Pio-Ribeiro, G.; Andrade, G.P. Etiologia e estratégias de controle de viroses do mamoeiro no Brasil. Fitopatol. Bras. 2001, 26, 689–702. [Google Scholar] [CrossRef]
- Nascimento, A.K.Q.; Lima, J.A.A.; Nascimento, A.L.L.; Beserra, E.A.; Purcifull, D.E. Biological, physical, and molecular properties of a papaya lethal yellowing virus isolate. Plant Dis. 2010, 94, 1206–1212. [Google Scholar] [CrossRef]
- Traore, O.; Pinel-Galzi, A.; Sorho, F.; Sarra, S.; Rakotomalala, M.; Sangu, E.; Kanyeka, Z.; Sere, Y.; Konate, G.; Fargette, D. A reassessment of the epidemiology of Rice yellow mottle virus following recent advances in field and molecular studies. Virus Res. 2009, 141, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Ochola, D.; Tusiime, G. Survey on incidences and severity of rice yellow mottle virus disease in eastern Uganda. Int. J. Plant Pathol. 2011, 2, 15–25. [Google Scholar] [CrossRef]
- Ochola, D.; Issaka, S.; Rakotomalala, M.; Pinel-Galzi, A.; Ndikumana, I.; Hubert, J.; Hebrard, E.; Sere, Y.; Tusiime, G.; Fargette, D. Emergence of Rice yellow mottle virus in eastern Uganda: Recent and singular interplay between strains in East Africa and in Madagascar. Virus Res. 2015, 195, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Pinel-Galzi, A.; Traore, O.; Sere, Y.; Hebrard, E.; Fargette, D. The biogeography of viral emergence: Rice yellow mottle virus as a case study. Curr. Opin. Virol. 2015, 10, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Walters, H.J.; Henry, D.G. Bean leaf beetle as a vector of the cowpea strain of Southern bean mosaic virus. Phytopathology 1970, 60, 177–178. [Google Scholar] [CrossRef]
- Hollings, M.; Stone, O. Turnip rosette virus. In CMI/AAB Description of Plant Viruses No. 125; Commonwealth Mycological Institute, Kew, Surrey, and Association of Applied Biologists: Wellesbourne, Warwick, UK, 1973. [Google Scholar]
- Bakker, W. Characterization and Ecological Aspects of Rice yellow Mottle Virus in Kenya; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1974; p. 152. [Google Scholar]
- Catherall, P.L.; Andrews, E.A.; Chamberlain, J.A. Host-ranges of cocksfoot mottle and cynosurus mottle viruses. Ann. Appl. Biol. 1977, 87, 233–235. [Google Scholar] [CrossRef]
- Greber, R.S. Some characteristics of Solanum nodiflorum mottle virus—A beetle-transmitted isometric virus from Australia. Aust. J. Biol. Sci. 1981, 34, 369–378. [Google Scholar]
- Mohamed, N.A.; Mossop, D.W. Cynosurus and cocksfoot mottle viruses: A comparison. J. Gen. Virol. 1981, 55, 63–74. [Google Scholar] [CrossRef]
- Tremaine, J.H.; Hamilton, R.I. Southern bean mosaic virus. In CMI/AAB Description of Plant Viruses No. 274; Commonwealth Mycological Institute, Kew, Surrey, and Assocaiation of Applied Biologists: Wellesbourne, Warwick, UK, 1983. [Google Scholar]
- Bennet, C.W.; Costa, A.S. Sowbane mosaic caused by a seed-transmitted virus. Phytopathology 1961, 51, 546–550. [Google Scholar]
- Randles, J.W.; Davies, C.; Hatta, T.; Gould, A.R.; Francki, R.I.B. Studies on encapsidated viroid-like RNA. 1. Characterization of Velvet tobacco mottle virus. Virology 1981, 108, 111–122. [Google Scholar] [CrossRef]
- Gibb, K.S.; Randles, J.W. Transmission of Velvet tobacco mottle virus and related viruses by the mirid Cyrtopeltis nicotianae. Adv. Dis. Vector Res. 1991, 7, 1–17. [Google Scholar]
- Patino, Y.; Garrido, M.J.; Savini, V. Diaphaulaca aulica: New vector for bean southern mosaic sobemovirus. Bol. Entomol. Venez. 1997, 12, 121–122. [Google Scholar]
- Hardy, V.G.; Teakle, D.S. Transmission of Sowbane mosaic virus by thrips-tabaci in the presence and absence of virus-carrying pollen. Ann. Appl. Biol. 1992, 121, 315–320. [Google Scholar] [CrossRef]
- Mohamed, N.A. Cynosurus mottle virus a virus affecting grasses in New-Zealand. New Zeal. J. Agric. Res. 1978, 21, 709–714. [Google Scholar] [CrossRef]
- Morimoto, K.M.; Ramsdell, D.C.; Gillett, J.M.; Chaney, W.G. Acquisition and transmission of Blueberry shoestring virus by its aphid vector Illinoia pepperi. Phytopathology 1985, 75, 709–712. [Google Scholar] [CrossRef]
- Nwilene, F.E.; Traore, A.K.; Asidi, A.N.; Sere, Y.; Onasanya, A.; Abo, M.E. New records of insect vectors of Rice yellow mottle virus (RYMV) in Côte d’Ivoire, West Africa. J. Entomol. 2009, 6, 198–206. [Google Scholar] [CrossRef]
- Zaumeyer, W.J.; Harter, L.L. Inheritance of symptom expression of bean mosaic virus 4. J. Agric. Res. 1943, 67, 295–300. [Google Scholar]
- Shepherd, R.J.; Fulton, R.W. Identity of a seed-born virus of cowpea. Phytopathology 1962, 52, 489–493. [Google Scholar]
- Francki, R.I.B.; Randles, J.W.; Graddon, D.J. Subterranean clover mottle virus. In CMI/AAB Description of Plant Viruses No. 329; Commonwealth Mycological Institute, Kew, Surrey, and Assocaiation of Applied Biologists: Wellesbourne, Warwick, UK, 1988. [Google Scholar]
- Lecoq, H.; Fabre, F.; Joannon, B.; Wipf-Scheibel, C.; Chandeysson, C.; Schoeny, A.; Desbiez, C. Search for factors involved in the rapid shift in Watermelon mosaic virus (WMV) populations in south-eastern France. Virus Res. 2011, 159, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Uyemoto, J.K.; Grogan, R.G. Southern bean mosaic virus: Evidence for seed transmission in bean embryos. Phytopathology 1977, 67, 1190–1196. [Google Scholar] [CrossRef]
- Bos, L.; Huijberts, N. Occurrence and transmission of Sowbane mosaic virus in seed from naturally infected plants of spinach (Spinacia oleracea). Eur. J. Plant Pathol. 1996, 102, 707–711. [Google Scholar] [CrossRef]
- Hull, R. The stabilization of the particles of Turnip rosette virus and of other members of the southern bean mosaic virus group. Virology 1977, 79, 58–66. [Google Scholar] [CrossRef]
- Traore, O.; Pinel, A.; Hebrard, E.; Gumedzoe, M.Y.D.; Fargette, D.; Traore, A.S.; Konate, G. Occurrence of resistance-breaking isolates of Rice yellow mottle virus in west and central Africa. Plant Dis. 2006, 90, 259–263. [Google Scholar] [CrossRef]
- Sarra, S.; Peters, D. Rice yellow mottle virus is transmitted by cows, donkeys, and grass rats in irrigated rice crops. Plant Dis. 2003, 87, 804–808. [Google Scholar] [CrossRef]
- Peters, D.; Engels, C.; Sarra, S. Natural spread of plant viruses by birds. J. Phytopathol. 2012, 160, 591–594. [Google Scholar] [CrossRef]
- Sarra, S.; Oevering, P.; Guindo, S.; Peters, D. Wind-mediated spread of rice yellow mottle virus (RYMV) in irrigated rice crops. Plant Pathol. 2004, 53, 148–153. [Google Scholar] [CrossRef]
- Traore, M.D.; Traore, V.S.; Galzi-Pinel, A.; Fargette, D.; Konate, G.; Traore, A.S.; Traore, O. Abiotic transmission of Rice yellow mottle virus through soil and contact between plants. Pak. J. Biol. Sci. 2008, 11, 900–904. [Google Scholar] [CrossRef] [PubMed]
- Camarço, R.F.E.A.; Lima, J.A.A.; PioRibeiro, G. Transmissão e presença em solo do “papaya lethal yellowing virus”. Fitopatol. bras. 1998, 23, 453–458. [Google Scholar]
- Teakle, D.S.; Morris, T.J. Transmission of Southern bean mosaic virus from soil to bean-seeds. Plant Dis. 1981, 65, 599–600. [Google Scholar] [CrossRef]
- Ramel, M.E.; Gugerli, P.; Besse, S.; Simonnet, X.; Balmelli, C. Molecular characterization of artemisia virus A, a new sobemovirus isolated from Artemisia annua. Arch. Virol. 2013, 158, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Teakle, D.S. Abiotic transmission of Southern bean mosaic virus in soil. Aust. J. Biol. Sci. 1986, 39, 353–359. [Google Scholar]
- Hacker, D.L.; Sivakumaran, K. Mapping and expression of southern bean mosaic virus genomic and subgenomic RNAs. Virology 1997, 234, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, G.I.; Njeru, R.; Williamson, S.; Fosu-Nyarko, J.; Hopkins, R.; Jones, R.A.; Waterhouse, P.M.; Jones, M.G. The complete nucleotide sequence of Subterranean clover mottle virus. Arch. Virol. 2003, 148, 2237–2247. [Google Scholar] [CrossRef] [PubMed]
- Sivakumaran, K.; Hacker, D.L. The 105-kDa polyprotein of Southern bean mosaic virus is translated by scanning ribosomes. Virology 1998, 246, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Ling, R.; Pate, A.E.; Carr, J.P.; Firth, A.E. An essential fifth coding ORF in the sobemoviruses. Virology 2013, 446, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Meier, M.; Truve, E. Sobemoviruses possess a common CfMV-like genomic organization. Arch. Virol. 2007, 152, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Lucchesi, J.; Mäkeläinen, K.; Merits, A.; Tamm, T.; Mäkinen, K. Regulation of -1 ribosomal frameshifting directed by cocksfoot mottle sobemovirus genome. Eur. J. Biochem. 2000, 267, 3523–3529. [Google Scholar] [CrossRef] [PubMed]
- Mäkeläinen, K.; Mäkinen, K. Factors affecting translation at the programmed -1 ribosomal frameshifting site of cocksfoot mottle virus RNA in vivo. Nucl. Acids Res. 2005, 33, 2239–2247. [Google Scholar] [CrossRef] [PubMed]
- Tamm, T.; Suurväli, J.; Lucchesi, J.; Olspert, A.; Truve, E. Stem-loop structure of cocksfoot mottle virus RNA is indispensable for programmed -1 ribosomal frameshifting. Virus Res. 2009, 146, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Brisco, M.J.; Hull, R.; Wilson, T.M. Southern bean mosaic virus-specific proteins are synthesized in an in vitro system supplemented with intact, treated virions. Virology 1985, 143, 392–398. [Google Scholar] [CrossRef]
- Shields, S.A.; Brisco, M.J.; Wilson, T.M.; Hull, R. Southern bean mosaic virus RNA remains associated with swollen virions during translation in wheat germ cell-free extracts. Virology 1989, 171, 602–606. [Google Scholar] [CrossRef]
- Truve, E.; Fargette, D. Genus Sobemovirus. In Virus Taxonomy: Classification and Nomenclature of Viruses; King, A.M.Q., Lefkowitz, E., Adams, M.J., Carstens, E.B., Eds.; Elsevier Academic Press: Waltham, MA, USA, 2012; pp. 1185–1190. [Google Scholar]
- Olspert, A.; Arike, L.; Peil, L.; Truve, E. Sobemovirus RNA linked to VPg over a threonine residue. FEBS Lett. 2011, 585, 2979–2985. [Google Scholar] [CrossRef] [PubMed]
- Olspert, A.; Peil, L.; Hebrard, E.; Fargette, D.; Truve, E. Protein-RNA linkage and post-translational modifications of two sobemovirus VPgs. J. Gen. Virol. 2011, 92, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Hebrard, E.; Pinel-Galzi, A.; Bersoult, A.; Sire, C.; Fargette, D. Emergence of a resistance-breaking isolate of Rice yellow mottle virus during serial inoculations is due to a single substitution in the genome-linked viral protein VPg. J. Gen. Virol. 2006, 87, 1369–1373. [Google Scholar] [CrossRef] [PubMed]
- Hebrard, E.; Poulicard, N.; Gerard, C.; Traore, O.; Wu, H.C.; Albar, L.; Fargette, D.; Bessin, Y.; Vignols, F. Direct interaction between the rice yellow mottle virus (RYMV) VPg and the central domain of the rice eIF(iso)4G1 factor correlates with rice susceptibility and RYMV virulence. Mol. Plant Microbe Interact. 2010, 23, 1506–1513. [Google Scholar] [CrossRef] [PubMed]
- Mäkeläinen, K. Lost in Translation: Translation Mechanisms in Production of Cocksfoot Mottle Virus Proteins; University of Helsinki: Helsinki, Finland, 2006; p. 72. [Google Scholar]
- Dorokhov, Y.L.; Skulachev, M.V.; Ivanov, P.A.; Zvereva, S.D.; Tjulkina, L.G.; Merits, A.; Gleba, Y.Y.; Hohn, T.; Atabekov, J.G. Polypurine (A)-rich sequences promote cross-kingdom conservation of internal ribosome entry. Proc. Natl. Acad. Sci. USA 2002, 99, 5301–5306. [Google Scholar] [CrossRef] [PubMed]
- Yassi, M.N.; Ritzenthaler, C.; Brugidou, C.; Fauquet, C.; Beachy, R.N. Nucleotide sequence and genome characterization of rice yellow mottle virus RNA. J. Gen. Virol. 1994, 75, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Krutov, A.A.; Novikov, V.K.; Zheleznikova, O.V.; Morozov, S.Y.; Zavriev, S.K. Nucleotide sequence of RNA from the sobemovirus found in infected cocksfoot shows a luteovirus-like arrangement of the putative replicase and protease genes. Phytopathology 1996, 86, 391–397. [Google Scholar] [CrossRef]
- Wu, S.X.; Rinehart, C.A.; Kaesberg, P. Sequence and organization of southern bean mosaic virus genomic RNA. Virology 1987, 161, 73–80. [Google Scholar] [CrossRef]
- Othman, Y.; Hull, R. Nucleotide sequence of the bean strain of Southern bean mosaic virus. Virology 1995, 206, 287–297. [Google Scholar] [CrossRef]
- Lokesh, G.L.; Gopinath, K.; Satheshkumar, P.S.; Savithri, H.S. Complete nucleotide sequence of Sesbania mosaic virus: A new virus species of the genus Sobemovirus. Arch. Virol. 2001, 146, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Dreher, T.W. Functions of the 3′-untranslated regions of positive strand RNA viral genomes. Annu. Rev. Phytopathol. 1999, 37, 151–174. [Google Scholar] [CrossRef] [PubMed]
- Govind, K.; Savithri, H.S. Primer-independent initiation of RNA synthesis by SeMV recombinant RNA-dependent RNA polymerase. Virology 2010, 401, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Govind, K.; Mäkinen, K.; Savithri, H.S. Sesbania mosaic virus (SeMV) infectious clone: Possible mechanism of 3′ and 5′ end repair and role of polyprotein processing in viral replication. PLoS ONE 2012, 7, e31190. [Google Scholar] [CrossRef] [PubMed]
- Olspert, A.; Kamsol, K.; Sarmiento, C.; Gerassimenko, J.; Truve, E. Cocksfoot mottle virus coat protein is dispensable for the systemic infection. Virol J. 2014, 11, e19. [Google Scholar] [CrossRef] [PubMed]
- Govind, K.; Bakshi, A.; Savithri, H.S. Interaction of sesbania mosaic virus (SeMV) RNA-dependent RNA polymerase (RdRp) with the p10 domain of polyprotein 2a and its implications in SeMV replication. FEBS Open Bio 2014, 4, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.A.; Koev, G.; Mohan, B.R. Are there risks associated with transgenic resistance to luteoviruses? Plant Dis. 1997, 81, 700–710. [Google Scholar] [CrossRef]
- Mäkinen, K.; Tamm, T.; Naess, V.; Truve, E.; Puurand, U.; Munthe, T.; Saarma, M. Characterization of cocksfoot mottle sobemovirus genomic RNA and sequence comparison with related viruses. J. Gen. Virol. 1995, 76, 2817–2825. [Google Scholar] [CrossRef] [PubMed]
- Sereme, D.; Lacombe, S.; Konate, M.; Pinel-Galzi, A.; Traore, V.S.; Hebrard, E.; Traore, O.; Brugidou, C.; Fargette, D.; Konate, G. Biological and molecular characterization of a putative new sobemovirus infecting Imperata cylindrica and maize in Africa. Arch. Virol. 2008, 153, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Kokorev, N.V.; Koshkina, T.E.; Ryazantsev, D.Y.; Zavriev, S.K. Effect of cocksfoot mottle virus defective RNA on accumulation of virus capsid protein in wheat plants. Russ. Agric. Sci. 2007, 33, 152–154. [Google Scholar] [CrossRef]
- Rutgers, T.; Salerno-Rife, T.; Kaesberg, P. Messenger RNA for the coat protein of Southern bean mosaic virus. Virology 1980, 104, 506–509. [Google Scholar] [CrossRef]
- Salerno-Rife, T.; Rutgers, T.; Kaesberg, P. Translation of southern bean mosaic virus RNA in wheat embryo and rabbit reticulocyte extracts. J. Virol. 1980, 34, 51–58. [Google Scholar] [PubMed]
- Weber, K.A.; Sehgal, O.P. Subgenomic RNAs in virions of Southern bean mosaic virus. Phytopathology 1982, 72, 909–913. [Google Scholar] [CrossRef]
- Bonneau, C.; Brugidou, C.; Chen, L.; Beachy, R.N.; Fauquet, C. Expression of the rice yellow mottle virus P1 protein in vitro and in vivo and its involvement in virus spread. Virology 1998, 244, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Tamm, T.; Mäkinen, K.; Truve, E. Identification of genes encoding for the cocksfoot mottle virus proteins. Arch. Virol. 1999, 144, 1557–1567. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, K.; Generozov, E.; Arshava, N.; Kaloshin, A.; Morozov, S.; Zavriev, S. Detection and characterization of defective interfering RNAs associated with the cocksfoot mottle sobemovirus. Mol. Biol. 2000, 34, 332–338. [Google Scholar] [CrossRef]
- White, K.A.; Morris, T.J. Defective and defective interfering RNAs of monopartite plus-strand RNA plant viruses. Curr. Top. Microbiol. Immunol. 1999, 239, 1–17. [Google Scholar] [PubMed]
- Hu, C.C.; Hsu, Y.H.; Lin, N.S. Satellite RNAs and satellite viruses of plants. Viruses 2009, 1, 1325–1350. [Google Scholar] [CrossRef] [PubMed]
- Haseloff, J.; Symons, R.H. Comparative sequence and structure of viroid-like RNAs of two plant viruses. Nucleic Acids Res. 1982, 10, 3681–3691. [Google Scholar] [CrossRef] [PubMed]
- Keese, P.; Bruening, G.; Symons, R.H. Comparative sequence and structure of circular RNAs from 2 isolates of Lucerne transient streak virus. FEBS Lett. 1983, 159, 185–190. [Google Scholar] [CrossRef]
- Abouhaidar, M.G. Nucleotide sequence of the capsid protein gene and 3′ non-coding region of papaya mosaic virus RNA. J. Gen. Virol. 1988, 69, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Davies, C.; Haseloff, J.; Symons, R.H. Structure, self-cleavage, and replication of two viroid-like satellite RNAs (virusoids) of Subterranean clover mottle virus. Virology 1990, 177, 216–224. [Google Scholar] [CrossRef]
- Collins, R.F.; Gellatly, D.L.; Sehgal, O.P.; Abouhaidar, M.G. Self-cleaving circular RNA associated with Rice yellow mottle virus is the smallest viroid-like RNA. Virology 1998, 241, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.T.; Mayo, M.A. Satellite nature of the viroid-like RNA-2 of Solanum nodiflorum mottle virus and the ability of other plant-viruses to support the replication of viroid-like RNA molecules. J. Gen. Virol. 1984, 65, 1713–1721. [Google Scholar] [CrossRef]
- Sehgal, O.P.; Sinha, R.C.; Gellatly, D.L.; Ivanov, I.; AbouHaidar, M.G. Replication and encapsidation of the viroid-like satellite RNA of Lucerne transient streak virus are supported in divergent hosts by Cocksfoot mottle virus and Turnip rosette virus. J. Gen. Virol. 1993, 74, 785–788. [Google Scholar] [CrossRef] [PubMed]
- Francki, R.I.B. Possible viroid origin: Encapsidated viroid-like RNA. In The Viroids, 1st ed.; Diener, T.O., Ed.; Plenum Press: New York, NY, USA, 1987; Volume 1, pp. 205–218. [Google Scholar]
- Forster, A.C.; Symons, R.H. Self-cleavage of plus and minus RNAs of a virusoid and a structural model for the active-sites. Cell 1987, 49, 211–220. [Google Scholar] [CrossRef]
- Gellatly, D.; Mirhadi, K.; Venkataraman, S.; AbouHaidar, M.G. Structural and sequence integrity are essential for the replication of the viroid-like satellite RNA of Lucerne transient streak virus. J. Gen. Virol. 2011, 92, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- AbouHaidar, M.G.; Venkataraman, S.; Golshani, A.; Liu, B.; Ahmad, T. Novel coding, translation, and gene expression of a replicating covalently closed circular RNA of 220 nt. Proc. Natl. Acad. Sci. USA 2014, 111, 14542–14547. [Google Scholar] [CrossRef] [PubMed]
- Meier, M.; Paves, H.; Olspert, A.; Tamm, T.; Truve, E. P1 protein of Cocksfoot mottle virus is indispensable for the systemic spread of the virus. Virus Genes 2006, 32, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.R.; Savithri, H.S. Interaction of sesbania mosaic virus movement protein with the coat protein—Implications for viral spread. FEBS J. 2011, 278, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.R.; Savithri, H.S. Interaction of sesbania mosaic virus movement protein with VPg and P10: Implication to specificity of genome recognition. PLoS ONE 2011, 6, e15609. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Savithri, H.S. Natively unfolded nucleic acid binding P8 domain of SeMV polyprotein 2a affects the novel ATPase activity of the preceding P10 domain. FEBS Lett. 2010, 584, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Csorba, T.; Kontra, L.; Burgyan, J. Viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voinnet, O.; Pinto, Y.M.; Baulcombe, D.C. Suppression of gene silencing: A general strategy used by diverse DNA and RNA viruses of plants. Proc. Natl. Acad. Sci. USA 1999, 96, 14147–14152. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento, C.; Gomez, E.; Meier, M.; Kavanagh, T.A.; Truve, E. Cocksfoot mottle virus P1 suppresses RNA silencing in Nicotiana benthamiana and Nicotiana tabacum. Virus Res. 2007, 123, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Sire, C.; Bangratz-Reyser, M.; Fargette, D.; Brugidou, C. Genetic diversity and silencing suppression effects of Rice yellow mottle virus and the P1 protein. Virol. J. 2008, 5, e55. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, A.F.; Correa, R.L.; Nakasugi, K.; Jackson, C.; Kawchuk, L.; Vaslin, M.F.; Waterhouse, P.M. The enamovirus P0 protein is a silencing suppressor which inhibits local and systemic RNA silencing through AGO1 degradation. Virology 2012, 426, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, S.; Bangratz, M.; Vignols, F.; Brugidou, C. The rice yellow mottle virus P1 protein exhibits dual functions to suppress and activate gene silencing. Plant J. 2010, 61, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.A.; Sarmiento, C.; Kiisma, M.; Koivumaki, S.; Lemmetty, A.; Truve, E.; Lehto, K. Effects of viral silencing suppressors on tobacco ringspot virus infection in two Nicotiana species. J. Gen. Virol. 2008, 89, 1502–1508. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Chen, Z.; Song, X.; Liu, C.; Cui, X.; Zhao, X.; Fang, J.; Xu, W.; Zhang, H.; Wang, X.; et al. Oryza sativa dicer-like4 reveals a key role for small interfering RNA silencing in plant development. Plant Cell 2007, 19, 2705–2718. [Google Scholar] [CrossRef] [PubMed]
- Tamm, T.; Truve, E. RNA-binding activities of cocksfoot mottle sobemovirus proteins. Virus Res. 2000, 66, 197–207. [Google Scholar] [CrossRef]
- Van Wezel, R.; Liu, H.; Wu, Z.; Stanley, J.; Hong, Y. Contribution of the zinc finger to zinc and DNA binding by a suppressor of posttranscriptional gene silencing. J. Virol. 2003, 77, 696–700. [Google Scholar] [CrossRef] [PubMed]
- Chiba, S.; Hleibieh, K.; Delbianco, A.; Klein, E.; Ratti, C.; Ziegler-Graff, V.; Bouzoubaa, S.; Gilmer, D. The benyvirus RNA silencing suppressor is essential for long-distance movement, requires both zinc-finger and NoLS basic residues but not a nucleolar localization for its silencing-suppression activity. Mol. Plant Microbe Interact. 2013, 26, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Gillet, F.X.; Cattoni, D.I.; Petiot-Becard, S.; Delalande, F.; Poignavent, V.; Brizard, J.P.; Bessin, Y.; Dorsselaer, A.V.; Declerck, N.; Sanglier-Cianferani, S.; et al. The RYMV-encoded viral suppressor of RNA silencing P1 is a zinc-binding protein with redox-dependent flexibility. J. Mol. Biol. 2013, 425, 2423–2435. [Google Scholar] [CrossRef] [PubMed]
- Sereme, D.; Lacombe, S.; Konate, M.; Bangratz, M.; Pinel-Galzi, A.; Fargette, D.; Traore, A.S.; Konate, G.; Brugidou, C. Sites under positive selection modulate the RNA silencing suppressor activity of rice yellow mottle virus movement protein P1. J. Gen. Virol. 2014, 95, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Weinheimer, I.; Boonrod, K.; Moser, M.; Zwiebel, M.; Fullgrabe, M.; Krczal, G.; Wassenegger, M. Analysis of an autoproteolytic activity of rice yellow mottle virus silencing suppressor P1. Biol. Chem. 2010, 391, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Folimonov, A.; Shintaku, M.; Li, W.X.; Falk, B.W.; Dawson, W.O.; Ding, S.W. Three distinct suppressors of RNA silencing encoded by a 20-kb viral RNA genome. Proc. Natl. Acad. Sci. USA 2004, 101, 15742–15747. [Google Scholar] [CrossRef] [PubMed]
- Senshu, H.; Yamaji, Y.; Minato, N.; Shiraishi, T.; Maejima, K.; Hashimoto, M.; Miura, C.; Neriya, Y.; Namba, S. A dual strategy for the suppression of host antiviral silencing: Two distinct suppressors for viral replication and viral movement encoded by potato virus M. J. Virol. 2011, 85, 10269–10278. [Google Scholar] [CrossRef] [PubMed]
- Revill, P.A.; Davidson, A.D.; Wright, P.J. Mushroom bacilliform virus RNA: The initiation of translation at the 5′ end of the genome and identification of the VPg. Virology 1998, 249, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Van der Wilk, F.; Verbeek, M.; Dullemans, A.; van den Heuvel, J. The genome-linked protein (VPg) of southern bean mosaic virus is encoded by the ORF2. Virus Genes 1998, 17, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Wobus, C.E.; Skaf, J.S.; Schultz, M.H.; de Zoeten, G.A. Sequencing, genomic localization and initial characterization of the VPg of pea enation mosaic enamovirus. J. Gen. Virol. 1998, 79, 2023–2025. [Google Scholar] [PubMed]
- Van der Wilk, F.; Verbeek, M.; Dullemans, A.M.; van den Heuvel, J.F. The genome-linked protein of Potato leafroll virus is located downstream of the putative protease domain of the ORF1 product. Virology 1998, 234, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Satheshkumar, P.S.; Lokesh, G.L.; Savithri, H.S. Polyprotein processing: Cis and trans proteolytic activities of sesbania mosaic virus serine protease. Virology 2004, 318, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Koonin, E.V.; Blinov, V.M.; Donchenko, A.P. Sobemovirus genome appears to encode a serine protease related to cysteine proteases of picornaviruses. FEBS Lett. 1988, 236, 287–290. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Donchenko, A.P.; Blinov, V.M.; Koonin, E.V. Cysteine proteases of positive strand RNA viruses and chymotrypsin-like serine proteases. A distinct protein superfamily with a common structural fold. FEBS Lett. 1989, 243, 103–114. [Google Scholar] [CrossRef]
- Koonin, E.V.; Dolja, V.V. Evolution and taxonomy of positive-strand RNA viruses: Implications of comparative analysis of amino acid sequences. Crit. Rev. Biochem. Mol. Biol. 1993, 28, 375–430. [Google Scholar] [CrossRef] [PubMed]
- Gayathri, P.; Satheshkumar, P.S.; Prasad, K.; Nair, S.; Savithri, H.S.; Murthy, M.R. Crystal structure of the serine protease domain of sesbania mosaic virus polyprotein and mutational analysis of residues forming the S1-binding pocket. Virology 2006, 346, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Sõmera, M.; Truve, E. The genome organization of lucerne transient streak and turnip rosette sobemoviruses revisited. Arch. Virol. 2013, 158, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Satheshkumar, P.S.; Gayathri, P.; Prasad, K.; Savithri, H.S. “Natively unfolded” VPg is essential for sesbania mosaic virus serine protease activity. J. Biol. Chem. 2005, 280, 30291–30300. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Gayathri, P.; Murthy, M.R.; Savithri, H.S. Stacking interactions of W271 and H275 of SeMV serine protease with W43 of natively unfolded VPg confer catalytic activity to protease. Virology 2008, 382, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, K.; Mäkeläinen, K.; Arshava, N.; Tamm, T.; Merits, A.; Truve, E.; Zavriev, S.; Saarma, M. Characterization of VPg and the polyprotein processing of Cocksfoot mottle virus (genus Sobemovirus). J. Gen. Virol. 2000, 81, 2783–2789. [Google Scholar] [PubMed]
- Uversky, V.N. What does it mean to be natively unfolded? Eur. J. Biochem. 2002, 269, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Hebrard, E.; Bessin, Y.; Michon, T.; Longhi, S.; Uversky, V.N.; Delalande, F.; van Dorsselaer, A.; Romero, P.; Walter, J.; Declerck, N.; et al. Intrinsic disorder in viral proteins genome-linked: Experimental and predictive analyses. Virol. J. 2009, 6, e23. [Google Scholar] [CrossRef] [PubMed]
- Pinel-Galzi, A.; Rakotomalala, M.; Sangu, E.; Sorho, F.; Kanyeka, Z.; Traore, O.; Sereme, D.; Poulicard, N.; Rabenantoandro, Y.; Sere, Y.; et al. Theme and variations in the evolutionary pathways to virulence of an RNA plant virus species. PLoS Pathog. 2007, 3, e180. [Google Scholar] [CrossRef] [PubMed]
- Poulicard, N.; Pinel-Galzi, A.; Traore, O.; Vignols, F.; Ghesquiere, A.; Konate, G.; Hebrard, E.; Fargette, D. Historical contingencies modulate the adaptability of Rice yellow mottle virus. PLoS Pathog. 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Traore, O.; Pinel-Galzi, A.; Issaka, S.; Poulicard, N.; Aribi, J.; Ake, S.; Ghesquiere, A.; Sere, Y.; Konate, G.; Hebrard, E.; et al. The adaptation of Rice yellow mottle virus to the eIF(iso)4G-mediated rice resistance. Virology 2010, 408, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Poulicard, N.; Pinel-Galzi, A.; Fargette, D.; Hebrard, E. Alternative mutational pathways, outside the VPg, of Rice yellow mottle virus to overcome eIF(iso)4G-mediated rice resistance under strong genetic constraints. J. Gen. Virol. 2014, 95, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Buck, K.W. Comparison of the replication of positive-stranded RNA viruses of plants and animals. Adv. Virus Res. 1996, 47, 159–251. [Google Scholar] [PubMed]
- Koonin, E.V. The phylogeny of RNA-dependent RNA polymerases of positive-strand RNA viruses. J. Gen. Virol. 1991, 72, 2197–2206. [Google Scholar] [CrossRef] [PubMed]
- Abad-Zapatero, C.; Abdel-Meguid, S.S.; Johnson, J.E.; Leslie, A.G.; Rayment, I.; Rossmann, M.G.; Suck, D.; Tsukihara, T. Structure of Southern bean mosaic virus at 2.8 Å resolution. Nature 1980, 286, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Bhuvaneshwari, M.; Subramanya, H.S.; Gopinath, K.; Savithri, H.S.; Nayudu, M.V.; Murthy, M.R. Structure of Sesbania mosaic virus at 3 Å resolution. Structure 1995, 3, 1021–1030. [Google Scholar] [CrossRef]
- Qu, C.; Liljas, L.; Opalka, N.; Brugidou, C.; Yeager, M.; Beachy, R.N.; Fauquet, C.M.; Johnson, J.E.; Lin, T. 3D domain swapping modulates the stability of members of an icosahedral virus group. Structure 2000, 8, 1095–1103. [Google Scholar] [CrossRef]
- Tars, K.; Zeltins, A.; Liljas, L. The three-dimensional structure of Cocksfoot mottle virus at 2.7 Å resolution. Virology 2003, 310, 287–297. [Google Scholar] [CrossRef]
- Plevka, P.; Tars, K.; Zeltins, A.; Balke, I.; Truve, E.; Liljas, L. The three-dimensional structure of Ryegrass mottle virus at 2.9 Å resolution. Virology 2007, 369, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Rossmann, M.G.; Johnson, J.E. Icosahedral RNA virus structure. Annu. Rev. Biochem. 1989, 58, 533–573. [Google Scholar] [CrossRef] [PubMed]
- Dolja, V.V.; Koonin, E.V. Phylogeny of capsid proteins of small icosahedral RNA plant viruses. J. Gen. Virol. 1991, 72, 1481–1486. [Google Scholar] [CrossRef] [PubMed]
- Sõmera, M.; Truve, E. Rottboellia yellow mottle virus is a distinct species within the genus Sobemovirus. Arch. Virol. 2015, 160, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Tripp, M.; Shepherd, C.M.; Borelli, I.A.; Venkataraman, S.; Lander, G.; Natarajan, P.; Johnson, J.E.; Brooks, C.L.; Reddy, V.S. VIPERdb(2): An enhanced and web API enabled relational database for structural virology. Nucleic Acids Res. 2009, 37, D436–D442. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.H.; Sehgal, O.P.; Pickett, E.E. Stabilizing effect of divalent metal ions on virions of Southern bean mosaic virus. Virology 1976, 69, 587–595. [Google Scholar] [CrossRef]
- Kruse, J.; Timmins, P.A.; Witz, J. A neutron-scattering study of the structure of compact and swollen forms of Southern bean mosaic virus. Virology 1982, 119, 42–50. [Google Scholar] [CrossRef]
- Satheshkumar, P.S.; Lokesh, G.L.; Sangita, V.; Saravanan, V.; Vijay, C.S.; Murthy, M.R.; Savithri, H.S. Role of metal ion-mediated interactions in the assembly and stability of sesbania mosaic virus T=3 and T=1 capsids. J. Mol. Biol. 2004, 342, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Tama, F.; Brooks, C.L. Diversity and identity of mechanical properties of icosahedral viral capsids studied with elastic network normal mode analysis. J. Mol. Biol. 2005, 345, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Opalka, N.; Tihova, M.; Brugidou, C.; Kumar, A.; Beachy, R.N.; Fauquet, C.M.; Yeager, M. Structure of native and expanded sobemoviruses by electron cryo-microscopy and image reconstruction. J. Mol. Biol. 2000, 303, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Hermodson, M.A.; Abadzapatero, C.; Abdelmeguid, S.S.; Pundak, S.; Rossmann, M.G.; Tremaine, J.H. Amino-acid-sequence of southern bean mosaic-virus coat protein and its relation to the 3-dimensional structure of the virus. Virology 1982, 119, 133–149. [Google Scholar] [CrossRef]
- Sangita, V.; Lokesh, G.L.; Satheshkumar, P.S.; Vijay, C.S.; Saravanan, V.; Savithri, H.S.; Murthy, M.R. T=1 capsid structures of sesbania mosaic virus coat protein mutants: Determinants of T=3 and T=1 capsid assembly. J. Mol. Biol. 2004, 342, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Brugidou, C.; Opalka, N.; Yeager, M.; Beachy, R.N.; Fauquet, C. Stability of Rice yellow mottle virus and cellular compartmentalization during the infection process in Oryza sativa (L.). Virology 2002, 297, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Pappachan, A.; Chinnathambi, S.; Satheshkumar, P.S.; Savithri, H.S.; Murthy, M.R. A single point mutation disrupts the capsid assembly in Sesbania mosaic virus resulting in a stable isolated dimer. Virology 2009, 392, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Rossmann, M.G.; Abad-Zapatero, C.; Hermodson, M.A.; Erickson, J.W. Subunit interactions in Southern bean mosaic virus. J. Mol. Biol. 1983, 166, 37–73. [Google Scholar] [CrossRef]
- Lokesh, G.L.; Gowri, T.D.; Satheshkumar, P.S.; Murthy, M.R.; Savithri, H.S. A molecular switch in the capsid protein controls the particle polymorphism in an icosahedral virus. Virology 2002, 292, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Satheshkumar, P.S.; Lokesh, G.L.; Murthy, M.R.; Savithri, H.S. The role of arginine-rich motif and beta-annulus in the assembly and stability of sesbania mosaic virus capsids. J. Mol. Biol. 2005, 353, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Savithri, H.S.; Erickson, J.W. The self-assembly of the cowpea strain of Southern bean mosaic virus: Formation of T=1 and T=3 nucleoprotein particles. Virology 1983, 126, 328–335. [Google Scholar] [CrossRef]
- Hacker, D.L. Identification of a coat protein binding site on southern bean mosaic virus RNA. Virology 1995, 207, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.S. The anatomy and taxonomy of protein structure. Adv. Protein Chem. 1981, 34, 167–339. [Google Scholar] [PubMed]
- Erickson, J.W.; Rossmann, M.G. Assembly and crystallization of a T=1 icosahedral particle from trypsinized southern bean mosaic virus coat protein. Virology 1982, 116, 128–136. [Google Scholar] [CrossRef]
- Pappachan, A.; Subashchandrabose, C.; Satheshkumar, P.S.; Savithri, H.S.; Murthy, M.R. Structure of recombinant capsids formed by the beta-annulus deletion mutant—rCP (Delta48-59) of Sesbania mosaic virus. Virology 2008, 375, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Hacker, D.L. In vitro analysis of an RNA binding site within the N-terminal 30 amino acids of the southern cowpea mosaic virus coat protein. Virology 2001, 286, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Olspert, A.; Paves, H.; Toomela, R.; Tamm, T.; Truve, E. Cocksfoot mottle sobemovirus coat protein contains two nuclear localization signals. Virus Genes 2010, 40, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Dabney-Smith, C.; Hacker, D.L.; Bruce, B.D. Membrane activity of the southern cowpea mosaic virus coat protein: The role of basic amino acids, helix-forming potential, and lipid composition. Virology 2001, 291, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Belyi, V.A.; Muthukumar, M. Electrostatic origin of the genome packing in viruses. Proc. Natl. Acad. Sci. USA 2006, 103, 17174–17178. [Google Scholar] [CrossRef] [PubMed]
- Savithri, H.S.; Murthy, M.R.N. Structure and assembly of Sesbania mosaic virus. Curr. Sci. 2010, 98, 346–351. [Google Scholar]
- Zink, M.; Grubmuller, H. Primary changes of the mechanical properties of Southern bean mosaic virus upon calcium removal. Biophys. J. 2010, 98, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.M.; Cachau, R.E.; Goldstein, D.J. Ion channels in southern bean mosaic virus capsid. Biophys. J. 1987, 52, 595–602. [Google Scholar] [CrossRef]
- Fuentes, A.L.; Hamilton, R.I. Failure of long-distance movement of Southern bean mosaic virus in a resistant host is correlated with lack of normal virion formation. J. Gen. Virol. 1993, 74, 1903–1910. [Google Scholar] [CrossRef] [PubMed]
- Opalka, N.; Brugidou, C.; Bonneau, C.; Nicole, M.; Beachy, R.N.; Yeager, M.; Fauquet, C. Movement of Rice yellow mottle virus between xylem cells through pit membranes. Proc. Natl. Acad. Sci. USA 1998, 95, 3323–3328. [Google Scholar] [CrossRef] [PubMed]
- Kouassi, N.K.; Chen, L.; Sire, C.; Bangratz-Reyser, M.; Beachy, R.N.; Fauquet, C.M.; Brugidou, C. Expression of rice yellow mottle virus coat protein enhances virus infection in transgenic plants. Arch. Virol. 2006, 151, 2111–2122. [Google Scholar] [CrossRef] [PubMed]
- Callaway, A.S.; George, C.G.; Lommel, S.A. A sobemovirus coat protein gene complements long-distance movement of a coat protein-null dianthovirus. Virology 2004, 330, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Gergerich, R.C.; Scott, H.A.; Fulton, J.P. Evidence that ribonuclease in beetle regurgitant determines the transmission of plant-viruses. J. Gen. Virol. 1986, 67, 367–370. [Google Scholar] [CrossRef]
- Wang, R.Y.; Gergerich, R.C.; Kim, K.S. Entry of ingested plant-viruses into the hemocoel of the beetle vector Diabrotica undecimpunctata Howardi. Phytopathology 1994, 84, 147–153. [Google Scholar] [CrossRef]
- Gibb, K.S.; Randles, J.W. Distribution of Velvet tobacco mottle virus in its mirid vector and its relationship to transmissibility. Ann. Appl. Biol. 1990, 116, 513–521. [Google Scholar] [CrossRef]
- Meier, M.; Olspert, A.; Sarmiento, C.; Truve, E. Sobemoviruses. In Encyclopedia of Virology, 3rd ed.; Mahy, B., van Regenmortel, M., Eds.; Elsevier: Oxford, UK, 2008; Volume 5, pp. 644–652. [Google Scholar]
- Weintraub, M.; Ragetli, H.W. Identification of the constituents of Southern bean mosaic virus in crystals of infected cells, and their distribution within the virion. Virology 1970, 41, 729–739. [Google Scholar] [CrossRef]
- Lombardo, G.; Simonetta, M.; Bassi, M. An ultrastructural study of the localization of Sowbane mosaic virus. Caryologia 1971, 24, 323–330. [Google Scholar] [CrossRef]
- Chamberlain, J.A.; Catherall, P.L. Electron microscopy of some grasses and cereals infected with cocksfoot mottle, phleum mottle and cocksfoot mild mosaic viruses. J. Gen. Virol. 1976, 30, 41–50. [Google Scholar] [CrossRef]
- Descriptions of Plant Viruses. Available online: http://www.dpvweb.net (accessed on 18 May 2015).
- Gergerich, R.C. Beetles. Adv. Bot. Res. 2002, 36, 101–112. [Google Scholar]
- Kozela, C.; Regan, S. How plants make tubes. Trends Plant Sci. 2003, 8, 159–164. [Google Scholar] [CrossRef]
- Hartmann, J.X.; Bath, J.E.; Hooper, G.R. Electron microscopy of viruslike particles from shoestring-diseased highbush blueberry, Vaccinium corymbosum L. Phytopathology 1973, 63, 432–436. [Google Scholar] [CrossRef]
- Urban, L.A.; Ramsdell, D.C.; Klomparens, K.L.; Lynch, T.; Hancock, J.F. Detection of blueberry shoestring virus in xylem and phloem tissues of highbush blueberry. Phytopathology 1989, 79, 488–493. [Google Scholar] [CrossRef]
- Schneider, I.R.; Worley, J.F. Upward and downward transport of infectious particles of Southern bean mosaic virus through steamed portions of bean stems. Virology 1959, 8, 230–242. [Google Scholar] [CrossRef]
- Morales, E.J.; Castano, M.; Arroyave, J.A.; Ospina, M.D.; Calvert, L.A. A sobemovirus hindering the utilization of Calopogonium mucunoides as a forage legume in the Lowland tropics. Plant Dis. 1995, 79, 1220–1224. [Google Scholar] [CrossRef]
- Otsus, M.; Uffert, G.; Sõmera, M.; Paves, H.; Olspert, A.; Islamov, B.; Truve, E. Cocksfoot mottle sobemovirus establishes infection through the phloem. Virus Res. 2012, 166, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Thurston, M.I.; Pallett, D.W.; Cortina-Borja, M.; Edwards, M.L.; Raybould, A.F.; Cooper, J.I. The incidence of viruses in wild Brassica nigra in Dorset (UK). Ann. Appl. Biol. 2001, 139, 277–284. [Google Scholar] [CrossRef]
- Pallett, D.W.; Thurston, M.I.; Cortina-Borja, M.; Edwards, M.L.; Alexander, M.; Mitchell, E.; Raybould, A.F.; Cooper, J.I. The incidence of viruses in wild Brassica rapa ssp sylvestris in southern UK. Ann. Appl. Biol. 2002, 141, 163–170. [Google Scholar] [CrossRef]
- Forster, R.L.S.; Jones, A.T. Properties of Lucerne transient streak virus, and evidence of its affinity to Southern bean mosaic virus. Ann. Appl. Biol. 1979, 93, 181–189. [Google Scholar] [CrossRef]
- Yao, N.; Imai, S.; Tada, Y.; Nakayashiki, H.; Tosa, Y.; Park, P.; Mayama, S. Apoptotic cell death is a common response to pathogen attack in oats. Mol. Plant. Microbe Interact. 2002, 15, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Brizard, J.P.; Carapito, C.; Delalande, F.; van Dorsselaer, A.; Brugidou, C. Proteome analysis of plant-virus interactome: Comprehensive data for virus multiplication inside their hosts. Mol. Cell. Proteomics 2006, 5, 2279–2297. [Google Scholar] [CrossRef] [PubMed]
- Ventelon-Debout, M.; Delalande, F.; Brizard, J.P.; Diemer, H.; van Dorsselaer, A.; Brugidou, C. Proteome analysis of cultivar-specific deregulations of Oryza sativa indica and O. sativa japonica cellular suspensions undergoing rice yellow mottle virus infection. Proteomics 2004, 4, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.L.; Burritt, D.J. The influence of Cocksfoot mottle virus on antioxidant metabolism in the leaves of Dactylis glomerata L. Physiol. Mol. Plant Pathol. 2003, 62, 285–295. [Google Scholar] [CrossRef]
- Catherall, P.L. Resistances of grasses to 2 sobemoviruses, cocksfoot mottle and cynosurus mottle. Grass Forage Sci. 1985, 40, 311–316. [Google Scholar] [CrossRef]
- Rognli, O.A.; Aastveit, K.; Munthe, T. Genetic variation in cocksfoot (Dactylis glomerata L.) populations for mottle virus resistance. Euphytica 1995, 83, 109–116. [Google Scholar] [CrossRef]
- Thottappilly, G.; Rossel, H.W. Evaluation of resistance to Rice yellow mottle virus in Oryza species. Indian J. Virol. 1993, 9, 65–73. [Google Scholar]
- Ghesquiere, A.; Albar, L.; Lorieux, M.; Ahmadi, N.; Fargette, D.; Huang, N.; McCouch, S.R.; Notteghem, J.L. A major quantitative trait locus for rice yellow mottle virus resistance maps to a cluster of blast resistance genes on chromosome 12. Phytopathology 1997, 87, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Ndjiondjop, M.N.; Albar, L.; Fargette, D.; Fauquet, C.; Ghesquiere, A. The genetic basis of high resistance to Rice yellow mottle virus (RYMV) in cultivars of two cultivated rice species. Plant Dis. 1999, 83, 931–935. [Google Scholar] [CrossRef]
- Paul, C.N.; Ng, N.Q.; Ladeinde, T. Diallel analysis of resistance to Rice yellow mottle virus in African rice Oryza glaberrima Steud. J. Genet. Breed. 1995, 49, 217–222. [Google Scholar]
- Zaumeyer, W.J.; Harter, L.L. Two new virus diseases of beans. J. Agric. Res. 1943, 67, 305–328. [Google Scholar]
- Hobbs, H.A.; Kuhn, C.W.; Papa, K.E.; Brantley, B.B. Inheritance of non-necrotic resistance to Southern bean mosaic virus in cowpea. Phytopathology 1987, 77, 1624–1629. [Google Scholar] [CrossRef]
- Ferris, D.G.; Jones, R.A.C.; Wroth, J.M. Determining the effectiveness of resistance to subterranean clover mottle sobemovirus in different genotypes of subterranean clover in the field using the grazing animal as virus vector. Ann. Appl. Biol. 1996, 128, 303–315. [Google Scholar] [CrossRef]
- Wroth, J.M.; Jones, R.A.C. Subterranean clover mottle sobemovirus: Its host range, resistance in subterranean clover and transmission through seed and by grazing animals. Ann. Appl. Biol. 1992, 121, 329–343. [Google Scholar] [CrossRef]
- Albar, L.; Ndjiondjop, M.N.; Esshak, Z.; Berger, A.; Pinel, A.; Jones, M.; Fargette, D.; Ghesquiere, A. Fine genetic mapping of a gene required for rice yellow mottle virus cell-to-cell movement. Theor. Appl. Genet. 2003, 107, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albar, L.; Bangratz-Reyser, M.; Hebrard, E.; Ndjiondjop, M.N.; Jones, M.; Ghesquiere, A. Mutations in the eIF(iso)4G translation initiation factor confer high resistance of rice to Rice yellow mottle virus. Plant J. 2006, 47, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Thiemele, D.; Boisnard, A.; Ndjiondjop, M.N.; Cheron, S.; Sere, Y.; Ake, S.; Ghesquiere, A.; Albar, L. Identification of a second major resistance gene to Rice yellow mottle virus, rymv2, in the African cultivated rice species, O. glaberrima. Theor. Appl. Genet. 2010, 121, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Rakotomalala, M.; Pinel-Galzi, A.; Albar, L.; Ghesquiere, A.; Rabenantoandro, Y.; Ramavovololona, P.; Fargette, D. Resistance to Rice yellow mottle virus in rice germplasm in Madagascar. Eur. J. Plant Pathol. 2008, 122, 277–286. [Google Scholar] [CrossRef]
- Hebrard, E.; Pinel-Galzi, A.; Fargette, D. Virulence domain of the RYMV genome-linked viral protein VPg towards rice rymv1-2-mediated resistance. Arch. Virol. 2008, 153, 1161–1164. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, N.; Albar, L.; Pressoir, G.; Pinel, A.; Fargette, D.; Ghesquiere, A. Genetic basis and mapping of the resistance to Rice yellow mottle virus. III. Analysis of QTL efficiency in introgressed progenies confirmed the hypothesis of complementary epistasis between two resistance QTLs. Theor. Appl. Genet. 2001, 103, 1084–1092. [Google Scholar]
- Albar, L.; Lorieux, M.; Ahmadi, N.; Rimbault, I.; Pinel, A.; Sy, A.A.; Fargette, D.; Ghesquiere, A. Genetic basis and mapping of the resistance to Rice yellow mottle virus. I. QTLs identification and relationship between resistance and plant morphology. Theor. Appl. Genet. 1998, 97, 1145–1154. [Google Scholar] [CrossRef]
- Ioannidou, D.; Lett, J.M.; Pinel, A.; Assigbetse, K.; Brugidou, C.; Ghesquiere, A.; Nicole, M.; Fargette, D. Responses of Oryza sativa japonica sub-species to infection with Rice yellow mottle virus. Physiol. Mol. Plant Pathol. 2000, 57, 177–188. [Google Scholar] [CrossRef]
- Boisnard, A.; Albar, L.; Thiemele, D.; Rondeau, M.; Ghesquiere, A. Evaluation of genes from eIF4E and eIF4G multigenic families as potential candidates for partial resistance QTLs to Rice yellow mottle virus in rice. Theor. Appl. Genet. 2007, 116, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Orjuela, J.; Deless, E.F.; Kolade, O.; Cheron, S.; Ghesquiere, A.; Albar, L. A recessive resistance to Rice yellow mottle virus is associated with a rice homolog of the cpr5 gene, a regulator of active defense mechanisms. Mol. Plant Microbe Interact. 2013, 26, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Fargette, D.; Pinel-Galzi, A.; Sereme, D.; Lacombe, S.; Hebrard, E.; Traore, O.; Konate, G. Diversification of Rice yellow mottle virus and related viruses spans the history of agriculture from the neolithic to the present. PLoS Pathog. 2008, 4, e1000125. [Google Scholar] [CrossRef] [PubMed]
- Arthur, K.; Dogra, S.; Randles, J.W. Complete nucleotide sequence of velvet tobacco mottle virus isolate K1. Arch. Virol. 2010, 155, 1893–1896. [Google Scholar] [CrossRef] [PubMed]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Pagan, I.; Holmes, E.C. Long-term evolution of the Luteoviridae: Time scale and mode of virus speciation. J. Virol. 2010, 84, 6177–6187. [Google Scholar] [CrossRef] [PubMed]
- Arthur, K.; Collins, N.C.; Randles, J.W. Mutation rate in Velvet tobacco mottle virus varies between genomic region and virus variant but is not influenced by obligatory mirid transmission. Virus Genes 2012, 45, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.R.; Keese, P.K.; Young, M.J.; Waterhouse, P.M.; Gerlach, W.L. Evolution and molecular biology of luteoviruses. Annu. Rev. Phytopathol. 1990, 28, 341–363. [Google Scholar] [CrossRef]
- Domier, L. Family—Luteoviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses; King, A.M.Q., Lefkowitz, E., Adams, M.J., Carstens, E.B., Eds.; Elsevier Academic Press: Waltham, MA, USA, 2012; pp. 1045–1053. [Google Scholar]
- Gunawardene, C.D.; Jaluba, K.; White, K.A. Conserved motifs in a tombusvirus polymerase modulate genome replication, subgenomic transcription, and amplification of defective interfering RNAs. J. Virol. 2015, 89, 3236–3246. [Google Scholar] [CrossRef] [PubMed]
- Revill, P.A.; Davidson, A.D.; Wright, P.J. The nucleotide sequence and genome organization of Mushroom bacilliform virus: A single-stranded RNA virus of Agaricus bisporus (Lange) Imbach. Virology 1994, 202, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Aus dem Siepen, M.; Pohl, J.O.; Koo, B.J.; Wege, C.; Jeske, H. Poinsettia latent virus is not a cryptic virus, but a natural polerovirus-sobemovirus hybrid. Virology 2005, 336, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Boulila, M. Selective constraints, molecular recombination structure and phylogenetic reconstruction of isometric plant RNA viruses of the families Luteoviridae and Tymoviridae. Biochimie 2011, 93, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.M.S.; Felix, M.R.; Clara, M.I.E.; Oliveira, S. The complete genome sequence of a new necrovirus isolated from Olea europaea L. Arch. Virol. 2005, 150, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Xi, D.; Li, J.; Han, C.; Li, D.; Yu, J.; Zhou, X. Complete nucleotide sequence of a new strain of tobacco necrosis virus A infecting soybean in China and infectivity of its full-length cDNA clone. Virus Genes 2008, 36, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Boulila, M. Positive selection, molecular recombination structure and phylogenetic reconstruction of members of the family Tombusviridae: Implication in virus taxonomy. Genet. Mol. Biol. 2011, 34, 647–660. [Google Scholar] [CrossRef] [PubMed]
- Chare, E.R.; Holmes, E.C. A phylogenetic survey of recombination frequency in plant RNA viruses. Arch. Virol. 2006, 151, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Meier, M.; Truve, E. An attempt to identify recombinants between two sobemoviruses in doubly infected oat plants. Environ. Biosaf. Res. 2006, 5, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Pinel-Galzi, A.; Mpunami, A.; Sangu, E.; Rakotomalala, M.; Traore, O.; Sereme, D.; Sorho, F.; Sere, Y.; Kanyeka, Z.; Konate, G.; et al. Recombination, selection and clock-like evolution of Rice yellow mottle virus. Virology 2009, 394, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Rakotomalala, M.; Pinel-Galzi, A.; Mpunami, A.; Randrianasolo, A.; Ramavovololona, P.; Rabenantoandro, Y.; Fargette, D. Rice yellow mottle virus in Madagascar and in the Zanzibar archipelago; island systems and evolutionary time scale to study virus emergence. Virus Res. 2013, 171, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Rochon, D.; Lommel, S.; Martelli, G.P.; Rubino, L.; Russo, M. Family Tombusviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses; King, A.M.Q., Lefkowitz, E., Adams, M.J., Carstens, E.B., Eds.; Elsevier Academic Press: Waltham, MA, USA, 2012; pp. 1111–1138. [Google Scholar]
- Adams, M.J.; Jeske, H. Genus Polemovirus. In Virus Taxonomy: Classification and Nomenclature of Viruses; King, A.M.Q., Lefkowitz, E., Adams, M.J., Carstens, E.B., Eds.; Elsevier Academic Press: Waltham, MA, USA, 2012; Volume 9, pp. 1181–1184. [Google Scholar]
- Revill, P.A. Family Barnaviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses; King, A.M.Q., Lefkowitz, E., Adams, M.J., Carstens, E.B., Eds.; Elsevier Academic Press: Waltham, MA, USA, 2012; pp. 961–964. [Google Scholar]
- Dolja, V.V.; Koonin, E.V. Common origins and host-dependent diversity of plant and animal viromes. Curr. Opin. Virol. 2011, 1, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, E.; Firth, A.E.; Miller, W.A.; Scheidecker, D.; Brault, V.; Reinbold, C.; Rakotondrafara, A.M.; Chung, B.Y.; Ziegler-Graff, V. Discovery of a small non-AUG-initiated ORF in poleroviruses and luteoviruses that is required for long-distance movement. PLoS Pathog. 2015, 11, e1004868. [Google Scholar] [CrossRef] [PubMed]
- Scheets, K. Infectious transcripts of an asymptomatic panicovirus identified from a metagenomic survey. Virus Res. 2013, 176, 161–168. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sõmera, M.; Sarmiento, C.; Truve, E. Overview on Sobemoviruses and a Proposal for the Creation of the Family Sobemoviridae. Viruses 2015, 7, 3076-3115. https://0-doi-org.brum.beds.ac.uk/10.3390/v7062761
Sõmera M, Sarmiento C, Truve E. Overview on Sobemoviruses and a Proposal for the Creation of the Family Sobemoviridae. Viruses. 2015; 7(6):3076-3115. https://0-doi-org.brum.beds.ac.uk/10.3390/v7062761
Chicago/Turabian StyleSõmera, Merike, Cecilia Sarmiento, and Erkki Truve. 2015. "Overview on Sobemoviruses and a Proposal for the Creation of the Family Sobemoviridae" Viruses 7, no. 6: 3076-3115. https://0-doi-org.brum.beds.ac.uk/10.3390/v7062761