
Molecular Characterization of a Trisegmented Mycovirus from the Plant Pathogenic Fungus Colletotrichum gloeosporioides

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolate and Culture Conditions
2.2. Detection and Purification of dsRNA
2.3. cDNA Cloning, Sequencing, and Phylogenetic Analysis
2.4. Preparation of Virus Particles
2.5. DNA Isolation and PCR Amplification
3. Results
3.1. Three dsRNA Segments in Strain HZ-1 Are Associated with a Mycovirus
3.2. Molecular Cloning of dsRNAs Associated with Strain HZ-1
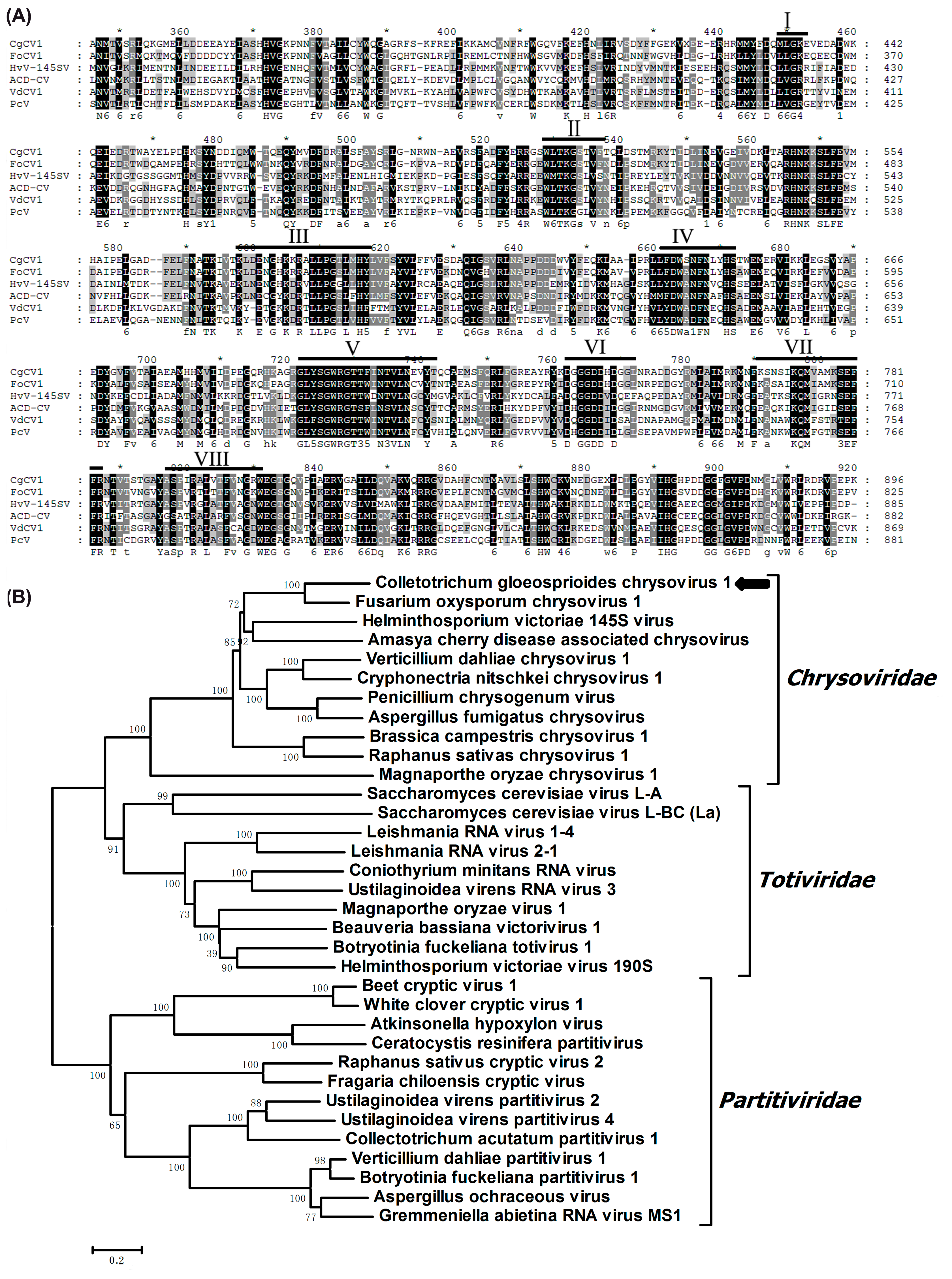
3.3. Molecular Analysis of the Three dsRNA Segments
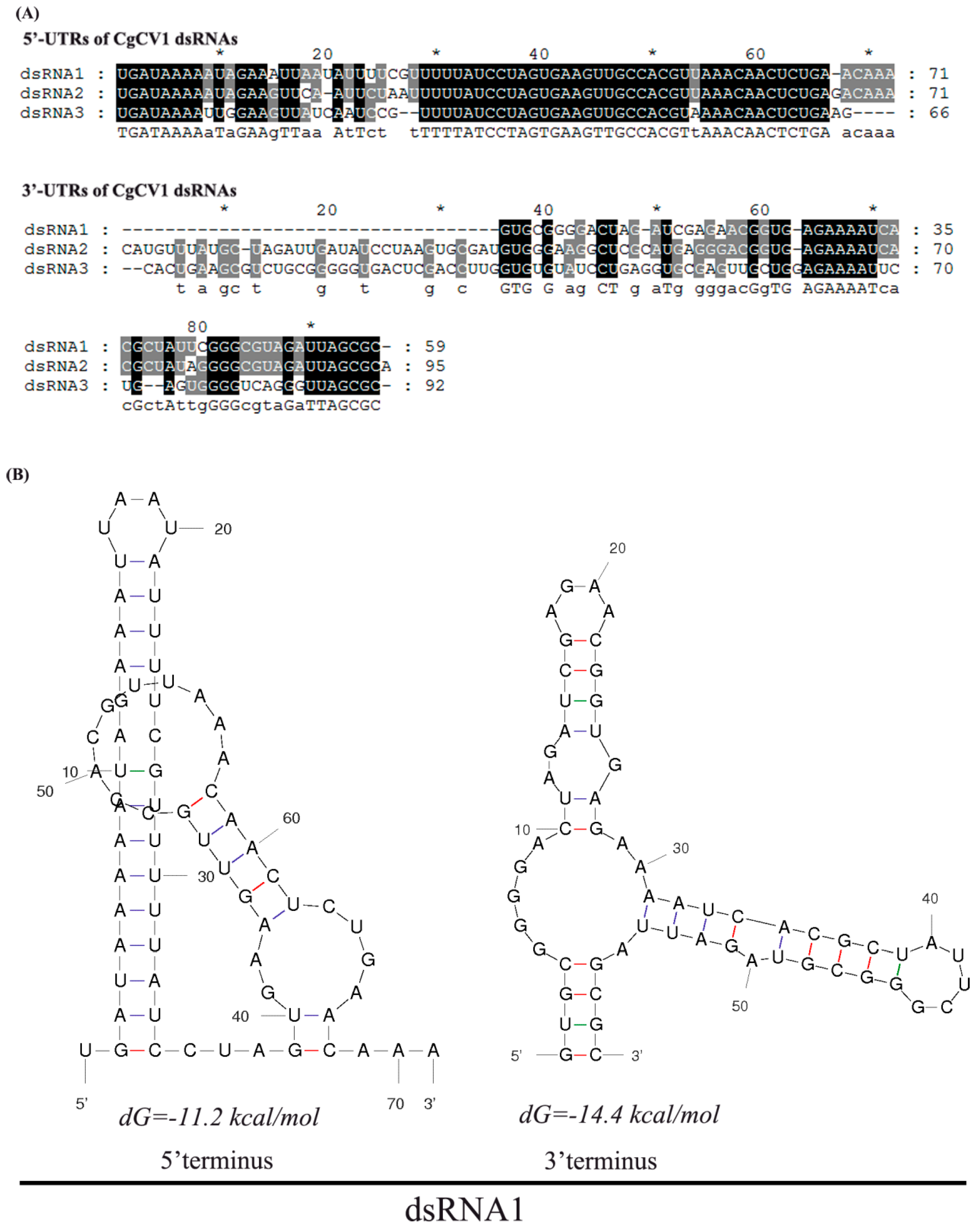
3.4. 5′- and 3′-Untranslated Regions (UTRs)
3.5. Searching for CgCV1-Like Sequences in Extant RNA Viral Sequences and Eukaryotic Nuclear Genomes
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant. Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.T.; Jiang, D.H. New Insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [PubMed]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Castón, J.R.; Flores, R.; Hillman, B.I.; Jiang, D. Chrysoviridae. In Virus Taxonomy 8th Report of the ICTV; EC 47: London, UK, 2005. [Google Scholar]
- Jiang, D.; Ghabrial, S.A. Molecular characterization of Penicillium chrysogenum virus: Reconsideration of the taxonomy of the genus Chrysovirus. J. Gen. Virol. 2004, 85, 2111–2121. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, J.; Xu, A.; Wang, T.; Chen, J.; Zhu, X. Molecular characterization of a trisegmented chrysovirus isolated from the radish Raphanus sativus. Virus Res. 2013, 176, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, J.O. Greasy spot and greasy spot rind blotch. In Compendium of Citrus Diseases; Whiteside, J.O., Garnsey, S.M., Timmer, L.W., Eds.; APS Press: St. Paul, MN, USA, 1988. [Google Scholar]
- Agostini, J.P.; Timmer, L.W.; Mitchell, D.J. Morphological and pathological characteristics of strains of Colletotrichum gloeosporioides from citrus. Phytopathology 1992, 82, 1377–1382. [Google Scholar] [CrossRef]
- Lima, W.G.; Spósito, M.B.; Amorim, L.; Gonçalves, F.P.; de Filho, P.A.M. Colletotrichum gloeosporioides, a new causal agent of citrus post-bloom fruit drop. Eur. J. Plant Pathol. 2011, 131, 157–165. [Google Scholar] [CrossRef]
- McGovern, R.J.; Seijo, T.E.; Hendricks, K.; Roberts, P.D. New report of Colletotrichum gloeosporioides causing postbloom fruit drop on citrus in Bermuda. Can. J. Plant Pathol. 2012, 34, 187–194. [Google Scholar] [CrossRef]
- Liao, C.Y.; Chen, M.Y.; Chen, Y.K.; Wang, T.C.; Sheu, Z.M.; Kuo, K.C.; Lee, M.H. Characterization of three Colletotrichum acutatum isolates from Capsicum spp. Eur. J. Plant Pathol. 2012, 133, 599–608. [Google Scholar] [CrossRef]
- Figueirêdo, L.C.D.; Figueirêdo, G.S.D.; Giancoli, Á.C.; Tanaka, F.A.; da Silva, L.A.; Kitajima, E.W.; Azevedo, J.L. Detection of isometric, dsRNA-containing viral particles in Colletotrichum gloeosporioides isolated from cashew tree. Trop. Plant Pathol. 2012, 37, 142–145. [Google Scholar] [CrossRef]
- Zhon, J.; Chen, D.; Lei, X.H.; Zhu, H.J.; Zhu, J.Z.; Gao, B.D. Detection and characterization of a novel Gammapartitivirus in the phytopathogenic fungus Colletotrichum acutatum strain HNZJ001. Virus Res. 2014, 190, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Rosseto, P.; Costa, A.T.; Polonio, J.C.; da Silva, A.A.; Pamphile, J.A.; Azevedo, J.L. Investigation of mycoviruses in endophytic and phytopathogenic strains of Colletotrichum from different hosts. Genet. Mol. Res. GMR 2016, 15, 15017651. [Google Scholar] [CrossRef] [PubMed]
- Campo, S.; Gilbert, K.B.; Carrington, J.C. Small RNA-Based Antiviral Defense in the Phytopathogenic Fungus Colletotrichum higginsianum. PLoS Pathog. 2016, 12, e1005640. [Google Scholar] [CrossRef] [PubMed]
- Morris, T.J.; Dodds, J.A. Isolation and analysis of double-stranded RNA from virus-infected plant and fungal tissue. Phytopathology 1979, 69, 854–858. [Google Scholar] [CrossRef]
- Cao, Y.F.; Zhu, X.W.; Xiang, Y.; Li, D.Q.; Yang, J.R.; Mao, Q.Z.; Chen, J.S. Genomic characterization of a novel dsRNA virus detected in the phytopathogenic fungus Verticillium dahliae Kleb. Virus Res. 2011, 159, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Rong, R.; Rao, S.; Scott, S.W.; Carner, G.R.; Tainter, F.H. Complete sequence of the genome of two dsRNA viruses from Discula destructiva. Virus Res. 2002, 90, 217–224. [Google Scholar] [CrossRef]
- Potgieter, A.C.; Steele, A.D.; Van Dijk, A.A. Cloning of complete genome sets of six dsRNA viruses using an improved cloning method for large dsRNA genes. J. Gen. Virol. 2002, 83, 2215–2223. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, J.S.; Liu, L.; Yu, X.; Yu, S.; Fu, T.Z.; Liu, W.H. Complete nucleotide sequences and genome characterization of double-stranded RNA 1 and RNA 2 in the Raphanus sativus-root cv. Yidianhong [corrected]. Arch. Virol. 2006, 151, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Arch. Virol. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic. Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A laboratory Manual; Sambrook, J., Fritsch, E.F., Maniatis, T., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Bruenn, J. Genes from Double-Stranded RNA Viruses in the Nuclear Genomes of Fungi. In Biocommunication of Fungi; Springer: Dordrecht, The Netherlands, 2012; pp. 71–83. [Google Scholar]
- Liu, H.; Fu, Y.; Jiang, D.; Li, G.; Xie, J.; Cheng, J.; Peng, Y.; Ghabrial, S.A.; Yi, X. Widespread horizontal gene transfer from double-stranded RNA viruses to eukaryotic nuclear genomes. J. Virol. 2010, 84, 11876–11887. [Google Scholar] [CrossRef]
- Covelli, L.; Coutts, R.H.A.; Serio, F.D.; Citir, A.; Ac¸ıkgöz, S.; Hernández, C.; Ragozzino, A.; Flores, R. Cherry chlorotic rusty spot and Amasya cherry diseases are associated with a complex pattern of mycoviral-like double-stranded RNAs. I. Characterization of a new species in the genus Chrysovirus. J. Gen. Virol. 2004, 85, 3389–3397. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Aravind, L.; Koonin, E.V. A novel superfamily of predicted cysteine proteases from eukaryotes, viruses and Chlamydia pneumoniae. Trends. Biochem. Sci. 2000, 25, 50–52. [Google Scholar] [CrossRef]
- Jamal, A.; Bignell, E.M.; Coutts, R.H. Complete nucleotide sequences of four dsRNAs associated with a new chrysovirus infecting Aspergillus fumigatus. Virus Res. 2010, 153, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Frias-Staheli, N.; Giannakopoulos, N.V.; Kikkert, M.; Taylor, S.L.; Bridgen, A.; Paragas, J.; Richt, J.A.; Rowland, R.R.; Schmaljohn, C.S.; Lenschow, D.J.; et al. Ovarian tumor domain-containing viral proteases evade ubiquitin- and ISG15-dependent innate immune responses. Cell Host. Microbe 2007, 2, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.Z.; Osaki, H.; Iwanami, T.; Matsumoto, N.; Ohtsu, Y. Molecular characterization of dsRNA segments 2 and 5 and electron microscopy of a novel reovirus from a hypovirulent isolate, W370, of the plant pathogen Rosellinia necatrix. J. Gen. Virol. 2003, 84, 2431–2437. [Google Scholar] [CrossRef] [PubMed]
- Darissa, O.; Willingmann, P.; Schfer, W.; Adam, G. A novel double-stranded RNA mycovirus from Fusarium graminearum: Nucleic acid sequence and genomic structure. Arch. Virol. 2011, 156, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Urayama, S.; Kato, S.; Suzuki, Y.; Aoki, N.; Le, M.T.; Arie, T.; Teraoka, T.; Fukuhara, T.; Moriyama, H. Mycoviruses related to chrysovirus affect vegetative growth in the rice blast fungus Magnaporthe oryzae. J. Gen. Virol. 2010, 91, 3085–3094. [Google Scholar] [CrossRef] [PubMed]
- Urayama, S.; Ohta, T.; Onozuka, N.; Sakoda, H.; Fukuhara, T.; Arie, T.; Teraoka, T.; Moriyama, H. Characterization of Magnaporthe oryzae chrysovirus structural proteins and their expression in Saccharomyces cerevisiae. J. Virol. 2012, 86, 8287–8295. [Google Scholar] [CrossRef] [PubMed]
- Chiba, S.; Kondo, H.; Tani, A.; Saisho, D.; Sakamoto, W.; Kanematsu, S.; Suzuki, N. Widespread endogenization of genome sequences of non-retroviral RNA viruses into plant genomes. PLoS Pathog. 2011, 7, e1002146. [Google Scholar] [CrossRef] [PubMed]




© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, J.; Pang, X.D.; Zhu, H.J.; Gao, B.D.; Huang, W.K.; Zhou, Q. Molecular Characterization of a Trisegmented Mycovirus from the Plant Pathogenic Fungus Colletotrichum gloeosporioides. Viruses 2016, 8, 268. https://0-doi-org.brum.beds.ac.uk/10.3390/v8100268
Zhong J, Pang XD, Zhu HJ, Gao BD, Huang WK, Zhou Q. Molecular Characterization of a Trisegmented Mycovirus from the Plant Pathogenic Fungus Colletotrichum gloeosporioides. Viruses. 2016; 8(10):268. https://0-doi-org.brum.beds.ac.uk/10.3390/v8100268
Chicago/Turabian StyleZhong, Jie, Xi Dan Pang, Hong Jian Zhu, Bi Da Gao, Wen Kun Huang, and Qian Zhou. 2016. "Molecular Characterization of a Trisegmented Mycovirus from the Plant Pathogenic Fungus Colletotrichum gloeosporioides" Viruses 8, no. 10: 268. https://0-doi-org.brum.beds.ac.uk/10.3390/v8100268