
Whole-Genome Analysis of a Novel Fish Reovirus (MsReV) Discloses Aquareovirus Genomic Structure Relationship with Host in Saline Environments

Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Isolation, Electron Microscopy and Electrophoretic Analysis
2.2. Viral Genome Sequencing
2.3. Sequence Analysis and Comparison
2.4. Plasmid Construction, Transfection and Cell Staining
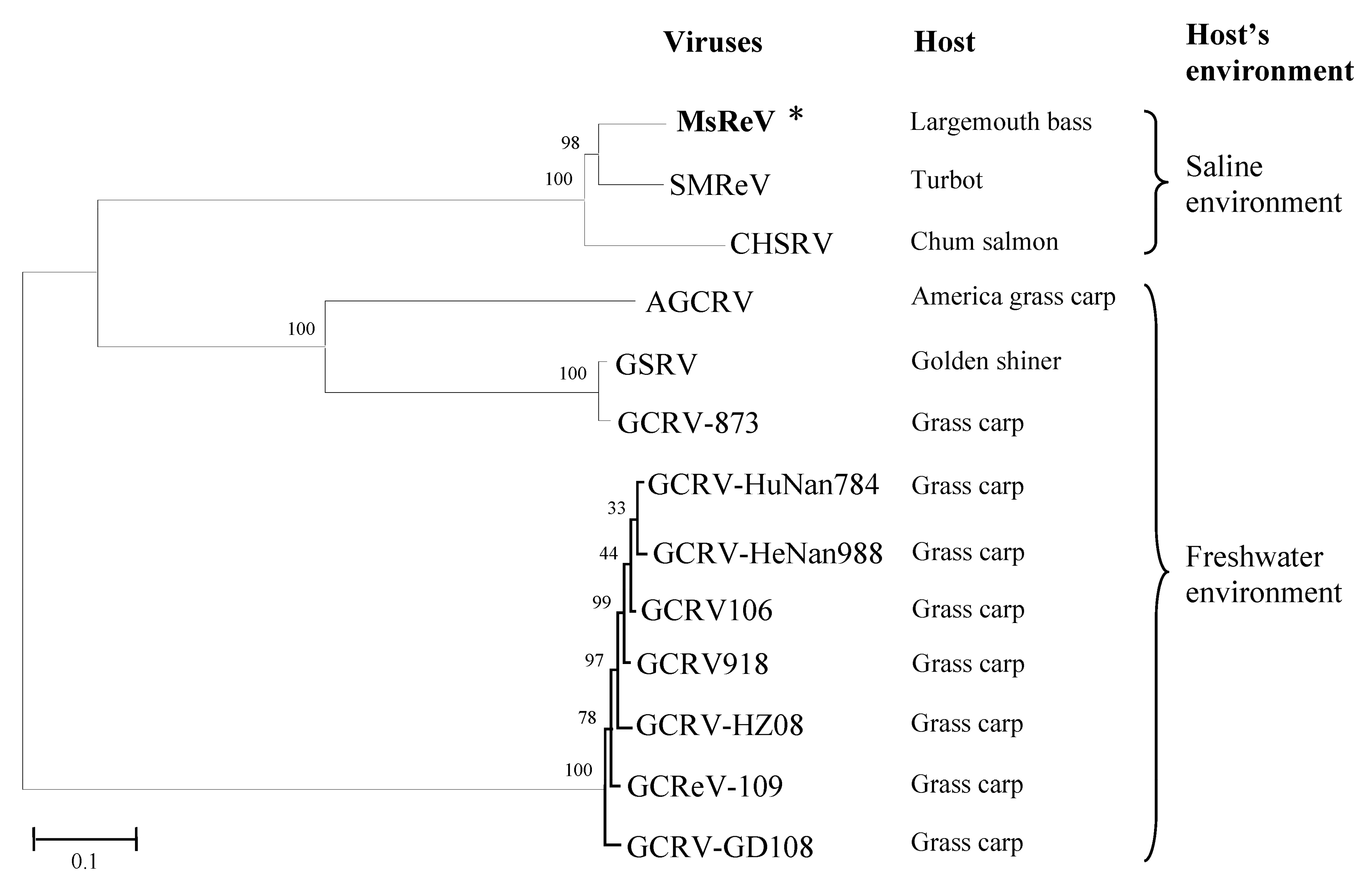
2.5. Phylogenetic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Different Aquareoviruses | Genome Segment/Length (bp) | Coding Segment | S1 | S2 | S3 | S5 | S6 | S8 | S10 | Coding Segment/GenBank acc. No. | Identity (%) of the Concatenated Seven Structural Proteins between MsReV and Other Aquareoviruses | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Structural Protein | VP1 | VP2 | VP3 | VP4 | VP5 | VP6 | VP7 | |||||||||||
| Size (aa) | MW (kDa) | Size (aa) | MW (kDa) | Size (aa) | MW (kDa) | Size (aa) | MW (kDa) | Size (aa) | MW (kDa) | Size (aa) | MW (kDa) | Size (aa) | MW (kDa) | |||||
| MsReV | S1/3947 S2/3866 S3/3687 S4/2622 S5/2242 S6/2056 S7/1399 S8/1317 S9/1118 S10/987 S11/783 | 1297 | 140.91 | 1274 | 141.24 | 1209 | 131.05 | 722 | 79.82 | 653 | 69.04 | 417 | 45.39 | 298 | 32.38 | S1/KJ740725 S2/KJ740726 S3/KJ740727 S5/KJ740729 S6/KJ740730 S8/KJ740732 S10/KJ740734 | 100 | |
| SMReV | S1/3947 S2/3866 S3/3687 S4/2640 S5/2241 S6/2057 S7/1399 S8/1317 S9/1118 S10/986 S11/784 | 1297 | 141.40 | 1274 | 140.97 | 1209 | 131.10 | 730 | 80.52 | 653 | 69.25 | 417 | 45.18 | 298 | 32.18 | S1/HM989930 S2/HM989931 S3/HM989932 S5/HM989934 S6/HM989935 S8/HM989937 S10/HM989939 | 91.2 | |
| CHSRV | S1/3947 S2/3867 S3/3690 S4/partial S5/2242 S6/2052 S7/1395 S8/1317 S9/1118 S10/985 S11/783 | 1297 | 140.93 | 1240 | 137.58 | 1210 | 131.95 | 723 | 80.15 | 643 | 68.89 | 417 | 45.34 | 298 | 32.41 | S1/AF418294 S2/AF418295 S3/AF418296 S5/AF418298 S6/AF418299 S8/AF418301 S10/AF418303 | 85.5 | |
| S5 | S6 | |||||||||||||||||
| VP5 | VP4 | |||||||||||||||||
| Size (aa) | MW (kDa) | Size (aa) | MW (kDa) | |||||||||||||||
| GCRV-873 | S1/3949 S2/3877 S3/3702 S4/2320 S5/2239 S6/2039 S7/1414 S8/1296 S9/1130 S10/909 S11/820 | 1299 | 141.41 | 1274 | 141.54 | 1214 | 132.10 | 728 | 80.24 | 648 | 68.60 | 412 | 44.58 | 276 | 29.81 | S1/AF260511 S2/AF260512 S3/AF260513 S5/AF403391 S6/AF403392 S8/AF403394 S10/AF403396 | 48.5 | |
| GSRV | S1/3949 S2/3877 S3/3702 S4/2320 S5/2239 S6/2039 S7/1414 S8/1297 S9/1130 S10/909 S11/820 | 1299 | 141.27 | 1274 | 141.59 | 1214 | 132.06 | 728 | 80.25 | 648 | 68.56 | 412 | 44.59 | 276 | 29.79 | S1/NC_005166 S2/NC_005167 S3/NC_005168 S5/NC_005170 S6/NC_005171 S8/NC_005173 S10/NC_005175 | 48.4 | |
| AGCRV | S1/3949 S2/3876 S3/3709 S4/2293 S5/2237 S6/2042 S7/1356 S8/1305 S9/1125 S10/912 S11/772 | 1289 | 141.11 | 1274 | 141.98 | 1215 | 131.93 | 728 | 80.14 | 650 | 68.99 | 413 | 44.97 | 273 | 30.36 | S1/NC_010584 S2/NC_010585 S3/NC_010586 S5/NC_010588 S6/NC_010589 S8/NC_010591 S10/NC_010593 | 47.8 | |
| S9 | S11 | |||||||||||||||||
| VP6 | VP7 | |||||||||||||||||
| Size (aa) | MW (kDa) | Size (aa) | MW (kDa) | |||||||||||||||
| GCRV106 | S1/3927 S2/3867 S3/3753 S4/2263 S5/2229 S6/2030 S7/1604 S8/1556 S9/1320 S10/1124 S11/1105 | 1294 | 143.68 | 1273 | 142.64 | 1232 | 135.82 | 726 | 80.61 | 650 | 68.37 | 418 | 48.00 | 310 | 35.46 | S1/KC201166 S2/KC201167 S3/KC201168 S5/KC201170 S6/KC201171 S9/KC201174 S11/KC201176 | 30.7 | |
| GCReV-109 | S1/3928 S2/3867 S3/3753 S4/2263 S5/2230 S6/2028 S7/1604 S8/1560 S9/1320 S10/1124 S11/1027 | 1294 | 143.72 | 1273 | 142.08 | 1232 | 135.73 | 726 | 80.64 | 650 | 68.21 | 418 | 47.96 | 310 | 35.48 | S1/KF712475 S2/KF712476 S3/KF712477 S5/KF712479 S6/KF712480 S9/KF712483 S11/KF712485 | 30.6 | |
| GCRV918 | S1/3927 S2/3867 S3/3753 S4/2263 S5/2229 S6/2030 S7/1604 S8/1556 S9/1320 S10/1124 S11/1107 | 1294 | 143.61 | 1273 | 142.64 | 1232 | 135.85 | 726 | 80.58 | 650 | 68.29 | 418 | 48.09 | 310 | 35.48 | S1/KC201177 S2/KC201178 S3/KC201179 S5/KC201181 S6/KC201182 S9/KC201185 S11/KC201187 | 30.6 | |
| GCRV-GD108 | S1/3928 S2/3867 S3/3752 S4/2263 S5/2230 S6/2028 S7/1604 S8/1560 S9/1320 S10/1124 S11/1027 | 1294 | 143.43 | 1273 | 142.61 | 1232 | 135.78 | 726 | 80.72 | 650 | 68.28 | 418 | 47.99 | 310 | 35.43 | S1/HQ231198 S2/HQ231199 S3/HQ231200 S5/HQ231202 S6/HQ231208 S9/HQ231205 S11/HQ231207 | 30.6 | |
| GCRV-HuNan794 | S1/3927 S2/3867 S3/3753 S4/2263 S5/2229 S6/2030 S7/1604 S8/1556 S9/1320 S10/1124 S11/1107 | 1294 | 143.68 | 1273 | 142.62 | 1232 | 135.66 | 726 | 80.65 | 650 | 68.37 | 418 | 48.03 | 310 | 35.43 | S1/KC238676 S2/KC238677 S3/KC238678 S5/KC238680 S6/KC238681 S9/KC238684 S11/KC238686 | 30.6 | |
| GCRV-HeNan988 | S1/3927 S2/3867 S3/3753 S4/2263 S5/2229 S6/2030 S7/1604 S8/1556 S9/1320 S10/1124 S11/1107 | 1294 | 143.72 | 1273 | 142.66 | 1232 | 135.45 | 726 | 80.62 | 650 | 68.37 | 418 | 48.10 | 310 | 35.48 | S1/KC847320 S2/KC847321 S3/KC847322 S5/KC847324 S6/KC847325 S9/KC847328 S11/KC847330 | 30.6 | |
| GCRV-HZ08 | S1/3927 S2/3870 S3/3753 S4/2263 S5/2229 S6/2030 S7/1604 S8/1560 S9/1320 S10/1124 S11/1027 | 1294 | 143.66 | 1273 | 143.10 | 1232 | 135.86 | 726 | 80.54 | 650 | 68.37 | 418 | 47.87 | 310 | 35.44 | S1/GQ896334 2/GQ896335 S3/GU350742 S5/GQ896336 S6/GQ896337 S9/GU350746 S11/GU350748 | 30.4 | |
3. Results
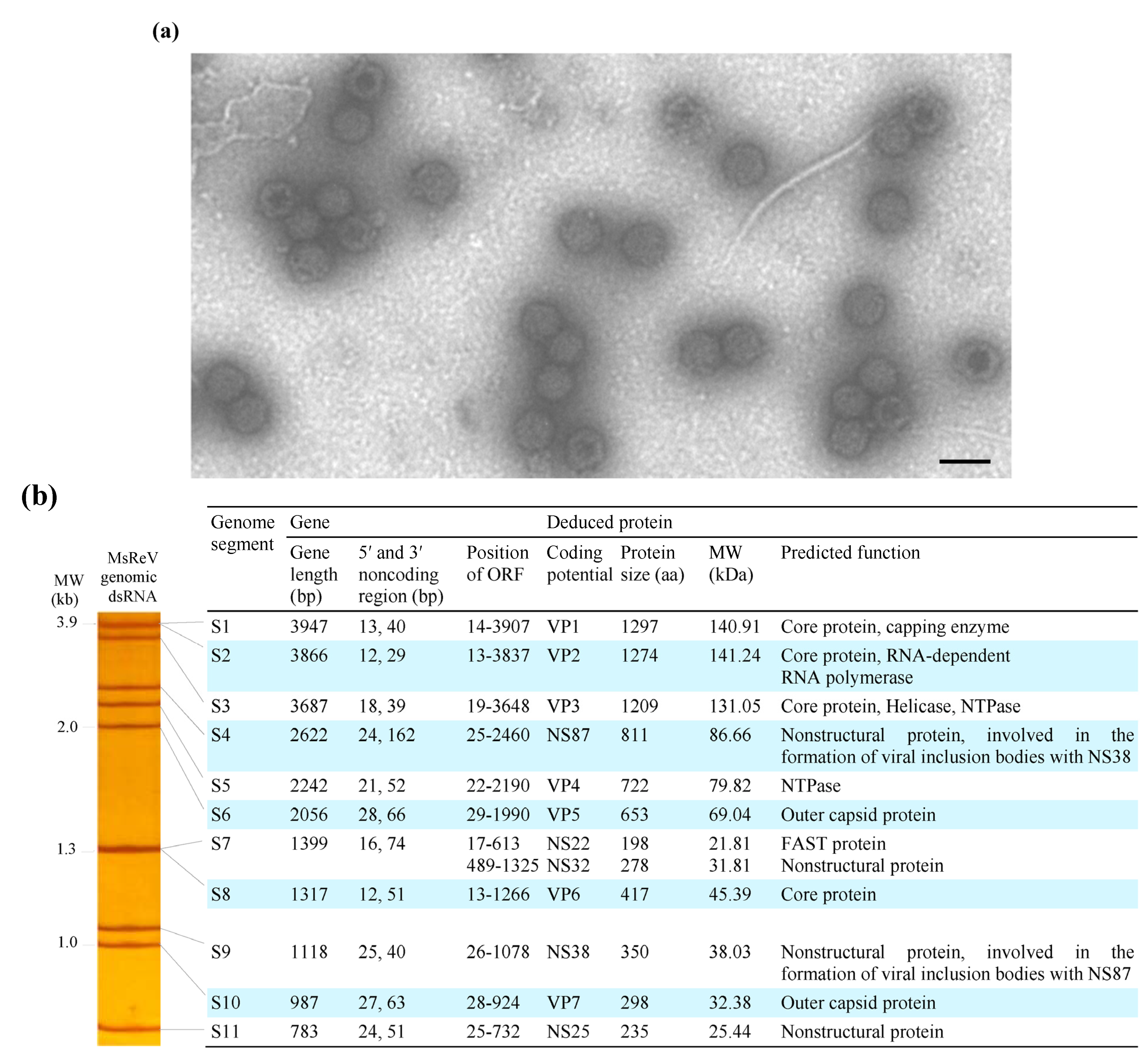
3.1. Identification of MsReV and Electrophoretic Pattern of Viral Genome Segments

3.2. Organization and Structure of MsReV Genome
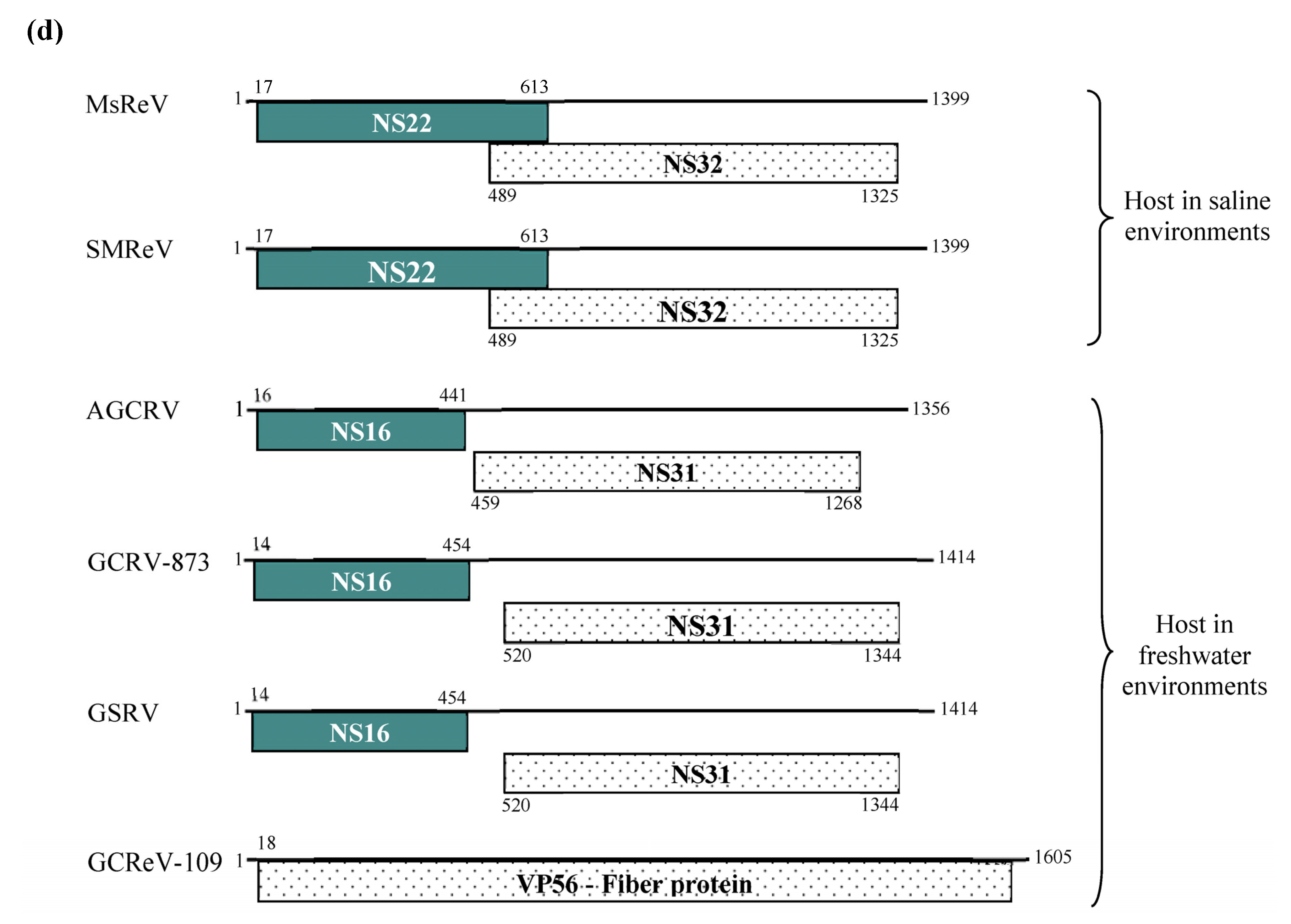
3.3. Comparison of the S4 Segments and Encoded Nonstructural Proteins of Three Aquareoviruses

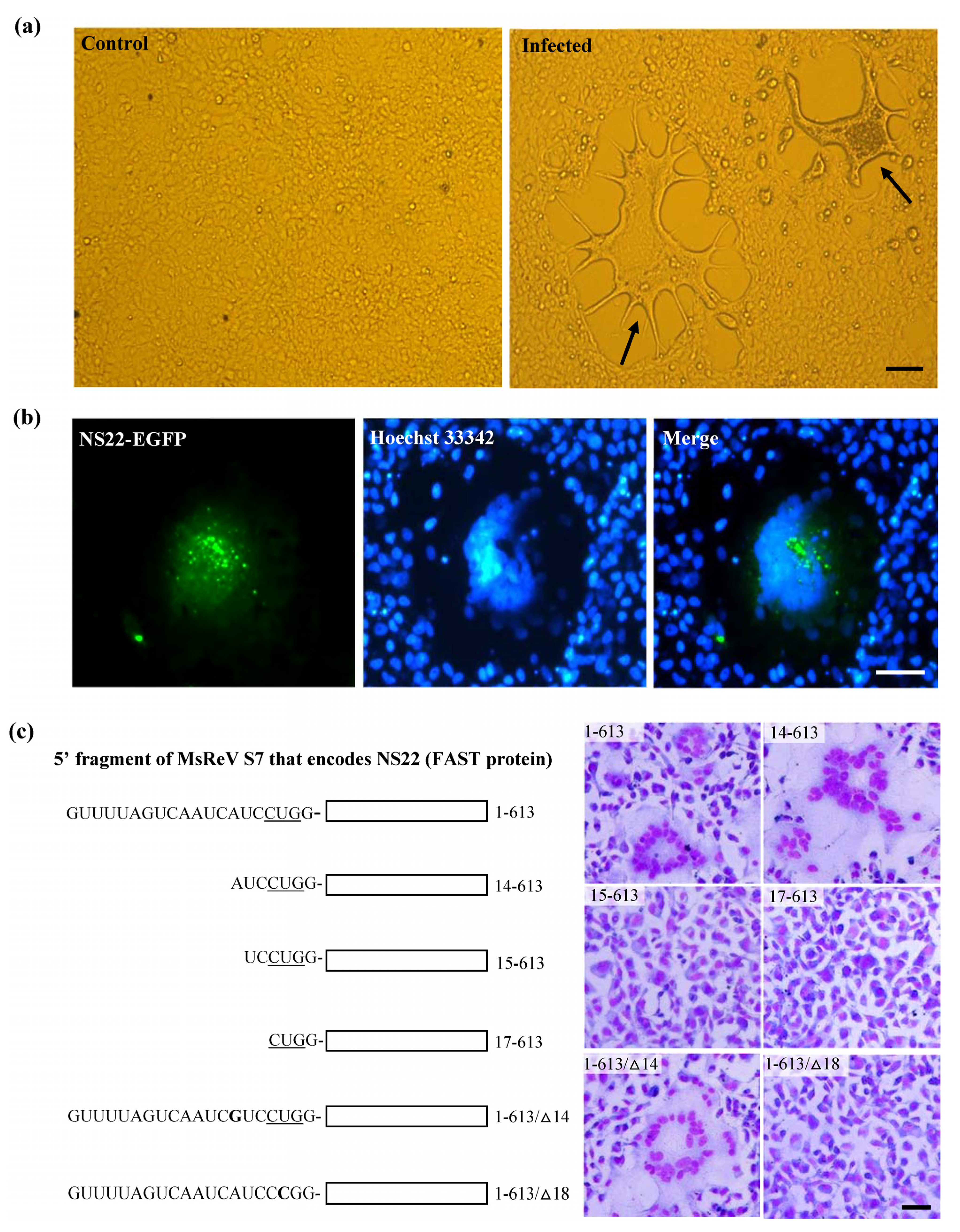
3.4. Experimental Verification of the Function of NS22 Protein Encoded by MsReV S7


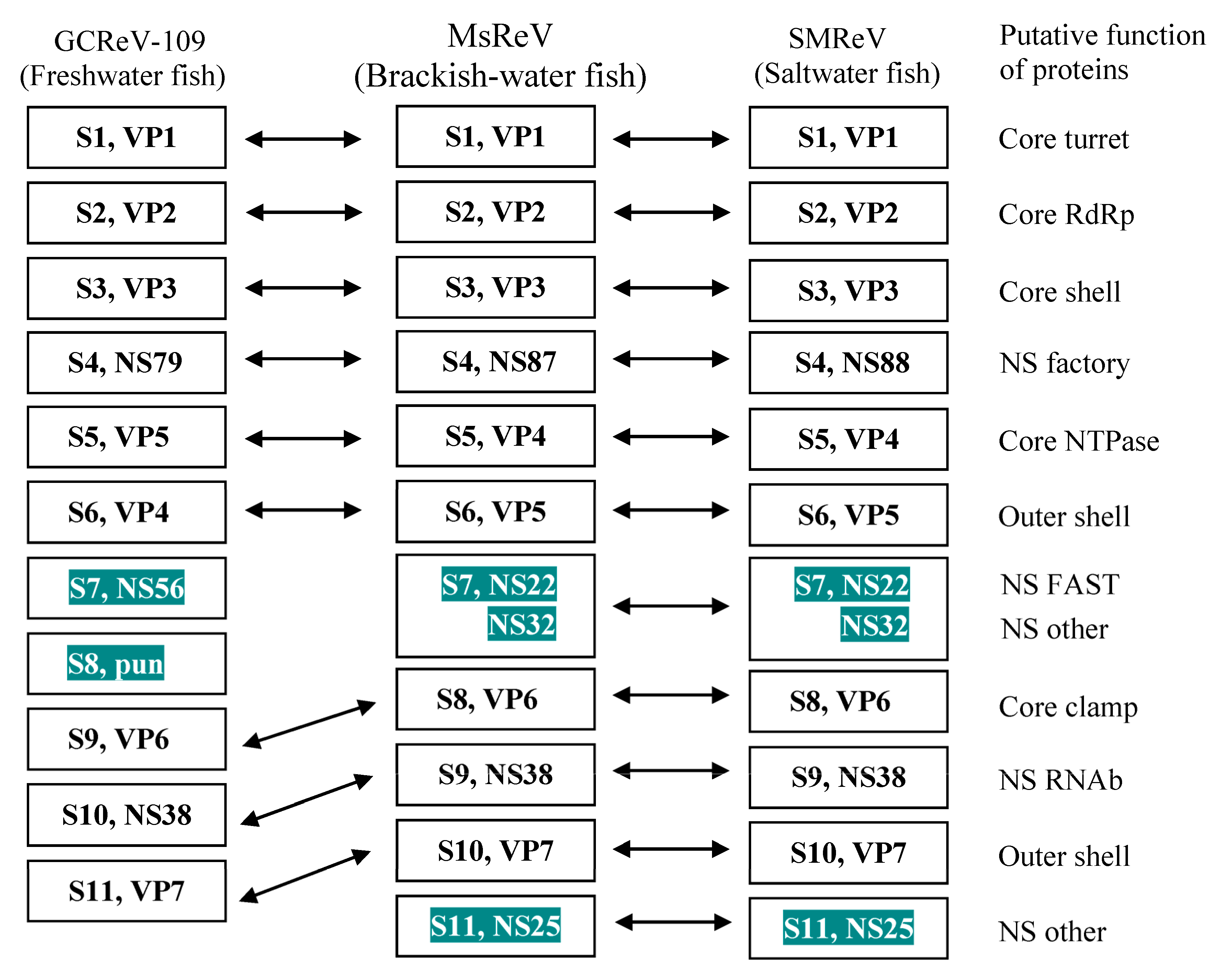
3.5. MsReV and SMReV Are Closely Related to Equivalent Genome Segments

3.6. MsReV Is More Closely Related to SMReV than to GCRV-109

4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Auguste, A.J.; Kaelber, J.T.; Fokam, E.B.; Guzman, H.; Carrington, C.V.; Erasmus, J.H.; Kamgang, B.; Popov, V.L.; Jakana, J.; Liu, X.; et al. A newly isolated reovirus has the simplest genomic and structural organization of any reovirus. J. Virol. 2015, 89, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Mertens, P.P.C.; Becnel, J.; Belaganahalli, S.; Bergoin, M.; Brussaard, C.P.; Chappell, J.D.; Ciarlet, M.; del Vas, M.; Dermody, T.S.; et al. Family Reoviridae. In Virus Taxonomy-Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: London, UK, 2012; pp. 541–637. [Google Scholar]
- Gui, J.F.; Zhu, Z.Y. Molecular basis and genetic improvement of economically important traits in aquaculture animals. Chin. Sci. Bull. 2012, 57, 1751–1760. [Google Scholar] [CrossRef]
- Mohd Jaafar, F.; Goodwin, A.E.; Belhouchet, M.; Merry, G.; Fang, Q.; Cantaloube, J.F.; Biagini, P.; de Micco, P.; Mertens, P.P.; Attoui, H. Complete characterisation of the American grass carp reovirus genome (genus Aquareovirus: Family Reoviridae) reveals an evolutionary link between aquareoviruses and coltiviruses. Virology 2008, 373, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Lovoll, M.; Tengs, T.; Hornig, M.; Hutchison, S.; Hui, J.; Kongtorp, R.T.; Savji, N.; Bussetti, A.V.; Solovyov, A. Heart and skeletal muscle inflammation of farmed salmon is associated with infection with a novel reovirus. PLoS ONE 2010, 5, e11487. [Google Scholar] [CrossRef] [PubMed]
- Racine, T.; Hurst, T.; Barry, C.; Shou, J.; Kibenge, F.; Duncan, R. Aquareovirus effects syncytiogenesis by using a novel member of the FAST protein family translated from a noncanonical translation start site. J. Virol. 2009, 83, 5951–5925. [Google Scholar] [CrossRef] [PubMed]
- Rangel, A.A.C.; Rockemann, D.D.; Hetrick, F.M.; Samal, S.K. Identification of grass carp haemorrhage virus as a new genogroup of aquareovirus. J. Gen. Virol. 1999, 80, 2399–2402. [Google Scholar] [PubMed]
- Schachner, O.; Soliman, H.; Straif, M.; Schilcher, F.; El-Matbouli, M. Isolation and characterization of a novel reovirus from white bream Blicca bjoerkna. Dis. Aquat. Organ. 2014, 112, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Seng, E.K.; Fang, Q.; Chang, S.F.; Ngoh, G.H.; Qin, Q.W.; Lam, T.J.; Sin, Y.M. Characterisation of a pathogenic virus isolated from marine threadfin fish (Eleutheronema tetradactylus) during a disease outbreak. Aquaculture 2002, 214, 1–18. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Ruan, H.M.; Li, Z.Q.; Zhang, J.; Gui, J.F. Detection of grass carp hemorrhage virus (GCHV) from Vietnam and comparison with GCHV strain from China. Chin. High Technol. Lett. 2003, 9, 7–13. [Google Scholar]
- Zhang, Q.Y.; Gui, J.F. Atlas of Aquatic Viruses and Viral Diseases; Science Press: Beijing, China, 2012. [Google Scholar]
- Zhang, Q.Y.; Gui, J.F. Virus genomes and virus-host interactions in aquaculture animals. Sci. China Life Sci. 2015, 58, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Key, T.; Read, J.; Nibert, M.L.; Duncan, R. Piscine reovirus encodes a cytotoxic, non-fusogenic, integral membrane protein and previously unrecognized virion outer-capsid proteins. J. Gen. Virol. 2013, 94, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Gui, J.F.; Gao, X.C.; Pei, C.; Hong, Y.J.; Zhang, Q.Y. Genome architecture changes and major gene variations of Andrias davidianus ranavirus (ADRV). Vet. Res. 2013, 44, 101. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.Y.; Ou, T.; Zhu, R.L.; Zhang, Q.Y. Sequencing and analysis of the complete genome of Rana grylio virus (RGV). Arch. Virol. 2012, 157, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Nibert, M.L.; Duncan, R. Bioinformatics of recent aqua- and orthoreovirus isolates from fish: Evolutionary gain or loss of FAST and fiber proteins and taxonomic implications. PLoS ONE 2013, 8, e68607. [Google Scholar] [CrossRef] [PubMed]
- Pepin, K.M.; Lass, S.; Pulliam, J.R.; Read, A.F.; Lloyd-Smith, J.O. Identifying genetic markers of adaptation for surveillance of viral host jumps. Nat. Rev. Microbiol. 2010, 8, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.Y.; Xiao, F.; Xie, J.; Li, Z.Q.; Gui, J.F. Complete genome sequence of lymphocystis disease virus isolated from China. J. Virol. 2004, 78, 6982–6994. [Google Scholar] [CrossRef] [PubMed]
- Eusebio-Cope, A.; Sun, L.; Hillman, B.I.; Suzuki, N. Mycoreovirus 1 S4-coded protein is dispensable for viral replication but necessary for efficient vertical transmission and normal symptom induction. Virology 2010, 397, 399–408. [Google Scholar] [PubMed]
- Shmulevitz, M.; Gujar, S.A.; Ahn, D.G.; Mohamed, A.; Lee, P.W. Reovirus variants with mutations in genome segments S1 and L2 exhibit enhanced virion infectivity and superior oncolysis. J. Virol. 2012, 86, 7403–7413. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.X.; Lü, L.; Ou, Y.J.; Su, H.J.; Li, G.; Guo, Z.X.; Zhang, R.; Zheng, P.R.; Chen, Y.G.; He, J.G.; et al. Sequence analysis of 12 genome segments of mud crab reovirus (MCRV). Virology 2012, 422, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Eaton, H.E.; Ring, B.A.; Brunetti, C.R. The genomic diversity and phylogenetic relationship in the family iridoviridae. Viruses 2010, 2, 1458–1475. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Fang, Q.; Mohd Jaafar, F.; Cantaloube, J.F.; Biagini, P.; de Micco, P.; de Lamballerie, X. Common evolutionary origin of aquareoviruses and orthoreoviruses revealed by genome characterization of Golden shiner reovirus, Grass carp reovirus, Striped bass reovirus and golden ide reovirus (genus Aquareovirus, family Reoviridae). J. Gen. Virol. 2002, 83, 1941–1951. [Google Scholar] [PubMed]
- Pei, C.; Ke, F.; Chen, Z.Y.; Zhang, Q.Y. Complete genome sequence and comparative analysis of grass carp reovirus strain 109 (GCReV-109) with other grass carp reovirus strains reveals no significant correlate regional distribution. Arch. Virol. 2014, 159, 2435–2440. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zeng, W.W.; Liu, C.; Zhang, C.; Wang, Y.Y.; Shi, C.B.; Wu, S.Q. Complete genome sequence of a reovirus isolated from grass carp, indicating different genotypes of GCRV in China. J. Virol. 2012, 86, 12466. [Google Scholar] [CrossRef] [PubMed]
- Ke, F.; He, L.B.; Pei, C.; Zhang, Q.Y. Turbot reovirus (SMReV) genome encoding a FAST protein with a non-AUG start site. BMC Genomics 2011, 12, e323. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.D.; Rao, S.J.; Zeng, L.B.; Ma, J.; Zhou, Y.; Xu, J.; Zhang, H. Identification and genomic characterization of a novel fish reovirus, Hubei grass carp disease reovirus, isolated in 2009 in China. J. Gen. Virol. 2013, 94, 2266–2277. [Google Scholar] [CrossRef] [PubMed]
- Blindheim, S.; Nylund, A.; Watanabe, K.; Plarre, H.; Erstad, B.; Nylund, S. A new aquareovirus causing high mortality in farmed Atlantic halibut fry in Norway. Arch. Virol. 2015, 160, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Kibenge, M.J.; Iwamoto, T.; Wang, Y.; Morton, A.; Godoy, M.G.; Kibenge, F.S. Whole-genome analysis of piscine reovirus (PRV) shows PRV represents a new genus in family Reoviridae and its genome segment S1 sequences group it into two separate sub-genotypes. Virol. J. 2013, 10, e230. [Google Scholar] [CrossRef] [PubMed]
- Wellehan, J.F., Jr.; Childress, A.L.; Marschang, R.E.; Johnson, A.J.; Lamirande, E.W.; Roberts, J.F.; Vickers, M.L.; Gaskin, J.M.; Jacobson, E.R. Consensus nested PCR amplification and sequencing of diverse reptilian, avian, and mammalian orthoreoviruses. Vet. Microbiol. 2009, 133, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Ke, F.; He, L.B.; Zhang, Q.Y. Nonstructural protein NS80 is crucial in recruiting viral components to form aquareoviral factories. PLoS ONE 2013, 8, e63737. [Google Scholar] [CrossRef] [PubMed]
- Sanjuán, R. From molecular genetics to phylodynamics: Evolutionary relevance of mutation rates across viruses. PLoS Pathog. 2012, 8, e1002685. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Zhu, J.; Hui, W.H.; Zhang, X.; Honig, B.; Fang, Q.; Zhou, Z.H. Backbone model of an aquareovirus virion by cryo-electron microscopy and bioinformatics. J. Mol. Biol. 2010, 397, 852–863. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Tian, Y.Y.; Deng, G.C.; Chi, Y.Y.; Jiang, X.Y. Complete genomic sequence of a reovirus isolated from grass carp in China. Virus Res. 2012, 163, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Sun, X.Y.; Yan, L.M.; Shao, L.; Fang, Q. The NS16 protein of aquareovirus-C is a fusion-associated small transmembrane (FAST) protein, and its activity can be enhanced by the nonstructural protein NS26. Virus Res. 2013, 171, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jin, L.; Fang, Q.; Hui, W.H.; Zhou, Z.H. 3.3 Å cryo-EM structure of a nonenveloped virus reveals a priming mechanism for cell entry. Cell 2010, 141, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- He, L.B.; Ke, F.; Wang, J.; Gao, X.C.; Zhang, Q.Y. Rana grylio virus (RGV) envelope protein 2L: Subcellular localization and essential roles in virus infectivity revealed by conditional lethal mutant. J. Gen. Virol. 2014, 95, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Ke, F.; Huang, Y.H.; Zhao, J.G.; Gui, J.F.; Zhang, Q.Y. Identification and characterization of a novel envelope protein in Rana grylio virus. J. Gen. Virol. 2008, 89, 1866–1872. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Winton, J.R.; Lannan, C.N.; Fryer, J.L.; Kimura, T. Isolation of a new reovirus from Chum salmon in Japan. Fish Pathol. 1981, 15, 155–162. [Google Scholar] [CrossRef]
- Brown, C.W.; Stephenson, K.B.; Hanson, S.; Kucharczyk, M.; Duncan, R.; Bell, J.C.; Lichty, B.D. The p14 FAST protein of reptilian reovirus increases vesicular stomatitis virus neuropathogenesis. J. Virol. 2009, 83, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Key, T.; Duncan, R. A compact, multifunctional fusion module directs cholesterol-dependent homomultimerization and syncytiogenic efficiency of reovirus p10 FAST proteins. PLoS Pathog. 2014, 10, e1004023. [Google Scholar] [CrossRef] [PubMed]
- Salsman, J.; Top, D.; Boutilier, J.; Duncan, R. Extensive syncytium formation mediated by the reovirus FAST proteins triggers apoptosis-induced membrane instability. J. Virol. 2005, 79, 8090–8100. [Google Scholar] [CrossRef] [PubMed]
- Markussen, T.; Dahle, M.K.; Tengs, T.; Løvoll, M.; Finstad, Ø.W.; Wiik-Nielsen, C.R.; Grove, S.; Lauksund, S.; Robertsen, B.; Rimstad, E. Sequence analysis of the genome of piscine orthoreovirus (PRV) associated with heart and skeletal muscle inflammation (HSMI) in Atlantic salmon (Salmo salar). PLoS ONE 2013, 8, e70075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, C.R.; Fletcher, N.F. Emerging virus diseases: Can we ever expect the unexpected? Emerg Microbes Infect. 2012, 1, e46. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–994. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Holland, J.J. RNA virus mutations and fitness for survival. Annu. Rev. Microbiol. 1997, 51, 151–178. [Google Scholar] [CrossRef] [PubMed]
- Lauring, A.S.; Frydman, J.; Andino, R. The role of mutational robustness in RNA virus evolution. Nat. Rev. Microbiol. 2013, 11, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Simon-Loriere, E.; Holmes, E.C. Why do RNA viruses recombine? Nat. Rev. Microbiol. 2011, 9, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Muñoz, S.L.; Tenaillon, O.; Goldhill, D.; Brao, K.; Turner, P.E.; Chao, L. Electrophoretic mobility confirms reassortment bias among geographic isolates of segmented RNA phages. BMC Evol. Biol. 2013, 13, e206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavazzi, C.; Yver, M.; Isel, C.; Smyth, R.P.; Rosa-Calatrava, M.; Lina, B.; Moulès, V.; Marquet, R. A functional sequence-specific interaction between influenza A virus genomic RNA segments. Proc. Natl. Acad. Sci. USA 2013, 110, 16604–16609. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.-Y.; Gao, X.-C.; Zhang, Q.-Y. Whole-Genome Analysis of a Novel Fish Reovirus (MsReV) Discloses Aquareovirus Genomic Structure Relationship with Host in Saline Environments. Viruses 2015, 7, 4282-4302. https://0-doi-org.brum.beds.ac.uk/10.3390/v7082820
Chen Z-Y, Gao X-C, Zhang Q-Y. Whole-Genome Analysis of a Novel Fish Reovirus (MsReV) Discloses Aquareovirus Genomic Structure Relationship with Host in Saline Environments. Viruses. 2015; 7(8):4282-4302. https://0-doi-org.brum.beds.ac.uk/10.3390/v7082820
Chicago/Turabian StyleChen, Zhong-Yuan, Xiao-Chan Gao, and Qi-Ya Zhang. 2015. "Whole-Genome Analysis of a Novel Fish Reovirus (MsReV) Discloses Aquareovirus Genomic Structure Relationship with Host in Saline Environments" Viruses 7, no. 8: 4282-4302. https://0-doi-org.brum.beds.ac.uk/10.3390/v7082820