
Ubiquitin in Influenza Virus Entry and Innate Immunity


1
Institute of Molecular Life Sciences, University of Zurich, Winterthurerstrasse 190, Zurich CH-8057, Switzerland
2
School of Cellular and Molecular Medicine, University of Bristol, Biomedical Sciences Building, University Walk, Bristol BS8 1TD, UK
3
Structural Biology Research Center, Department of Biological Science, Graduate School of Science, Nagoya University, Furo-cho, Chikusa-ku, Nagoya 464-8602, Japan
*
Author to whom correspondence should be addressed.
Viruses 2016, 8(10), 293; https://0-doi-org.brum.beds.ac.uk/10.3390/v8100293
Submission received: 14 September 2016
/
Revised: 14 October 2016
/
Accepted: 14 October 2016
/
Published: 24 October 2016
(This article belongs to the Special Issue Viruses 2016 - At the Forefront of Virus-Host Interactions)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Viruses are obligatory cellular parasites. Their mission is to enter a host cell, to transfer the viral genome, and to replicate progeny whilst diverting cellular immunity. The role of ubiquitin is to regulate fundamental cellular processes such as endocytosis, protein degradation, and immune signaling. Many viruses including influenza A virus (IAV) usurp ubiquitination and ubiquitin-like modifications to establish infection. In this focused review, we discuss how ubiquitin and unanchored ubiquitin regulate IAV host cell entry, and how histone deacetylase 6 (HDAC6), a cytoplasmic deacetylase with ubiquitin-binding activity, mediates IAV capsid uncoating. We also discuss the roles of ubiquitin in innate immunity and its implications in the IAV life cycle.
1. Ubiquitin and Ubiquitination
Ubiquitin is a small, 8.5 kDa protein composed of 76 amino acids expressed in different tissues and present in different subcellular compartments. Post-translational attachment of ubiquitin to other proteins, namely ubiquitination, alters the function, location, or trafficking of the protein, or targets it for destruction by the 26S proteasome. The ability of ubiquitin to form structurally and functionally distinct polymers greatly increases the complexity of ubiquitination. Ubiquitin has a globular shape with the last four C-terminal residues (LRGG) extending from the compact structure. C-terminal glycine (G) can be covalently conjugated to proteins by isopeptide linkage to the ε-amino group of lysine (K) residues or less frequently to the N-terminal α-amino group or the thiol group of cysteine residues. Ubiquitin itself contains eight amino groups to which another ubiquitin molecule can be conjugated: the ε-amino groups of seven K residues (K6, 11, 27, 29, 33, 48 and 63) and the α-amino group of the N-terminal methionine residue. All of the eight ubiquitin chain types are present in the cell, among which the K48- and K63-linked chains are most abundant and the best described. K48-based linkages lead mainly to the proteasome-mediated degradation of the ubiquitinated protein, while K63-based chains control primarily protein endocytosis, trafficking, and enzyme activity [1,2,3,4].
Mechanistically, the process of protein ubiquitination involves a three-step enzymatic cascade, which starts with the ubiquitin-activating enzyme E1, followed by the ubiquitin-conjugating enzyme E2, and the ubiquitin ligase E3. Ubiquitin is activated in an ATP-dependent manner, when a high-energy thioester bond is formed between the C-terminus of ubiquitin and an internal cysteine residue of the ubiquitin-activating enzyme E1. Activated ubiquitin is then transferred onto the active site cysteine of one of E2-conjugating enzymes. Finally, the formation of an isopeptide bond is catalyzed by E3 ubiquitin ligases, which link ubiquitin moieties to target proteins or elongate a polyubiquitin chain (Figure 1) [5,6,7].
In complex with E2, the E3 ubiquitin ligase forms an isopeptide bond between ubiquitin moieties or between ubiquitin and substrate protein. In most cases the type of linkage is determined by E2 enzymes, except for the linkage between the amino group of the N-terminal methionine residue, determined by the E3 ubiquitin ligase called linear ubiquitin chain assembly complex (LUBAC) [8]. The E3 ubiquitin ligases determine the substrate specificity of ubiquitination, and the diversity of the cellular functions of ubiquitination is reflected in the existence of hundreds of different E3s in mammals, compared with roughly thirty-five E2s and only two E1s in humans. E3 enzymes are currently classified into three main families with different structural and functional characteristics: the homologous to E6AP C-terminus (HECT) domain family of ubiquitin ligases, the cullin-really interesting new gene (RING) family of ubiquitin ligases, and the U-box containing ubiquitin ligases [2,3,9]. E3 ligases can be single- or multi-subunit enzymes; in the second case ubiquitin-binding and substrate binding domains reside on separate polypeptides brought together by adaptor proteins.
Ubiquitinated substrates are subsequently recognized by a large number of proteins that contain different ubiquitin-binding domains; among these are DUBs, a group of about 100 enzymes in mammals that hydrolyze isopeptide linkages between ubiquitin moieties, or between ubiquitin and the substrate. DUBs do more than cancelling the ubiquitin mark: they differ in substrate and linkage type specificity, and in the position of the linkage in the polyubiquitin chain (between ubiquitin moieties or between ubiquitin and the substrate) [10,11]. Hydrolytic activity of DUBs leads to recycling of mono-ubiquitin in the cell, and to generation of free ubiquitin chains, which regulate aggresome formation and innate immune signaling [12,13].
2. Viruses Interact with Ubiquitination
Viruses have evolved a large arsenal of strategies to exploit processes regulated by ubiquitin (for a detailed review, refer to [14,15]). They may target unwanted cellular proteins for degradation by K48-linked polyubiquitination or revert undesirable ubiquitination events through deubiquitination. Some viruses encode their own E3 ubiquitin ligases and DUBs, such as infected cell polypeptide 0 (ICP0) of herpes simplex virus type 1 (HSV-1), a multifunctional protein with RING domain E3 ubiquitin ligase activity [16]. Others subvert the specificity of cellular E3 ubiquitin ligases and DUBs in order to avoid self-degradation.
The importance of ubiquitination for virus infection has also been pointed out in multiple studies based on the treatment of infected cells with proteasome inhibitors. Such a treatment not only blocks the ubiquitin proteasome system (UPS), but also depletes the cellular pool of free ubiquitin, affecting many of the cellular pathways involving ubiquitin. Functional UPS is important for the replication of major human pathogens such as herpesviruses, poxviruses, hepadnaviruses, adenoviruses, influenzaviruses, retroviruses, coronaviruses, paramyxoviruses, picornaviruses and rotaviruses [14]. UPS components are upregulated in primary human airway epithelial cells infected by influenza A virus (IAV) [17]. Proteasome inhibition leads to sequestration of incoming IAV in the cytoplasmic compartments and blocks productive viral entry [18,19].
Ubiquitination also plays an important role in innate immunity to IAV. The host antiviral factor cyclophilin A targets the influenza matrix protein (M1) for ubiquitin-mediated degradation [20], while zinc finger antiviral protein ZAPL promotes ubiquitination and degradation of viral polymerase subunits PA and PB2 [21]. Interferon (IFN)-induced transmembrane protein 3 (IFITM3), a potent antiviral protein which inhibits IAV cytosolic entry [22,23], is downregulated by ubiquitination by E3 ubiquitin ligase neural precursor cell expressed developmentally downregulated protein 4 (NEDD4), the depletion of which boosts cell resistance to IAV infection [24]. Tripartite motif-containing (TRIM) proteins, which belong to the RING family of E3 ubiquitin ligases, counteract IAV replication. For example, TRIM25 is a key mediator in antiviral retinoic acid-inducible gene 1 (RIG-I) signaling [25] (described in Section 7). TRIM22 and TRIM32 ubiquitinate nucleoprotein (NP) and viral polymerase subunit PB1, respectively [26,27], targeting them for degradation. IAV proteins indirectly modulate ubiquitination events to evade immune responses and support virus replication. For example, nonstructural protein 1 (NS1) inhibits TRIM25 [28]. Expression of viral hemagglutinin (HA) in cells causes phosphorylation and ubiquitination of IFN receptor subunit 1 (IFNAR1) which then undergoes proteasome- and lysosome-dependent degradation [29]. Reduction of IFNAR1 on cell surface attenuates IFN signaling and innate antiviral responses.
3. Ubiquitin Regulates Influenza Virus Entry and Replication
The entry of viruses into animal cells follows a sequence of events including attachment of the virus particle to the cell surface, signaling, internalization, endocytic traffic, fusion, capsid uncoating, and genome release [30,31]. IAV binds to sialic acids on cell surface glycoproteins and glycolipids [32], which leads to lipid-raft clustering and activation of epidermal growth factor receptor (EGFR) and receptor tyrosine kinases that facilitate clathrin-mediated endocytosis and macropinocytosis [33,34,35].
The viral M2 channel opens in mildly acidic endosomes, allowing protons and potassium ions to enter the viral core [36,37]. This primes the core for uncoating by weakening core protein interactions. The importance of priming is signified by the low infection rate in acid-mediated bypass experiments (in which the virus enters the cytoplasm by direct fusion at the plasma membrane) compared to infection by the endocytic route [37]. In late endosomes (LEs) or mature macropinosomes, viral envelope glycoprotein HA undergoes an irreversible, low pH (5.5–5.0)-induced conformational change that activates its membrane fusion activity [38]. The viral envelope fuses with the limiting membrane of the endosome to form a fusion pore, through which the particle gains access to the cytosolic milieu for the first time during entry.
Ubiquitination plays a key role in IAV entry, especially during the step of uncoating. The itchy E3 ubiquitin protein ligase (ITCH) has been shown to promote IAV escape from LEs via the ubiquitination of M1 [39]. Uncoating is inhibited by the depletion of cullin 3 (Cul3), an E3 ubiquitin ligase that promotes endosome maturation [40,41]; the defects in the LE compartments in Cul3-depleted cells were reflected by a highly vacuolated LE/lysosome (LY) phenotype with large spherical, Rab7-positive, fluid-filled endosomes [40]. The endosomal sorting complexes required for transport (ESCRTs) mediate the sorting of ubiquitinated membrane proteins into intraluminal vesicles (ILVs) [42,43]. Depletion of components of the ESCRT machinery results in fewer ILVs and accumulation of cargo in endosomes with abnormal morphology [44,45,46].
Cul3 and the broad complex (BrC), tramtrack (Ttk) and bric-à-brac (Bab) (BTB)-adaptor speckle type BTB/pox virus and zinc finger (POZ) protein like (SPOPL) is responsible for the ubiquitination and degradation of the endosomal adaptor protein epidermal growth factor receptor pathway substrate 15 (EPS15) [47]. In the absence of SPOPL, EPS15 degradation and subsequent ILV formation are compromised, inhibiting EGFR degradation and IAV uncoating [40,47]. After uncoating via histone deacetylase 6 (HDAC6) and aggresome processing (discussed in Section 6), viral ribonucleoproteins (vRNPs) are released into the cytosol and trafficked to the nuclear pore complex (NPC) independent of microtubules or actin filaments [48,49,50]. They enter the nucleus via the activity of importin α/β where, in a complex with various co-opted host factors, the viral polymerase directs transcription and replication of the genome [51,52,53].
Post nuclear entry, the monoubiquitination of NP at residue K184 is crucial for virus RNA replication, possibly by regulation of NP–RNA interaction. Ubiquitination at K184 is counteracted by a host deubiquitinase USP11 [54]. Ubiquitination of viral polymerase and UPS activity promotes the activity of the polymerase, and ubiquitin expression leads to accumulation of vRNA, complementary RNA (cRNA) and viral mRNA [55]. Nonstructural protein 2 (NS2) of IAV binds to aminoacyl-tRNA synthase complex-interacting multifunctional protein 2 (AIMP2) which protects AIMP2 from K48-linked polyubiquitination and degradation. AIMP2 promotes nuclear export of vRNPs and IAV replication at late steps of infection. It also counteracts ubiquitination of M1 at K242 and subsequent proteasomal degradation by an unknown mechanism [56]. Modification of the same M1 K242 by a small ubiquitin-like modifier (SUMO) is crucial for nuclear export of newly produced vRNPs and viral morphogenesis [57], which could explain AIMP2-mediated enhancement of IAV replication at late steps of infection [56].
4. HDAC6 Binds to Unanchored Ubiquitin
HDAC6 is a cytoplasmic enzyme that promotes autophagic clearance of protein aggregates and protects cells from cytotoxic accumulation of misfolded aggregated proteins [58,59,60,61,62,63,64]. The two catalytic domains CD1 and CD2 of HDAC6 form together an ellipsoid-shaped complex of pseudo-two-fold symmetry [65]. Between the two catalytic domains there is a polypeptide that binds to the dynein motor [66,67]. Besides enzymatic activity, HDAC6 has an intrinsic ubiquitin-binding capability carried out by a zinc finger ubiquitin-binding domain (ZnF-UBP) close to the C-terminus (Figure 2), making it unique among HDAC family proteins [68].
Ubiquitin moieties in polyubiquitinated substrates are typically recognized by UBPs through their globular surface and hydrophobic core [69]. HDAC6 ZnF-UBP, on the other hand, has a unique deep binding pocket where ubiquitin’s C-terminal diglycine motif enters and binds with high affinity and specificity [70]. Thus, HDAC6 ZnF-UBP only binds to free mono ubiquitin and unanchored ubiquitin chains (Figure 2). Biochemical and structural analysis of the ZnF-UBP in complex with the full-length ubiquitin or the ubiquitin C-terminal RLRGG-peptide confirmed the necessity of the terminal diglycine for ZnF-UBP binding [63,71]. When ubiquitin is anchored to a target polypeptide via the C-terminus, HDAC6 ZnF-UBP cannot bind to it.
5. HDAC6 Regulates Aggresome Processing
Misfolded proteins are tagged by polyubiquitin and targeted for degradation by proteasome activity. However, if clearance of misfolded proteins is insufficient, they can shape sizable aggregates that interfere with cell homeostasis [74]. How does HDAC6 ZnF-UBP recruit protein aggregates? Ouyang et al. [63] showed that unanchored ubiquitin chains with various lengths are found in misfolded protein bundles. Unanchored ubiquitin chains are generated in situ by Ataxin-3, an aggregate-associated, polyubiquitin-editing DUB that cuts within ubiquitin chains. Ataxin-3 exposes C-terminal diglycine motifs that serve as recognition tags for protein waste. These tags recruit HDAC6 and facilitate subsequent dynein-mediated transport of the aggregate to the microtubule-organizing center (MTOC) (Figure 3). How do the ubiquitin chains, despite being unanchored, remain associated with the aggregates? One possibility is via interaction of the ubiquitin hydrophobic patch with exposed hydrophobic cores of the misfolded proteins. In this scenario, ubiquitin chains would become trapped inside a net of aggregated polypeptides and cannot diffuse away.
Ataxin-3 preferentially cleaves K63-polyubiquitinated substrates [75], and when Ataxin-3 is depleted, HDAC6 is incapable of associating with protein aggregates [63]. Proteasomes do not degrade aggresomes but tend to congregate in their vicinity [12,76]. A proteasome-associated K63-specific DUB called Poh1 is necessary for aggresome clearance because it produces unanchored K63-linked ubiquitin chains which activate HDAC6 [12]. Heat shock protein 90 (Hsp90) is a binding partner of HDAC6 [77] and contributes to aggresome clearance by stimulating Poh1 to produce unanchored ubiquitin chains [76]. Ubiquitin binding to HDAC6 regulates the repressive Hsp90–heat shock factor 1 (HSF1) complex, the dissociation of which leads to activation of HSF1 and expression of cellular chaperones [58]. In Poh1-deficient cells, aggresome clearance is inhibited but can be restored by microinjecting unanchored K63-linked ubiquitin [12]. Toxic protein aggregates are processed by autophagy after being sequestered to the MTOC, and HDAC6 regulates the formation of a cortactin-dependent F-actin network that is interspersed among such protein aggregates. This activity stimulates autophagosome-LY fusion and aggresome degradation [61] (Figure 3).
6. Influenza Virus Uses Aggresome Processing for Capsid Uncoating
IAV X31 virions encapsidate a variety of unanchored ubiquitin moieties which include mono-, di-, tri-, tetra-, penta-, hexa-, and hepta-ubiquitin chains [66]. Following viral fusion at LEs, these ubiquitin moieties are exposed to the cytoplasm and attract HDAC6 to the sites of fusion on the surface of LEs. HDAC6 in turn binds to the capsid, which during endocytic transit has been broken down into M1 dimers [81], and links it to cytoskeleton motors dynein and myosin. This generates a shearing force that disassembles the capsid and releases the vRNPs into the cytoplasm (Figure 3). Point mutations H1094/H1098 or W1116 of the mouse HDAC6 ZnF-UBP inactivate ubiquitin binding to HDAC6 and block uncoating [66,82]. By blocking HDAC6 binding to dynein, dynein and myosin activity, or by depolymerizing microtubules or actin, uncoating is diminished [66]. It is likely that DUBs assist in the process of IAV uncoating. This is because DUBs such as ataxin-3 [63] and Poh1 [12] that generate unanchored ubiquitin chains are required for HDAC6-mediated aggresome processing.
What promotes virus uncoating during cell entry can potentially interfere with the assembly of newly replicated virions. To ensure proper assembly, viruses remove uncoating factors during the late stage of infection [30]. The IAV strategy is dually cunning; it not only inactivates HDAC6 but also induces a stable microtubule network that promotes viral progeny egress and budding. During replication, IAV cleaves off the HDAC6 ZnF-UBP by activating caspase 3 [83], which prevents premature capsid uncoating. HDAC6 inactivation combined with virus replication increases acetylation of microtubules. Acetylation promotes the association of microtubules with kinesin-1 and dynein motors which regulate endosome trafficking [84,85,86,87]. Cells lacking HDAC6 deliver EGFR prematurely to LE/LY compartments, resulting in faster EGFR degradation [88]. Hyperacetylated microtubules promote IAV genome egress via Rab11-positive recycling endosomes [89,90,91,92] (Figure 4). Although hyperacetylated microtubules appear to play no role in IAV uncoating [66], we cannot exclude the possibility that kinesin, in addition to dynein and myosin, contributes to the uncoating process.
Some viruses including poxvirus, African swine fever virus (ASFV), herpesviruses and retroviruses induce the formation of inclusion bodies near the MTOC. These so-called virus factories or viroplasms resemble aggresomes, and their contribution to virus replication and innate immune response has been discussed in detail by Wileman [93]. Interestingly, HDAC6 activates IFN-β [94] and regulates the RIG-I-mediated antiviral innate immune response. Deacetylation of RIG-I K909 by HDAC6 promotes viral RNA-sensing activity [80], and RNA viruses replicate better in immune cells depleted of HDAC6 due to reduction in IFN-β and proinflammatory cytokines [80]. However, the in vivo effects of IAV challenge in these mice remain to be determined.
7. Unanchored Ubiquitin Regulates RIG-I-Like Receptor Signaling
The role of innate immunity in RNA virus infection has been reviewed on many occasions [95,96,97], and we will only discuss some of the general aspects linked to ubiquitination. Pathogen-associated molecular patterns (PAMPs) of invading viruses are recognized by pattern recognition receptors (PRR). This leads to the activation of different signaling cascades, with the final production of IFNs the mediators of antiviral responses. Regulation of innate immune signaling relies on post-translational modifications including ubiquitination, and many viruses have evolved mechanisms to alter these ubiquitination events [14].
RIG-I, a member of the RIG-I-like receptor (RLR) family, is the most important PRR for viral detection and type I IFN production in infected epithelial cells. The two best-studied RLR members—RIG-I and melanoma differentiation-associated protein 5 (MDA5)—are cytoplasmic sensors critical for the detection of viral 5′-triphosphate single-stranded RNA (ssRNA) and long double-stranded RNA (dsRNA), respectively, which are generated after viral replication. Upon recognition of viral RNA, the helicase domain of RIG-I undergoes conformational changes that enable its caspase-recruitment domains (CARDs) to bind to mitochondrial antiviral signaling protein (MAVS). This finally leads to activation of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and interferon response factor (IRF), and to the expression of type I IFNs [95,96,97]. K63-linked polyubiquitination of CARD by the E3 ubiquitin ligase TRIM25 induces conformational change and CARD tetramerization, which is necessary for signal transduction to MAVS [25].
Another E3 ubiquitin ligase, Riplet, was shown to regulate RIG-I through K63-linked ubiquitination [98,99]. Recent reports suggest that not only covalently attached but also unanchored K63-linked polyubiquitin chains activate RIG-I signaling. However, according to structural studies, unanchored ubiquitin chains stabilize the signaling-active RIG-I tetramer less efficiently than covalent K63-ubiquitin [100]. IRF3 is activated by in vitro-generated K63-polyubiquitin chains that bind to RIG-I [13], but the role of unanchored K63-linked ubiquitin in MDA5-mediated IRF3 activation is not clear cut [101,102].
8. Regulation of RIG-I by Ubiquitin Is Species-Specific
Influenza pandemics can occur when zoonotic influenza viruses adapt for efficient replication and transmission in humans. The incompatibility of avian IAV strains with the human host can be explained by the lack of factors supporting viral replication in human cells or the presence of restriction factors that cannot be counteracted by avian strains until adaptive mutations are selected [103]. It has been known for some time that in human cells, avian-derived IAV polymerase is poorly active. In heterokaryons formed between human and avian cells, the activity of avian IAV polymerase is restored [104], and the responsible host factor co-opted in avian cells was recently identified as ANP32A [105].
While in humans activation of RIG-I by unanchored ubiquitin seems to be only an accessory mechanism to covalent ubiquitination, in ducks, the natural host of IAV, unanchored ubiquitin may play the main role in RIG-I signaling. It has been shown that duck TRIM25 can activate RIG-I CARD mutant lacking lysine residues by a mechanism independent of ubiquitination [106]. TRIM25 produces unanchored ubiquitin chains that activate RIG-I in vitro [13], suggesting that interaction with unanchored ubiquitin might regulate duck RIG-I. Adaptation of IAV strains to counteract different immune responses in avian and human hosts likely contributes to interspecies transmission and pandemic risk.
9. Ubiquitin Regulates Inflammasomes
RIG-I activates inflammasomes, the key components of the innate immune response to pathogens or tissue damage [107]. These large multiprotein complexes are formed rapidly in response to PAMPs and serve as scaffolds to activate caspase 1, which processes pro-interleukin (IL)-1β and pro-IL-18 into IL-1β and IL-18, respectively, which can then be secreted [95,107]. Nucleotide-binding oligomerization domain (NOD)-like receptor family member NACHT, LRR and PYD domains-containing protein 3 (NLRP3) inflammasome is involved in defense against infection with IAV and many other viruses including hepatitis C virus [107,108,109,110,111]. Ubiquitination and deubiquitination regulates NLRP3 inflammasome activation. For example, breast cancer 1 (BRCA1)/breast cancer 2 (BRCA2)-containing complex subunit 3 (BRCC3) deubiquitinates NLRP3, and pharmaceutical inhibition of DUBs restricts inflammasome activation [79,112,113], whereas E3 ubiquitin ligases LUBAC and TRIM33 promote inflammasome assembly [114,115]. In bone-marrow-derived macrophages (BMDMs) depleted of HDAC6, there is a significant boost in caspase 1 activation and IL-1β secretion as a response to NLRP3 activation. HDAC6 ZnF-UBP interaction with NLRP3 inhibits inflammasome activation [79], suggesting that unanchored ubiquitin regulates this pathway. NS1, the major suppressor of the IAV IFN response, antagonizes both TRIM25/Riplet-mediated RIG-I ubiquitination and NLRP3 inflammasome activation which underscores their significance in anti-viral immunity [28,108,116].
10. Future Perspectives
Incorporation of host cellular proteins into or onto the newly formed enveloped viruses is a known phenomenon. A few of such virus-incorporated host proteins have been described, especially in retroviruses [117]. IAV virions produced in different hosts contain a wide range of host-derived proteins as detected by mass spectrometry [118,119]. Among these are ubiquitin, cytoskeletal proteins, and tetraspanin cluster of differentiation (CD) 81 [118,119,120]. The biological significance of these non-viral proteins is largely unknown; encapsidated enzymes, for example, could play a regulatory role in IAV entry. In order to better understand how HDAC6 contributes to IAV uncoating, the following aspects should be addressed: (1) virion ubiquitin content (e.g., abundance of unanchored moieties and their linkages); (2) architecture of the viral fusion pore; and (3) availability of other uncoating factors in the cytoplasm.
Can viral ubiquitin be used as a predictor of IAV infectivity? Does unanchored ubiquitin regulate the immune response during IAV entry? It is intriguing that K63-linked ubiquitin chains are implicated in aggresome clearance, as this modification is an element of immune signaling [121] rather than the UPS (often K48-linked). There is perhaps an unexplored link between ubiquitin, aggresome processing, and immune response to viral infection. Unraveling such molecular interactions between virus and host cell is of critical importance for understanding the cell biology of virus infection, and for future design of antivirals.
Acknowledgments
This work was supported by VirX, a Swiss National Science Foundation (SNSF) SystemsX.ch Medical Research and Development (MRD) grant awarded to Y.Y. We thank Samuel Nobs, Dan Rocca, and Indranil Banerjee for comments on the manuscript.
Author Contributions
A.R. and Y.Y. conceived and wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chen, Z.J.; Sun, L.J. Nonproteolytic Functions of Ubiquitin in Cell Signaling. Mol. Cell. 2009, 33, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Komander, D.; Rape, M. The Ubiquitin Code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef] [PubMed]
- Kulathu, Y.; Komander, D. Atypical ubiquitylation — the unexplored world of polyubiquitin beyond Lys48 and Lys63 linkages. Nat. Rev. Mol. Cell Biol. 2012, 13, 508–523. [Google Scholar] [CrossRef] [PubMed]
- Tanno, H.; Komada, M. The ubiquitin code and its decoding machinery in the endocytic pathway. J. Biochem. 2013, 153, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Chau, V.; Tobias, J.W.; Bachmair, A.; Marriott, D.; Ecker, D.J.; Gonda, D.K.; Varshavsky, A. A multiubiquitin chain is confined to specific lysine in a targeted short-lived protein. Science 1989, 243, 1576–1583. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A. The unravelling of the ubiquitin system. Nat. Rev. Mol. Cell Biol. 2015, 16, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Hershko, A.; Heller, H.; Elias, S.; Ciechanover, A. Components of ubiquitin-protein ligase system. Resolution, affinity purification, and role in protein breakdown. J. Biol. Chem. 1983, 258, 8206–8214. [Google Scholar] [PubMed]
- Kirisako, T.; Kamei, K.; Murata, S.; Kato, M.; Fukumoto, H.; Kanie, M.; Sano, S.; Tokunaga, F.; Tanaka, K.; Iwai, K. A ubiquitin ligase complex assembles linear polyubiquitin chains. EMBO J. 2006, 25, 4877–4887. [Google Scholar] [CrossRef] [PubMed]
- Metzger, M.B.; Hristova, V.A.; Weissman, A.M. HECT and RING finger families of E3 ubiquitin ligases at a glance. J. Cell Sci. 2012, 125, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Clague, M.J.; Barsukov, I.; Coulson, J.M.; Liu, H.; Rigden, D.J.; Urbé, S. Deubiquitylases from genes to organism. Physiol. Rev. 2013, 93, 1289–1315. [Google Scholar] [CrossRef] [PubMed]
- Komander, D.; Clague, M.J.; Urbé, S. Breaking the chains: structure and function of the deubiquitinases. Nat. Rev. Mol. Cell Biol. 2009, 10, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Hao, R.; Nanduri, P.; Rao, Y.; Panichelli, R.S.; Ito, A.; Yoshida, M.; Yao, T.P. Proteasomes activate aggresome disassembly and clearance by producing unanchored ubiquitin chains. Mol. Cell. 2013, 51, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Sun, L.; Jiang, X.; Chen, X.; Hou, F.; Adhikari, A.; Xu, M.; Chen, Z.J. Reconstitution of the RIG-I pathway reveals a signaling role of unanchored polyubiquitin chains in innate immunity. Cell. 2010, 141, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Calistri, A.; Munegato, D.; Carli, I.; Parolin, C.; Palù, G. The Ubiquitin-Conjugating System: Multiple Roles in Viral Replication and Infection. Cells 2014, 3, 386–417. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, P.; Schreiner, S. Viral Mimicry to Usurp Ubiquitin and SUMO Host Pathways. Viruses 2015, 7, 4854–4872. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.C.; Boutell, C.; Davido, D.J. HSV-1 ICP0: paving the way for viral replication. Future Virol. 2011, 6, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Kroeker, A.L.; Ezzati, P.; Coombs, K.M.; Halayko, A.J. Influenza A infection of primary human airway epithelial cells up-regulates proteins related to purine metabolism and ubiquitin-related signaling. J. Proteome Res. 2013, 12, 3139–3151. [Google Scholar] [CrossRef] [PubMed]
- Khor, R.; McElroy, L.J.; Whittaker, G.R. The Ubiquitin-Vacuolar Protein Sorting System is Selectively Required During Entry of Influenza Virus into Host Cells. Traffic 2003, 4, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Widjaja, I.; de Vries, E.; Tscherne, D.M.; García-Sastre, A.; Rottier, P.J.M.; de Haan, C.A.M. Inhibition of the ubiquitin-proteasome system affects influenza A virus infection at a postfusion step. J. Virol. 2010, 84, 9625–9631. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, Z.; Xu, C.; Sun, L.; Chen, J.; Zhang, L.; Liu, W. Cyclophilin A Restricts Influenza A Virus Replication through Degradation of the M1 Protein. PLoS ONE 2012, 7, e31063. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-H.; Zhou, L.; Chen, G.; Krug, R.M. Battle between influenza A virus and a newly identified antiviral activity of the PARP-containing ZAPL protein. Proc. Natl. Acad. Sci. USA 2015, 112, 14048–14053. [Google Scholar] [CrossRef] [PubMed]
- Feeley, E.M.; Sims, J.S.; John, S.P.; Chin, C.R.; Pertel, T.; Chen, L.M.; Gaiha, G.D.; Ryan, B.J.; Donis, R.O.; Elledge, S.J.; et al. IFITM3 inhibits influenza a virus infection by preventing cytosolic entry. PLoS Pathog. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Desai, T.M.; Marin, M.; Chin, C.R.; Savidis, G.; Brass, A.L.; Melikyan, G.B. IFITM3 Restricts Influenza A Virus Entry by Blocking the Formation of Fusion Pores following Virus-Endosome Hemifusion. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Chesarino, N.M.; McMichael, T.M.; Yount, J.S. E3 Ubiquitin Ligase NEDD4 Promotes Influenza Virus Infection by Decreasing Levels of the Antiviral Protein IFITM3. PLoS Pathog. 2015, 11, e1005095. [Google Scholar] [CrossRef] [PubMed]
- Gack, M.U.; Shin, Y.C.; Joo, C.-H.; Urano, T.; Liang, C.; Sun, L.; Takeuchi, O.; Akira, S.; Chen, Z.; Inoue, S.; et al. TRIM25 RING-finger E3 ubiquitin ligase is essential for RIG-I-mediated antiviral activity. Nature 2007, 446, 916–920. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, A.; Kajaste-Rudnitski, A.; Oteiza, A.; Nicora, L.; Towers, G.J.; Mechti, N.; Vicenzi, E. TRIM22 inhibits influenza A virus infection by targeting the viral nucleoprotein for degradation. J. Virol. 2013, 87, 4523–4533. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Wang, L.; Ding, H.; Schwamborn, J.C.; Li, S.; Dorf, M.E. TRIM32 Senses and Restricts Influenza A Virus by Ubiquitination of PB1 Polymerase. PLoS Pathog. 2015, 11, e1004960. [Google Scholar] [CrossRef] [PubMed]
- Gack, M.U.; Albrecht, R.A.; Urano, T.; Inn, K.-S.; Huang, I.-C.; Carnero, E.; Farzan, M.; Inoue, S.; Jung, J.U.; García-Sastre, A. Influenza A virus NS1 targets the ubiquitin ligase TRIM25 to evade recognition by the host viral RNA sensor RIG-I. Cell Host Microbe 2009, 5, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Vijayan, M.; Pritzl, C.J.; Fuchs, S.Y.; McDermott, A.B.; Hahm, B. Hemagglutinin of Influenza A Virus Antagonizes Type I Interferon (IFN) Responses by Inducing Degradation of Type I IFN Receptor 1. J. Virol. 2016, 90, 2403–2417. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Greber, U.F. Principles of Virus Uncoating: Cues and the Snooker Ball. Traffic 2016, 17, 569–592. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Helenius, A. Virus entry at a glance. J. Cell Sci. 2013, 126, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Ito, T.; Suzuki, T.; Holland, R.E.; Chambers, T.M.; Kiso, M.; Ishida, H.; Kawaoka, Y. Sialic Acid Species as a Determinant of the Host Range of Influenza A Viruses. J. Virol. 2000, 74, 11825–11831. [Google Scholar] [CrossRef] [PubMed]
- Eierhoff, T.; Hrincius, E.R.; Rescher, U.; Ludwig, S.; Ehrhardt, C. The epidermal growth factor receptor (EGFR) promotes uptake of influenza a viruses (IAV) into host cells. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef] [PubMed]
- Matlin, K.S.; Reggio, H.; Helenius, A.; Simons, K. Infectious entry pathway of influenza virus in a canine kidney cell line. J. Cell Biol. 1981, 91, 601–613. [Google Scholar] [CrossRef] [PubMed]
- De Vries, E.; Tscherne, D.M.; Wienholts, M.J.; Cobos-Jiménez, V.; Scholte, F.; García-Sastre, A.; Rottier, P.J.; de Haan, C.A. Dissection of the influenza A virus endocytic routes reveals macropinocytosis as an alternative entry pathway. PLoS Pathog. 2011, 7, e1001329. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.H.; Holsinger, L.J.; Lamb, R.A. Influenza virus M2 protein has ion channel activity. Cell 1992, 69, 517–528. [Google Scholar] [CrossRef]
- Stauffer, S.; Feng, Y.; Nebioglu, F.; Heilig, R.; Picotti, P.; Helenius, A. Stepwise Priming by Acidic pH and a High K+ Concentration Is Required for Efficient Uncoating of Influenza A Virus Cores after Penetration. J. Virol. 2014, 88, 13029–13046. [Google Scholar] [CrossRef] [PubMed]
- White, J.; Kartenbeck, J.; Helenius, A. Membrane fusion activity of influenza virus. EMBO J. 1982, 1, 217–222. [Google Scholar] [PubMed]
- Su, W.-C.; Chen, Y.-C.; Tseng, C.-H.; Hsu, P.W.-C.; Tung, K.-F.; Jeng, K.-S.; Lai, M.M. Pooled RNAi screen identifies ubiquitin ligase Itch as crucial for influenza A virus release from the endosome during virus entry. Proc. Natl. Acad. Sci. USA 2013, 110, 17516–17521. [Google Scholar] [CrossRef] [PubMed]
- Huotari, J.; Meyer-Schaller, N.; Hubner, M.; Stauffer, S.; Katheder, N.; Horvath, P.; Mancini, R.; Helenius, A.; Peter, M. Cullin-3 regulates late endosome maturation. Proc. Natl. Acad. Sci. USA 2012, 109, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Hubner, M.; Peter, M. Cullin-3 and the endocytic system: New functions of ubiquitination for endosome maturation. Cell. Logist. 2012, 2, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Raiborg, C.; Bache, K.G.; Gillooly, D.J.; Madshus, I.H.; Stang, E.; Stenmark, H. Hrs sorts ubiquitinated proteins into clathrin-coated microdomains of early endosomes. Nat. Cell Biol. 2002, 4, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Sachse, M.; Urbé, S.; Oorschot, V.; Strous, G.J.; Klumperman, J. Bilayered clathrin coats on endosomal vacuoles are involved in protein sorting toward lysosomes. Mol. Biol. Cell 2002, 13, 1313–1328. [Google Scholar] [CrossRef] [PubMed]
- Doyotte, A.; Russell, M.R.G.; Hopkins, C.R.; Woodman, P.G. Depletion of TSG101 forms a mammalian “Class E” compartment: a multicisternal early endosome with multiple sorting defects. J. Cell Sci. 2005, 118, 3003–3017. [Google Scholar] [CrossRef] [PubMed]
- Razi, M.; Futter, C.E. Distinct roles for Tsg101 and Hrs in multivesicular body formation and inward vesiculation. Mol. Biol. Cell 2006, 17, 3469–3483. [Google Scholar] [CrossRef] [PubMed]
- Rieder, S.E.; Banta, L.M.; Köhrer, K.; McCaffery, J.M.; Emr, S.D. Multilamellar endosome-like compartment accumulates in the yeast vps28 vacuolar protein sorting mutant. Mol. Biol. Cell 1996, 7, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Gschweitl, M.; Ulbricht, A.; Barnes, C.A.; Enchev, R.I.; Stoffel-Studer, I.; Meyer-Schaller, N.; Huotari, J.; Yamauchi, Y.; Greber, U.F.; Helenius, A.; et al. A SPOPL/Cullin-3 ubiquitin ligase complex regulates endocytic trafficking by targeting EPS15 at endosomes. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, N.L.; Shen, Y.; Bao, X.; Fabrizio, T.; Elbahesh, H.; Webby, R.J.; Li, K. The C-terminal Tail of TRIM56 Dictates Antiviral Restriction of Influenza A and B Viruses by Impeding Viral RNA Synthesis. J. Virol. 2016, 90, JVI.03172-15. [Google Scholar] [CrossRef] [PubMed]
- Babcock, H.P.; Chen, C.; Zhuang, X. Using single-particle tracking to study nuclear trafficking of viral genes. Biophys. J. 2004, 87, 2749–2758. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.; Helenius, A. Transport of incoming influenza virus nucleocapsids into the nucleus. J. Virol. 1991, 65, 232–244. [Google Scholar] [CrossRef]
- Cros, J.F.; García-Sastre, A.; Palese, P. An Unconventional NLS is Critical for the Nuclear Import of the Influenza A Virus Nucleoprotein and Ribonucleoprotein. Traffic 2005, 6, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Eisfeld, A.J.; Neumann, G.; Kawaoka, Y. At the centre: influenza A virus ribonucleoproteins. Nat. Rev. Microbiol. 2014, 13, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.W.H.; Sun, Y.-H.B.; Panté, N. Nuclear import of influenza A viral ribonucleoprotein complexes is mediated by two nuclear localization sequences on viral nucleoprotein. Virol. J. 2007, 4, 49. [Google Scholar] [CrossRef] [PubMed]
- Liao, T.-L.; Wu, C.-Y.; Su, W.-C.; Jeng, K.-S.; Lai, M.M.C. Ubiquitination and deubiquitination of NP protein regulates influenza A virus RNA replication. EMBO J. 2010, 29, 3879–3890. [Google Scholar] [CrossRef] [PubMed]
- Kirui, J.; Mondal, A.; Mehle, A. Ubiquitination up-regulates influenza virus polymerase function. J. Virol. 2016. JVI.01829-16. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Wu, J.; Liu, R.-Y.; Li, J.; Song, L.; Teng, Y.; Sheng, C.; Liu, D.; Yao, C.; Chen, H.; et al. Interaction of NS2 with AIMP2 facilitates the switch from ubiquitination to SUMOylation of M1 in influenza A virus-infected cells. J. Virol. 2015, 89, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-Y.; Jeng, K.-S.; Lai, M.M. The SUMOylation of Matrix Protein M1 Modulates the Assembly and Morphogenesis of Influenza A Virus. J. Virol. 2011, 85, 6618–6628. [Google Scholar] [CrossRef] [PubMed]
- Boyault, C.; Zhang, Y.; Fritah, S.; Caron, C.; Gilquin, B.; Kwon, S.H.; Garrido, C.; Yao, T.P.; Vourc’h, C.; Matthias, P.; et al. HDAC6 controls major cell response pathways to cytotoxic accumulation of protein aggregates. Genes Dev. 2007, 21, 2172–2181. [Google Scholar] [CrossRef] [PubMed]
- Hubbert, C.; Guardiola, A.; Shao, R.; Kawaguchi, Y.; Ito, A.; Nixon, A.; Yoshida, M.; Wang, X.F.; Yao, T.P. HDAC6 is a microtubule-associated deacetylase. Nature 2002, 417, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Iwata, A.; Riley, B.E.; Johnston, J.A.; Kopito, R.R. HDAC6 and microtubules are required for autophagic degradation of aggregated Huntingtin. J. Biol. Chem. 2005, 280, 40282–40292. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Koga, H.; Kawaguchi, Y.; Tang, W.; Wong, E.; Gao, Y.-S.; Pandey, U.B.; Kaushik, S.; Tresse, E.; Lu, J.; et al. HDAC6 controls autophagosome maturation essential for ubiquitin-selective quality-control autophagy. EMBO J. 2010, 29, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Matthias, P.; Yoshida, M.; Khochbin, S. HDAC6 a new cellular stress surveillance factor. Cell Cycle 2008, 7, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, H.; Ali, Y.O.; Ravichandran, M.; Dong, A.; Qiu, W.; MacKenzie, F.; Dhe-Paganon, S.; Arrowsmith, C.H.; Zhai, R.G. Protein aggregates are recruited to aggresome by histone deacetylase 6 via unanchored ubiquitin C termini. J. Biol. Chem. 2012, 287, 2317–2327. [Google Scholar] [CrossRef] [PubMed]
- Pandey, U.B.; Nie, Z.; Batlevi, Y.; McCray, B.A.; Ritson, G.P.; Nedelsky, N.B.; Schwartz, S.L.; DiProspero, N.A.; Knight, M.A.; Schuldiner, O.; et al. HDAC6 rescues neurodegeneration and provides an essential link between autophagy and the UPS. Nature 2007, 447, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Keusch, J.J.; Wang, L.; Saito, M.; Hess, D.; Wang, X.; Melancon, B.J.; Helquist, P.; Gut, H.; Matthias, P. Structural insights into HDAC6 tubulin deacetylation and its selective inhibition. Nat. Chem. Biol. 2016, 12, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, I.; Miyake, Y.; Nobs, S.P.; Schneider, C.; Horvath, P.; Kopf, M.; Matthias, P.; Helenius, A.; Yamauchi, Y. Influenza A virus uses the aggresome processing machinery for host cell entry. Science 2014, 346, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, Y.; Kovacs, J.J.; McLaurin, A.; Vance, J.M.; Ito, A.; Yao, T.-P. The Deacetylase HDAC6 Regulates Aggresome Formation and Cell Viability in Response to Misfolded Protein Stress. Cell 2003, 115, 727–738. [Google Scholar] [CrossRef]
- Bonnet, J.; Romier, C.; Tora, L.; Devys, D. Zinc-finger UBPs: regulators of deubiquitylation. Trends Biochem. Sci. 2008, 33, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Dikic, I.; Wakatsuki, S.; Walters, K.J. Ubiquitin-binding domains - from structures to functions. Nat. Rev. Mol. Cell Biol. 2009, 10, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Turcu, F.E.; Horton, J.R.; Mullally, J.E.; Heroux, A.; Cheng, X.; Wilkinson, K.D. The Ubiquitin Binding Domain ZnF UBP Recognizes the C-Terminal Diglycine Motif of Unanchored Ubiquitin. Cell 2006, 124, 1197–1208. [Google Scholar] [CrossRef] [PubMed]
- Dong, A.; Ravichandran, M.; Loppnau, P.; Li, Y.; MacKenzie, F.; Kozieradzki, I. Crystal structure of human HDAC6 zinc finger domain and ubiquitin C-terminal peptide RLRGG. Unpublished. [CrossRef]
- Hai, Y.; Christianson, D.W. Histone deacetylase 6 structure and molecular basis of catalysis and inhibition. Nat. Chem. Biol. 2016, 12, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Vijay-Kumar, S.; Bugg, C.; Cook, W.J. Structure of ubiquitin refined at 1.8 A resolution. J. Mol. Biol. 1987, 194, 531–544. [Google Scholar] [CrossRef]
- Johnston, J.A.; Ward, C.L.; Kopito, R.R. Aggresomes: A Cellular Response to Misfolded Proteins. J. Cell Biol. 1998, 143, 1883–1898. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ying, Z.; Wang, G. Ataxin-3 Regulates Aggresome Formation of Copper-Zinc Superoxide Dismutase (SOD1) by Editing K63-linked Polyubiquitin Chains. J. Biol. Chem. 2012, 287, 28576–28585. [Google Scholar] [CrossRef] [PubMed]
- Nanduri, P.; Hao, R.; Fitzpatrick, T.; Yao, T.-P. Chaperone-mediated 26S proteasome remodeling facilitates free K63 ubiquitin chain production and aggresome clearance. J. Biol. Chem. 2015, 290, 9455–9464. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, J.J.; Murphy, P.J.M.; Gaillard, S.; Zhao, X.; Wu, J.T.; Nicchitta, C.V.; Yoshida, M.; Toft, D.O.; Pratt, W.B.; Yao, T.P. HDAC6 regulates Hsp90 acetylation and chaperone-dependent activation of glucocorticoid receptor. Mol. Cell 2005, 18, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Thrower, J.S.; Hoffman, L.; Rechsteiner, M.; Pickart, C.M. Recognition of the polyubiquitin proteolytic signal. EMBO J. 2000, 19, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Lee, E.; Jeon, S.-A.; Yu, J.-W. Histone deacetylase 6 negatively regulates NLRP3 inflammasome activation. Biochem. Biophys. Res. Commun. 2015, 467, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J.; Lee, H.-C.; Kim, J.-H.; Park, S.Y.; Kim, T.-H.; Lee, W.-K.; Jang, D.J.; Yoon, J.E.; Choi, Y.I.; Kim, S.; et al. HDAC6 regulates cellular viral RNA sensing by deacetylation of RIG-I. EMBO J. 2016, 35, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wang, Z.; Liu, X.; Yin, C.; Basit, Z.; Xia, B.; Liu, W. Dissection of influenza A virus M1 protein: PH-dependent oligomerization of N-terminal domain and dimerization of C-terminal domain. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Seigneurin-Berny, D.; Verdel, A.; Curtet, S.; Lemercier, C.; Garin, J.; Rousseaux, S.; Khochbin, S. Identification of components of the murine histone deacetylase 6 complex: link between acetylation and ubiquitination signaling pathways. Mol. Cell. Biol. 2001, 21, 8035–8044. [Google Scholar] [CrossRef] [PubMed]
- Husain, M.; Harrod, K.S. Influenza A virus-induced caspase-3 cleaves the histone deacetylase 6 in infected epithelial cells. FEBS Lett. 2009, 583, 2517–2520. [Google Scholar] [CrossRef] [PubMed]
- Dompierre, J.P.; Godin, J.D.; Charrin, B.C.; Cordelières, F.P.; King, S.J.; Humbert, S.; Saudou, F. Histone deacetylase 6 inhibition compensates for the transport deficit in Huntington’s disease by increasing tubulin acetylation. J. Neurosci. 2007, 27, 3571–3583. [Google Scholar] [CrossRef] [PubMed]
- Reed, N.A.; Cai, D.; Blasius, T.L.; Jih, G.T.; Meyhofer, E.; Gaertig, J.; Verhey, K.J. Microtubule Acetylation Promotes Kinesin-1 Binding and Transport. Curr. Biol. 2006, 16, 2166–2172. [Google Scholar] [CrossRef] [PubMed]
- Verhey, K.J.; Hammond, J.W. Traffic control: regulation of kinesin motors. Nat. Rev. Mol. Cell Biol. 2009, 10, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.S.; Hubbert, C.C.; Yao, T.P. The microtubule-associated histone deacetylase 6 (HDAC6) regulates epidermal growth factor receptor (EGFR) endocytic trafficking and degradation. J. Biol. Chem. 2010, 285, 11219–11226. [Google Scholar] [CrossRef] [PubMed]
- Momose, F.; Sekimoto, T.; Ohkura, T.; Jo, S.; Kawaguchi, A.; Nagata, K.; Morikawa, Y. Apical transport of influenza A virus ribonucleoprotein requires Rab11-positive recycling endosome. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Husain, M.; Cheung, C.-Y. Histone deacetylase 6 inhibits influenza A virus release by downregulating the trafficking of viral components to the plasma membrane via its substrate acetylated microtubules. J. Virol. 2014, 88, 11229–11239. [Google Scholar] [CrossRef] [PubMed]
- Husain, M.; Harrod, K.S. Enhanced acetylation of alpha-tubulin in influenza A virus infected epithelial cells. FEBS Lett. 2011, 585, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Eisfeld, A.J.; Kawakami, E.; Watanabe, T.; Neumann, G.; Kawaoka, Y. RAB11A is essential for transport of the influenza virus genome to the plasma membrane. J. Virol. 2011, 85, 6117–6126. [Google Scholar] [CrossRef] [PubMed]
- Wileman, T. Aggresomes and Pericentriolar Sites of Virus Assembly: Cellular Defense or Viral Design? Annu. Rev. Microbiol. 2007, 61, 149–167. [Google Scholar] [CrossRef] [PubMed]
- Nusinzon, I.; Horvath, C.M. Positive and negative regulation of the innate antiviral response and beta interferon gene expression by deacetylation. Mol. Cell. Biol. 2006, 26, 3106–3113. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Pillai, P.S. Innate immunity to influenza virus infection. Nat. Rev. Immunol. 2014, 14, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.; Thomsen, A.R. Sensing of RNA Viruses: a Review of Innate Immune Receptors Involved in Recognizing RNA Virus Invasion. J. Virol. 2012, 86, 2900–2910. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Innate immunity to virus infection. Immunol. Rev. 2009, 227, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Yang, Y.-K.; Wang, R.-P.; Zhou, X.; Diao, F.-C.; Li, M.-D.; Zhai, Z.H.; Jiang, Z.F.; Chen, D.Y. REUL Is a Novel E3 Ubiquitin Ligase and Stimulator of Retinoic-Acid-Inducible Gene-I. PLoS ONE 2009, 4, e5760. [Google Scholar] [CrossRef] [PubMed]
- Oshiumi, H.; Matsumoto, M.; Hatakeyama, S.; Seya, T. Riplet/RNF135, a RING Finger Protein, Ubiquitinates RIG-I to Promote Interferon-β Induction during the Early Phase of Viral Infection. J. Biol. Chem. 2009, 284, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Peisley, A.; Wu, B.; Xu, H.; Chen, Z.J.; Hur, S. Structural basis for ubiquitin-mediated antiviral signal activation by RIG-I. Nature 2014, 509, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Kinch, L.N.; Brautigam, C.A.; Chen, X.; Du, F.; Grishin, N.V.; Chen, Z.J. Ubiquitin-induced oligomerization of the RNA sensors RIG-I and MDA5 activates antiviral innate immune response. Immunity 2012, 36, 959–973. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Peisley, A.; Richards, C.; Yao, H.; Zeng, X.; Lin, C.; Chu, F.; Walz, T.; Hur, S. Structural Basis for dsRNA Recognition, Filament Formation, and Antiviral Signal Activation by MDA5. Cell 2013, 152, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Mänz, B.; Schwemmle, M.; Brunotte, L. Adaptation of avian influenza A virus polymerase in mammals to overcome the host species barrier. J. Virol. 2013, 87, 7200–7209. [Google Scholar] [CrossRef] [PubMed]
- Moncorgé, O.; Mura, M.; Barclay, W. Evidence for avian and human host cell factors that affect the activity of influenza virus polymerase. J. Virol. 2010, 84, 9978–9986. [Google Scholar] [CrossRef] [PubMed]
- Long, J.S.; Giotis, E.S.; Moncorgé, O.; Frise, R.; Mistry, B.; James, J.; Morisson, M.; Iqbal, M.; Vignal, A.; Skinner, M.A.; et al. Species difference in ANP32A underlies influenza A virus polymerase host restriction. Nature 2016, 529, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Miranzo-Navarro, D.; Magor, K.E. Activation of duck RIG-I by TRIM25 is independent of anchored ubiquitin. PLoS ONE 2014, 9, e86968. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-Y.; Ichinohe, T. Response of host inflammasomes to viral infection. Trends Microbiol. 2015, 23, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, M.; Chen, I.-Y.; Kawaguchi, A.; Koshiba, T.; Nagata, K.; Takeyama, H.; Hasegawa, H.; Ichinohe, T. The RNA- and TRIM25-Binding Domains of Influenza Virus NS1 Protein Are Essential for Suppression of NLRP3 Inflammasome-Mediated Interleukin-1β Secretion. J. Virol. 2016, 90, 4105–4114. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jiang, W.; Yan, Y.; Gong, T.; Han, J.; Tian, Z.; Zhou, R. RNA viruses promote activation of the NLRP3 inflammasome through a RIP1-RIP3-DRP1 signaling pathway. Nat. Immunol. 2014, 15, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Negash, A.A.; Ramos, H.J.; Crochet, N.; Lau, D.T.Y.; Doehle, B.; Papic, N.; Delker, D.A.; Jo, J.; Bertoletti, A.; Hagedorn, C.H.; et al. IL-1β Production through the NLRP3 Inflammasome by Hepatic Macrophages Links Hepatitis C Virus Infection with Liver Inflammation and Disease. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Xu, Y.; Li, H.; Tao, W.; Xiang, Y.; Huang, B.; Niu, J.; Zhong, J.; Meng, G. HCV genomic RNA activates the NLRP3 inflammasome in human myeloid cells. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Juliana, C.; Fernandes-Alnemri, T.; Kang, S.; Farias, A.; Qin, F.; Alnemri, E.S. Non-transcriptional priming and deubiquitination regulate NLRP3 inflammasome activation. J. Biol. Chem. 2012, 287, 36617–36622. [Google Scholar] [CrossRef] [PubMed]
- Py, B.F.; Kim, M.S.; Vakifahmetoglu-Norberg, H.; Yuan, J. Deubiquitination of NLRP3 by BRCC3 Critically Regulates Inflammasome Activity. Mol. Cell 2013, 49, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, M.A.; Bowman, J.W.; Fujita, H.; Orazio, N.; Shi, M.; Liang, Q.; Amatya, R.; Kelly, T.J.; Iwai, K.; Ting, J.; et al. The linear ubiquitin assembly complex (LUBAC) is essential for NLRP3 inflammasome activation. J. Exp. Med. 2014, 211, 1333–1347. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.; Mitoma, H.; Trichot, C.; Bao, M.; Liu, Y.; Zhang, Z.; Liu, Y.J. The E3 ubiquitin ligase tripartite motif 33 is essential for cytosolic RNA-induced NLRP3 inflammasome activation. J. Immunol. 2014, 193, 3676–3682. [Google Scholar] [CrossRef] [PubMed]
- Rajsbaum, R.; Albrecht, R.A.; Wang, M.K.; Maharaj, N.P.; Versteeg, G.A.; Nistal-Villán, E.; García-Sastre, A.; Gack, M.U. Species-Specific Inhibition of RIG-I Ubiquitination and IFN Induction by the Influenza A Virus NS1 Protein. PLoS Pathog. 2012, 8, e1003059. [Google Scholar] [CrossRef] [PubMed]
- Cantin, R.; Methot, S.; Tremblay, M.J. Plunder and Stowaways: Incorporation of Cellular Proteins by Enveloped Viruses. J. Virol. 2005, 79, 6577–6587. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, E.C.; Charles, P.D.; Hester, S.S.; Thomas, B.; Trudgian, D.; Martínez-Alonso, M.; Fodor, E. Conserved and host-specific features of influenza virion architecture. Nat. Commun. 2014, 5, 4816. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.L.; Stone, K.L.; Colangelo, C.M.; Gulcicek, E.E.; Palese, P. Cellular proteins in influenza virus particles. PLoS Pathog. 2008, 4, e1000085. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Sun, E.; Bujny, M.V.; Kim, D.; Davidson, M.W.; Zhuang, X. Dual Function of CD81 in Influenza Virus Uncoating and Budding. PLoS Pathog. 2013, 9, e1003701. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.E.; Gack, M.U. Ubiquitination in the antiviral immune response. Virology 2015, 1–14. [Google Scholar] [CrossRef] [PubMed]
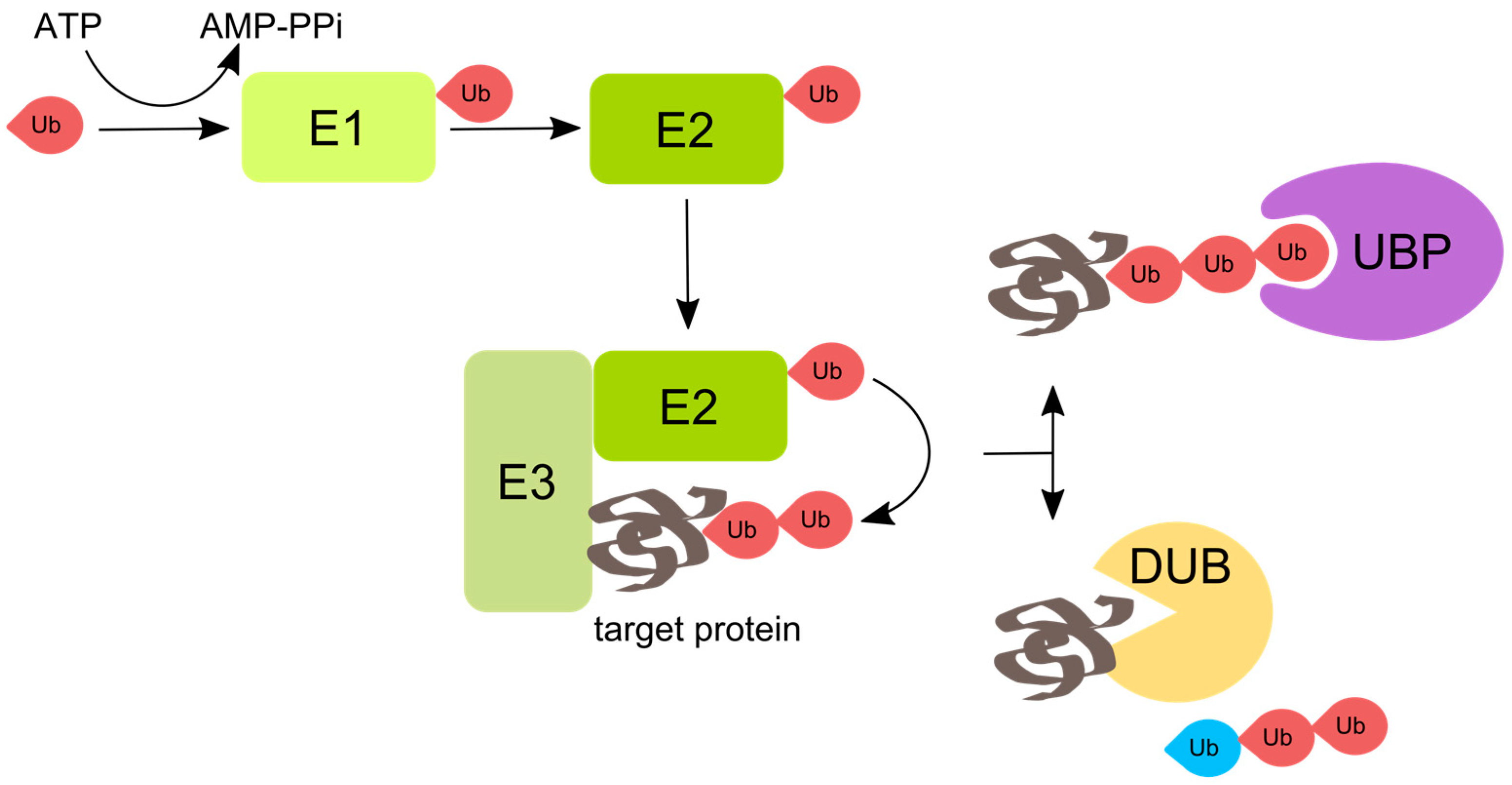
Figure 1.
Enzymatic pathway of protein ubiquitination. The attachment of ubiquitin (Ub) to proteins involves consecutive action of three classes of enzymes: ubiquitin-activating enzyme E1, ubiquitin-conjugating enzyme E2, and ubiquitin ligase E3. First, the C-terminus of ubiquitin binds to E1 in an ATP-dependent manner. Ubiquitin is then transferred from the E1 to E2. Finally, the E3 binds both the ubiquitin-bound E2 and the substrate and catalyzes formation of an isopeptide bond between the C-terminus of ubiquitin and the substrate lysine residue. The lysines on the substrate-conjugated ubiquitin can be further polyubiquitinated. A multitude of cellular proteins that contain different ubiquitin-binding domains—namely ubiquitin binding proteins (UBPs)—mediate the cellular functions of ubiquitination. Deubiquitinases (DUBs) that also contain ubiquitin-binding domains can revert or modify ubiquitination. DUBs vary in the specificity towards different types of polyubiquitin linkages, the position of cleavage within polyubiquitin chains, the ability to separate single ubiquitin moieties, etc. Some DUBs produce unanchored polyubiquitin chains that regulate aggresome processing and innate immunity. The red ubiquitin depicts ubiquitin that is anchored, and the blue ubiquitin depicts unanchored ubiquitin with a free C-terminus.
Figure 1.
Enzymatic pathway of protein ubiquitination. The attachment of ubiquitin (Ub) to proteins involves consecutive action of three classes of enzymes: ubiquitin-activating enzyme E1, ubiquitin-conjugating enzyme E2, and ubiquitin ligase E3. First, the C-terminus of ubiquitin binds to E1 in an ATP-dependent manner. Ubiquitin is then transferred from the E1 to E2. Finally, the E3 binds both the ubiquitin-bound E2 and the substrate and catalyzes formation of an isopeptide bond between the C-terminus of ubiquitin and the substrate lysine residue. The lysines on the substrate-conjugated ubiquitin can be further polyubiquitinated. A multitude of cellular proteins that contain different ubiquitin-binding domains—namely ubiquitin binding proteins (UBPs)—mediate the cellular functions of ubiquitination. Deubiquitinases (DUBs) that also contain ubiquitin-binding domains can revert or modify ubiquitination. DUBs vary in the specificity towards different types of polyubiquitin linkages, the position of cleavage within polyubiquitin chains, the ability to separate single ubiquitin moieties, etc. Some DUBs produce unanchored polyubiquitin chains that regulate aggresome processing and innate immunity. The red ubiquitin depicts ubiquitin that is anchored, and the blue ubiquitin depicts unanchored ubiquitin with a free C-terminus.

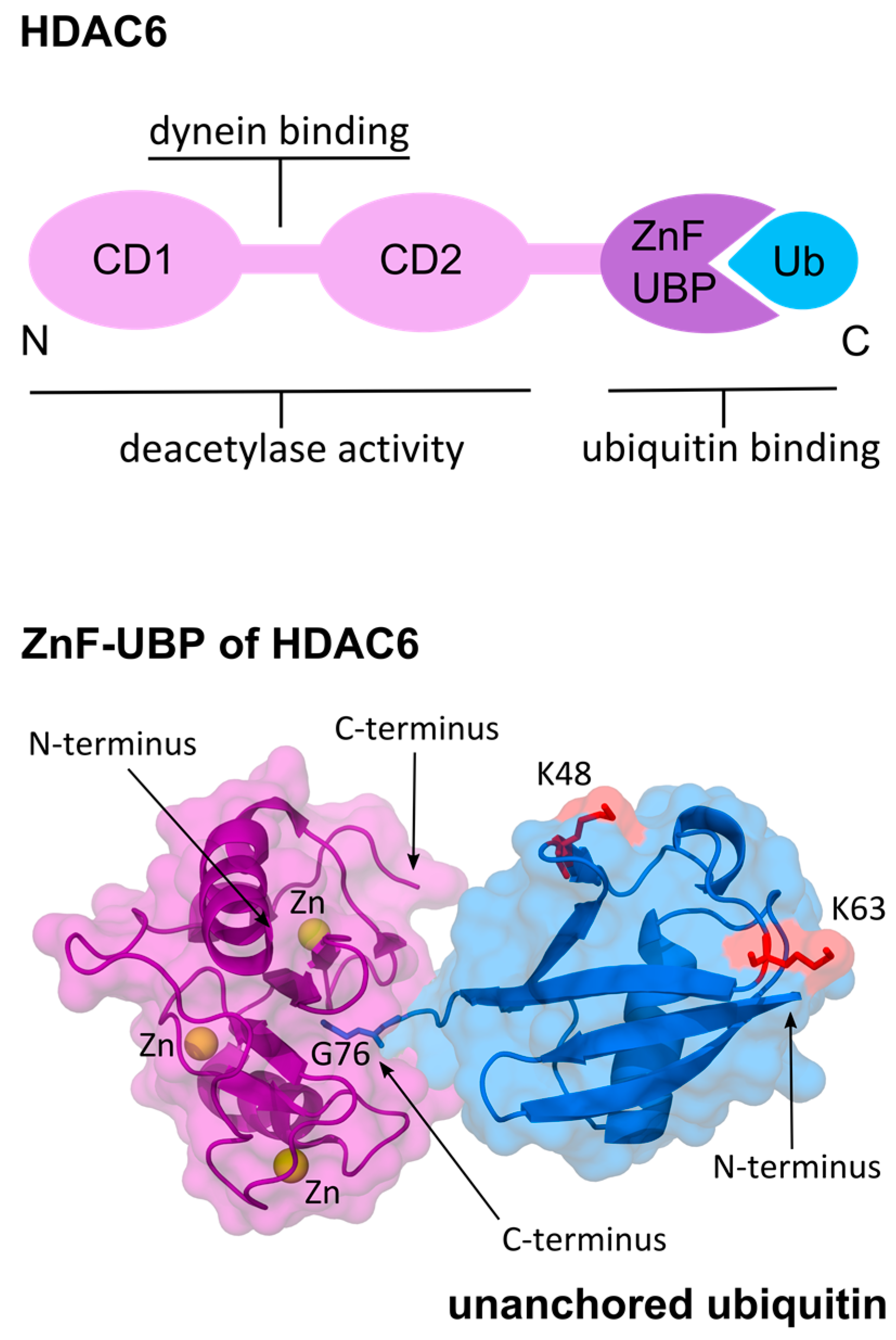
Figure 2.
Functional domains of histone deacetylase 6 (HDAC6) and zinc finger ubiquitin-binding domain (ZnF-UBP) binding to ubiquitin (Ub) C-terminus. Top: Functional domain organization of HDAC6. HDAC6 contains tandem catalytic domains CD1 and CD2, as well as, a ZnF-UBP [63,65,72]. The linker sequence between the two deacetylase domains is responsible for interaction with dynein via p150glued [66,67]; Bottom: Structure of the ZnF-UBP of human HDAC6 bound to ubiquitin (PDB ID: ubq1 [73] and 3gv4 [71]). The ZnF-UBP forms a deep pocket that specifically binds the C-terminal diglycine motif (G75, G76) of unanchored ubiquitin [70]. The coordination of zinc ions (Zn) in the ZnF-UBP, and K48, K63 residues of ubiquitin are shown.
Figure 2.
Functional domains of histone deacetylase 6 (HDAC6) and zinc finger ubiquitin-binding domain (ZnF-UBP) binding to ubiquitin (Ub) C-terminus. Top: Functional domain organization of HDAC6. HDAC6 contains tandem catalytic domains CD1 and CD2, as well as, a ZnF-UBP [63,65,72]. The linker sequence between the two deacetylase domains is responsible for interaction with dynein via p150glued [66,67]; Bottom: Structure of the ZnF-UBP of human HDAC6 bound to ubiquitin (PDB ID: ubq1 [73] and 3gv4 [71]). The ZnF-UBP forms a deep pocket that specifically binds the C-terminal diglycine motif (G75, G76) of unanchored ubiquitin [70]. The coordination of zinc ions (Zn) in the ZnF-UBP, and K48, K63 residues of ubiquitin are shown.

Figure 3.
HDAC6 and unanchored ubiquitin promote aggresome processing and influenza A virus (IAV) uncoating. Left side: HDAC6 regulates the aggresome processing machinery. E3 ubiquitin ligases polyubiquitinate misfolded proteins, and those with a tetra K48-linked ubiquitin chain or longer are degraded by the proteasome [78]. Proteasome inhibition leads to formation of large, misfolded protein aggregates with polyubiquitinated chains, which are cleaved into unanchored ubiquitin chains by DUBs such as Ataxin-3. The C-terminus of ubiquitin is exposed on the unanchored ubiquitin chains and bind to HDAC6 ZnF-UBP. This, by yet unknown mechanisms, activates dynein binding of HDAC6 via its linker region between CD1 and CD2. The unanchored ubiquitin chain(s) is now linked to the dynein motor and microtubules, allowing retrograde transport of the protein aggregate toward the microtubule-organizing center (MTOC) (aggresome formation). Misfolded aggregates that are sequestered to the MTOC eventually form a large complex called the aggresome. HDAC6 promotes autophagy, which involves the actin network, myosin II, and Poh1, a proteasome-associated DUB. Poh1 generates K63-linked unanchored ubiquitin chains that activate HDAC6 (aggresome clearance) [12]. The red ubiquitin (Ub) depicts ubiquitin that is anchored, the blue ubiquitin depicts unanchored ubiquitin with a free C-terminus; Right side: IAV hijacks the aggresome processing machinery during host cell entry. After binding to the cell surface, IAV internalizes by endocytosis, travels to late endosomes (LEs) in the vicinity of the MTOC. In LEs the low pH (5.5–5.0) triggers hemagglutinin (HA) acidification and fusion of the viral envelope with the limiting endosomal membrane. The fusion pore exposes the viral core containing unanchored ubiquitin chains to the cytosol, which recruit HDAC6 and activate the aggresome processing machinery. HDAC6 binds to matrix protein M1, dynein, and myosin, and promotes capsid disassembly by the shearing force of the cytoskeletal motors (box; uncoating). M1 becomes dispersed in the cytosol, the viral ribonucleoproteins (vRNPs) penetrate into the cytosol and are imported into the nucleus through nuclear pore complexes by importin α/β [52,66]. Unanchored ubiquitin carried by IAV might activate HDAC6 similar to aggresome processing [12]. HDAC6, unanchored ubiquitin chains are implicated in retinoic acid-inducible gene 1 (RIG-I) immune signaling and NACHT, LRR and PYD domains-containing protein 3 (NLRP3) inflammasome regulation [13,79,80].
Figure 3.
HDAC6 and unanchored ubiquitin promote aggresome processing and influenza A virus (IAV) uncoating. Left side: HDAC6 regulates the aggresome processing machinery. E3 ubiquitin ligases polyubiquitinate misfolded proteins, and those with a tetra K48-linked ubiquitin chain or longer are degraded by the proteasome [78]. Proteasome inhibition leads to formation of large, misfolded protein aggregates with polyubiquitinated chains, which are cleaved into unanchored ubiquitin chains by DUBs such as Ataxin-3. The C-terminus of ubiquitin is exposed on the unanchored ubiquitin chains and bind to HDAC6 ZnF-UBP. This, by yet unknown mechanisms, activates dynein binding of HDAC6 via its linker region between CD1 and CD2. The unanchored ubiquitin chain(s) is now linked to the dynein motor and microtubules, allowing retrograde transport of the protein aggregate toward the microtubule-organizing center (MTOC) (aggresome formation). Misfolded aggregates that are sequestered to the MTOC eventually form a large complex called the aggresome. HDAC6 promotes autophagy, which involves the actin network, myosin II, and Poh1, a proteasome-associated DUB. Poh1 generates K63-linked unanchored ubiquitin chains that activate HDAC6 (aggresome clearance) [12]. The red ubiquitin (Ub) depicts ubiquitin that is anchored, the blue ubiquitin depicts unanchored ubiquitin with a free C-terminus; Right side: IAV hijacks the aggresome processing machinery during host cell entry. After binding to the cell surface, IAV internalizes by endocytosis, travels to late endosomes (LEs) in the vicinity of the MTOC. In LEs the low pH (5.5–5.0) triggers hemagglutinin (HA) acidification and fusion of the viral envelope with the limiting endosomal membrane. The fusion pore exposes the viral core containing unanchored ubiquitin chains to the cytosol, which recruit HDAC6 and activate the aggresome processing machinery. HDAC6 binds to matrix protein M1, dynein, and myosin, and promotes capsid disassembly by the shearing force of the cytoskeletal motors (box; uncoating). M1 becomes dispersed in the cytosol, the viral ribonucleoproteins (vRNPs) penetrate into the cytosol and are imported into the nucleus through nuclear pore complexes by importin α/β [52,66]. Unanchored ubiquitin carried by IAV might activate HDAC6 similar to aggresome processing [12]. HDAC6, unanchored ubiquitin chains are implicated in retinoic acid-inducible gene 1 (RIG-I) immune signaling and NACHT, LRR and PYD domains-containing protein 3 (NLRP3) inflammasome regulation [13,79,80].

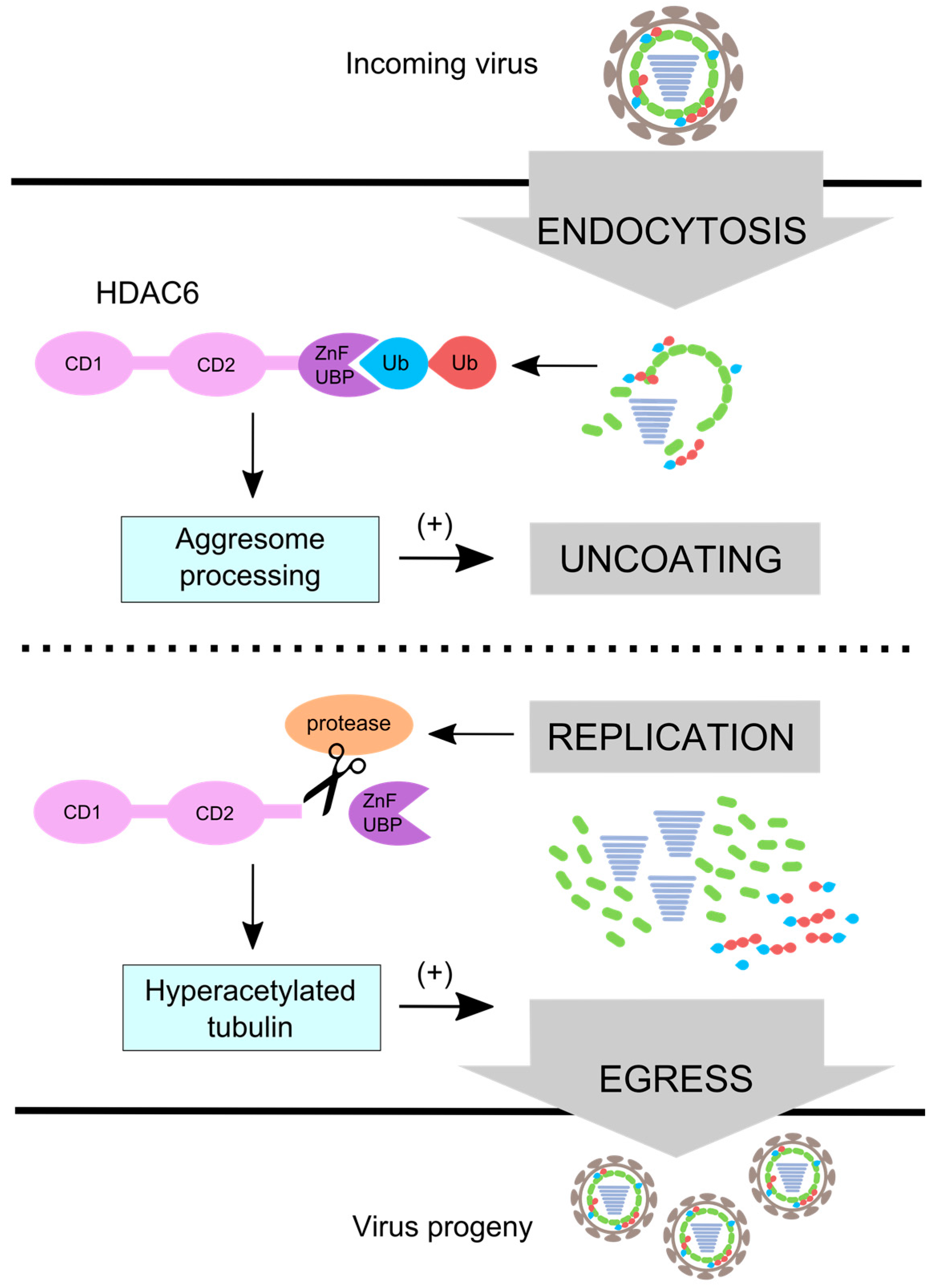
Figure 4.
IAV uses HDAC6 differentially during entry and egress. IAV uses aggresome processing and unanchored ubiquitin chains in an HDAC6 ZnF-UBP-dependent manner to promote capsid uncoating during entry. During replication IAV induces ZnF-UBP cleavage by caspase 3 [83] which inactivates virus uncoating activity. Microtubules become hyperacetylated [59] and promote viral egress [90]. The red ubiquitin (Ub) depicts ubiquitin that is anchored, and the blue ubiquitin depicts unanchored ubiquitin with a free C-terminus.
Figure 4.
IAV uses HDAC6 differentially during entry and egress. IAV uses aggresome processing and unanchored ubiquitin chains in an HDAC6 ZnF-UBP-dependent manner to promote capsid uncoating during entry. During replication IAV induces ZnF-UBP cleavage by caspase 3 [83] which inactivates virus uncoating activity. Microtubules become hyperacetylated [59] and promote viral egress [90]. The red ubiquitin (Ub) depicts ubiquitin that is anchored, and the blue ubiquitin depicts unanchored ubiquitin with a free C-terminus.

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rudnicka, A.; Yamauchi, Y. Ubiquitin in Influenza Virus Entry and Innate Immunity. Viruses 2016, 8, 293. https://0-doi-org.brum.beds.ac.uk/10.3390/v8100293
AMA Style
Rudnicka A, Yamauchi Y. Ubiquitin in Influenza Virus Entry and Innate Immunity. Viruses. 2016; 8(10):293. https://0-doi-org.brum.beds.ac.uk/10.3390/v8100293
Chicago/Turabian StyleRudnicka, Alina, and Yohei Yamauchi. 2016. "Ubiquitin in Influenza Virus Entry and Innate Immunity" Viruses 8, no. 10: 293. https://0-doi-org.brum.beds.ac.uk/10.3390/v8100293
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.