
Three-Dimensional Rotating Wall Vessel-Derived Cell Culture Models for Studying Virus-Host Interactions


Abstract
:1. Introduction
2. Virus Infection and Replication in 3D Bioreactor Models
3. Three-Dimensional Models of Human Neuronal Cells to Study Persistent Viral Infections
4. Three-Dimensional Models of Lymphoid Tissue and Circulating Lymphocytes for Long-Term Culture to Study Virus Replication, Latency and Reactivation
5. Three-Dimensional Models for Respiratory Viral Infections
6. Three-Dimensional Models for Studying Viral Gastroenteritis
7. Three-Dimensional Models of Liver Tissue for Studying Hepatitis Viral Infections
8. Three-Dimensional Models of Female Reproductive Tract Tissues for Studying Sexually-Transmitted Infections and Emerging Viruses
9. Future Directions for RWV Bioreactors
9.1. Considering the Site-Specific Microbiome for the Advancement of RWV-Derived Tissue Models
9.2. Advancing RWV Bioreactor Tissue Models with Enhanced Cellular Complexity
10. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Abbott, A. Cell culture: Biology’s new dimension. Nature 2003, 424, 870–872. [Google Scholar] [CrossRef] [PubMed]
- Nickerson, C.A.; Richter, E.G.; Ott, C.M. Studying host-pathogen interactions in 3D: Organotypic models for infectious disease and drug development. J. Neuroimmune Pharmacol. 2007, 2, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S. Beyond the petri dish. Nat. Biotechnol. 2004, 22, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Torres-Flores, J.M.; Arias, C.F. Tight junctions go viral! Viruses 2015, 7, 5145–5154. [Google Scholar] [PubMed]
- Cohen, M.; Zhang, X.Q.; Senaati, H.P.; Chen, H.W.; Varki, N.M.; Schooley, R.T.; Gagneux, P. Influenza a penetrates host mucus by cleaving sialic acids with neuraminidase. Virol. J. 2013, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- Zanin, M.; Baviskar, P.; Webster, R.; Webby, R. The interaction between respiratory pathogens and mucus. Cell Host Microbe 2016, 19, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Zanin, M.; Marathe, B.; Wong, S.S.; Yoon, S.W.; Collin, E.; Oshansky, C.; Jones, J.; Hause, B.; Webby, R. Pandemic swine H1N1 influenza viruses with almost undetectable neuraminidase activity are not transmitted via aerosols in ferrets and are inhibited by human mucus but not swine mucus. J. Virol. 2015, 89, 5935–5948. [Google Scholar] [CrossRef] [PubMed]
- Carrel, A. On the permanent life of tissues outside of the organism. J. Exp. Med. 1912, 15, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Strangeways, T.P. The Technique of Tissue Culture ‘In Vitro’; Heffer and Sons: Cambridge, UK, 1924. [Google Scholar]
- Grivel, J.C.; Margolis, L. Use of human tissue explants to study human infectious agents. Nat. Protoc. 2009, 4, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Haycock, J.W. 3D cell culture: A review of current approaches and techniques. Methods Mol. Biol. 2011, 695, 1–15. [Google Scholar] [PubMed]
- Li, X.J.; Valadez, A.V.; Zuo, P.; Nie, Z. Microfluidic 3D cell culture: Potential application for tissue-based bioassays. Bioanalysis 2012, 4, 1509–1525. [Google Scholar] [CrossRef] [PubMed]
- van Duinen, V.; Trietsch, S.J.; Joore, J.; Vulto, P.; Hankemeier, T. Microfluidic 3D cell culture: From tools to tissue models. Curr. Opin. Biotechnol. 2015, 35, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, T.J.; Schroeder, W.F.; Wolf, D.A.; Moyer, M.P. Rotating-wall vessel coculture of small intestine as a prelude to tissue modeling: Aspects of simulated microgravity. Proc. Soc. Exp. Biol. Med. 1993, 202, 181–192. [Google Scholar]
- Schwarz, R.P.; Goodwin, T.J.; Wolf, D.A. Cell culture for three-dimensional modeling in rotating-wall vessels: An application of simulated microgravity. J. Tissue Cult. Methods 1992, 14, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Barrila, J.; Radtke, A.L.; Crabbe, A.; Sarker, S.F.; Herbst-Kralovetz, M.M.; Ott, C.M.; Nickerson, C.A. Organotypic 3d cell culture models: Using the rotating wall vessel to study host-pathogen interactions. Nat. Rev. Microbiol. 2010, 8, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Barzegari, A.; Saei, A.A. An update to space biomedical research: Tissue engineering in microgravity bioreactors. BioImpacts 2012, 2, 23–32. [Google Scholar] [PubMed]
- Grimm, D.; Wehland, M.; Pietsch, J.; Aleshcheva, G.; Wise, P.; van Loon, J.; Ulbrich, C.; Magnusson, N.E.; Infanger, M.; Bauer, J. Growing tissues in real and simulated microgravity: New methods for tissue engineering. Tissue Eng. Part B Rev. 2014, 20, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Navran, S. The application of low shear modeled microgravity to 3D cell biology and tissue engineering. Biotechnol. Annu. Rev. 2008, 14, 275–296. [Google Scholar] [PubMed]
- Cherry, R.S.; Papoutsakis, E.T. Physical mechanisms of cell damage in microcarrier cell culture bioreactors. Biotechnol. Bioeng. 1988, 32, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Ayyaswamy, P.S.; Ducheyne, P. Dynamics of a microcarrier particle in the simulated microgravity environment of a rotating-wall vessel. Microgravity Sci. Technol. 1997, 10, 154–165. [Google Scholar] [PubMed]
- Goodwin, T.J.; McCarthy, M.; Cohrs, R.J.; Kaufer, B.B. 3D tissue-like assemblies: A novel approach to investigate virus-cell interactions. Methods 2015, 90, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Jessup, J.M.; Brown, K.; Ishii, S.; Ford, R.; Goodwin, T.J.; Spaulding, G. Simulated microgravity does not alter epithelial cell adhesion to matrix and other molecules. Adv. Space Res. 1994, 14, 71–76. [Google Scholar] [CrossRef]
- Drummond, C.G.; Nickerson, C.A.; Coyne, C.B. A three-dimensional cell culture model to study enterovirus infection of polarized intestinal epithelial cells. mSphere 2016, 1, pii: e00030-15. [Google Scholar] [CrossRef] [PubMed]
- Hjelm, B.E.; Berta, A.N.; Nickerson, C.A.; Arntzen, C.J.; Herbst-Kralovetz, M.M. Development and characterization of a three-dimensional organotypic human vaginal epithelial cell model. Biol. Reprod. 2010, 82, 617–627. [Google Scholar] [CrossRef] [PubMed]
- McConkey, C.A.; Delorme-Axford, E.; Nickerson, C.A.; Kim, K.S.; Sadovsky, Y.; Boyle, J.P.; Coyne, C.B. A three-dimensional culture system recapitulates placental syncytiotrophoblast development and microbial resistance. Sci. Adv. 2016, 2, e1501462. [Google Scholar] [CrossRef] [PubMed]
- Radtke, A.L.; Herbst-Kralovetz, M.M. Culturing and applications of rotating wall vessel bioreactor derived 3d epithelial cell models. J. Vis. Exp. 2012. [Google Scholar] [CrossRef] [PubMed]
- Radtke, A.L.; Quayle, A.J.; Herbst-Kralovetz, M.M. Microbial products alter the expression of membrane-associated mucin and antimicrobial peptides in a three-dimensional human endocervical epithelial cell model. Biol. Reprod. 2012, 87, 132. [Google Scholar] [CrossRef] [PubMed]
- Zwezdaryk, K.J.; Warner, J.A.; Machado, H.L.; Morris, C.A.; Honer zu Bentrup, K. Rotating cell culture systems for human cell culture: Human trophoblast cells as a model. J. Vis. Exp. 2012, 59, pii: 3367. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, T.J.; Lin, Y.-H.; McCarthy, M.; Chen, W.; Miller, L.Z.; Quiroz, J.; Nowak, B.M.; Lerch, R.A.; Udem, S.A.; Deatly, A.M. Paramyxovirus infection mimics in vivo cellular dynamics in 3D human broncho-epitlial tissue-like assemblies. NASA Tech. Pap. 2012, 217363. [Google Scholar]
- Goodwin, T.J.; McCarthy, M.; Osterrieder, N.; Cohrs, R.J.; Kaufer, B.B. Three-dimensional normal human neural progenitor tissue-like assemblies: A model of persistent varicella-zoster virus infection. PLoS Pathog. 2013, 9, e1003512. [Google Scholar] [CrossRef] [PubMed]
- Margolis, L.B.; Fitzgerald, W.; Glushakova, S.; Hatfill, S.; Amichay, N.; Baibakov, B.; Zimmerberg, J. Lymphocyte trafficking and HIV infection of human lymphoid tissue in a rotating wall vessel bioreactor. AIDS Res. Hum. Retrovir. 1997, 13, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Long, J.P.; Pierson, S.; Hughes, J.H. Suppression of Epstein-Barr virus reactivation in lymphoblastoid cells cultured in simulated microgravity. In Vitro Cell. Dev. Biol. Anim. 1999, 35, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Long, J.P.; Hughes, J.H. Epstein-Barr virus latently infected cells are selectively deleted in simulated-microgravity cultures. In Vitro Cell. Dev. Biol. Anim. 2001, 37, 223–230. [Google Scholar] [PubMed]
- Brinley, A.A.; Theriot, C.A.; Nelman-Gonzalez, M.; Crucian, B.; Stowe, R.P.; Barrett, A.D.; Pierson, D.L. Characterization of Epstein-Barr virus reactivation in a modeled spaceflight system. J. Cell. Biochem. 2013, 114, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Suderman, M.T.; McCarthy, M.; Mossell, M.; Watts, D.M.; Peters, C.J.; Shope, R.; Goodwin, T.J. Three-dimensional human bronchial-tracheal epithelial tissue-like assemblies as hosts for severe acute respiratory syndrome-CoV infection. NASA Tech. Pap. 2006, 213723. [Google Scholar]
- Goodwin, T.J.; McCarthy, M.; Lin, Y.-H.; Deatly, A.M. Three-dimensionally engineered normal human lung tissue-like assemblies: Target tissues for human respiratory viral infections. NASA Tech. Pap. 2008, 214771. [Google Scholar]
- Sainz, B., Jr.; TenCate, V.; Uprichard, S.L. Three-dimensional Huh7 cell culture system for the study of hepatitis c virus infection. Virol. J. 2009, 6, 103. [Google Scholar] [CrossRef] [PubMed]
- Berto, A.; Van der Poel, W.H.; Hakze-van der Honing, R.; Martelli, F.; La Ragione, R.M.; Inglese, N.; Collins, J.; Grierson, S.; Johne, R.; Reetz, J.; Dastjerdi, A.; Banks, M. Replication of hepatitis E virus in three-dimensional cell culture. J. Virol. Methods 2013, 187, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Straub, T.M.; Honer zu Bentrup, K.; Orosz-Coghlan, P.; Dohnalkova, A.; Mayer, B.K.; Bartholomew, R.A.; Valdez, C.O.; Bruckner-Lea, C.J.; Gerba, C.P.; Abbaszadegan, M.; et al. In vitro cell culture infectivity assay for human noroviruses. Emerg. Infect. Dis. 2007, 13, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Herbst-Kralovetz, M.M.; Radtke, A.L.; Lay, M.K.; Hjelm, B.E.; Bolick, A.N.; Sarker, S.S.; Atmar, R.L.; Kingsley, D.H.; Arntzen, C.J.; Estes, M.K.; Nickerson, C.A. Lack of norovirus replication and histo-blood group antigen expression in 3-dimensional intestinal epithelial cells. Emerg. Infect. Dis. 2013, 19, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Papafragkou, E.; Hewitt, J.; Park, G.W.; Greening, G.; Vinje, J. Challenges of culturing human norovirus in three-dimensional organoid intestinal cell culture models. PLoS ONE 2014, 8, e63485. [Google Scholar] [CrossRef] [PubMed]
- Takanashi, S.; Saif, L.J.; Hughes, J.H.; Meulia, T.; Jung, K.; Scheuer, K.A.; Wang, Q. Failure of propagation of human norovirus in intestinal epithelial cells with microvilli grown in three-dimensional cultures. Arch. Virol. 2014, 159, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Straub, T.M.; Bartholomew, R.A.; Valdez, C.O.; Valentine, N.B.; Dohnalkova, A.; Ozanich, R.M.; Bruckner-Lea, C.J.; Call, D.R. Human norovirus infection of Caco-2 cells grown as a three-dimensional tissue structure. J. Water Health 2011, 9, 225. [Google Scholar] [CrossRef] [PubMed]
- Swan, K.F.; Ferris, M.; Pridjian, G.; Morris, C.A.; Sullivan, D.E. In vitro model of placental trophoblast differentiation and cytomegalovirus infection. Pregnancy Hypertens. 2015, 74, 6808–6820. [Google Scholar]
- Lee, N.P.; Luk, J.M. Hepatic tight junctions: From viral entry to cancer metastasis. World J. Gastroenterol. 2010, 16, 289–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina-Jimenez, F.; Benedicto, I.; Dao Thi, V.L.; Gondar, V.; Lavillette, D.; Marin, J.J.; Briz, O.; Moreno-Otero, R.; Aldabe, R.; Baumert, T.F.; et al. Matrigel-embedded 3D culture of Huh-7 cells as a hepatocyte-like polarized system to study hepatitis C virus cycle. Virology 2012, 425, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Ishii, K.; Ishihara, Y.; Yoshizaki, S.; Tanaka, K.; Gotoh, Y.; Aizaki, H.; Kohara, M.; Yoshioka, H.; Mori, Y.; et al. Production of infectious hepatitis c virus particles in three-dimensional cultures of the cell line carrying the genome-length dicistronic viral rna of genotype 1b. Virology 2006, 351, 381–392. [Google Scholar] [CrossRef] [PubMed]
- David, J.; Sayer, N.M.; Sarkar-Tyson, M. The use of a three-dimensional cell culture model to investigate host-pathogen interactions of francisella tularensis in human lung epithelial cells. Microbes Infect. 2014, 16, 735–745. [Google Scholar] [CrossRef] [PubMed]
- McGowin, C.L.; Radtke, A.L.; Abraham, K.; Martin, D.H.; Herbst-Kralovetz, M. Mycoplasma genitalium infection activates cellular host defense and inflammation pathways in a 3-dimensional human endocervical epithelial cell model. J. Infect. Dis. 2013, 207, 1857–1868. [Google Scholar] [CrossRef] [PubMed]
- Nickerson, C.A.; Goodwin, T.J.; Terlonge, J.; Ott, C.M.; Buchanan, K.L.; Uicker, W.C.; Emami, K.; LeBlanc, C.L.; Ramamurthy, R.; Clarke, M.S.; et al. Three-dimensional tissue assemblies: Novel models for the study of salmonella enterica serovar typhimurium pathogenesis. Infect. Immun. 2001, 69, 7106–7120. [Google Scholar] [CrossRef] [PubMed]
- Malenovska, H. 3D rotating wall vessel and 2D cell culture of four veterinary virus pathogens: A comparison of virus yields, portions of infectious particles and virus growth curves. J. Virol. Methods 2016, 228, 10–15. [Google Scholar] [CrossRef] [PubMed]
- LaRussa, P.; Steinberg, S.P.; Shapiro, E.; Vazquez, M.; Gershon, A.A. Varicella vaccine revisited. Nat. Med. 2000, 6, 1299–1300. [Google Scholar] [CrossRef] [PubMed]
- Virgin, H.W.; Wherry, E.J.; Ahmed, R. Redefining chronic viral infection. Cell 2009, 138, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Bourdon-Wouters, C.; Merville-Louis, M.P.; Sadzot-Delvaux, C.; Marc, P.; Piette, J.; Delree, P.; Moonen, G.; Rentier, B. Acute and persistent varicella-zoster virus infection of human and murine neuroblastoma cell lines. J. Neurosci. Res. 1990, 26, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Brazeau, E.; Mahalingam, R.; Gilden, D.; Wellish, M.; Kaufer, B.B.; Osterrieder, N.; Pugazhenthi, S. Varicella-zoster virus-induced apoptosis in mewo cells is accompanied by down-regulation of bcl-2 expression. J. Neurovirol. 2010, 16, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Hood, C.; Cunningham, A.L.; Slobedman, B.; Boadle, R.A.; Abendroth, A. Varicella-zoster virus-infected human sensory neurons are resistant to apoptosis, yet human foreskin fibroblasts are susceptible: Evidence for a cell-type-specific apoptotic response. J. Virol. 2003, 77, 12852–12864. [Google Scholar] [CrossRef] [PubMed]
- König, A.; Hömme, C.; Hauröder, B.; Dietrich, A.; Wolff, M.H. The varicella-zoster virus induces apoptosis in vitro in subpopulations of primary human peripheral blood mononuclear cells. Microbes Infect. 2003, 5, 879–889. [Google Scholar] [CrossRef]
- Pugazhenthi, S.; Nair, S.; Velmurugan, K.; Liang, Q.; Mahalingam, R.; Cohrs, R.J.; Nagel, M.A.; Gilden, D. Varicella-zoster virus infection of differentiated human neural stem cells. J. Virol. 2011, 85, 6678–6686. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.I. Epstein-barr virus infection. N. Engl. J. Med. 2000, 343, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Babcock, G.J.; Decker, L.L.; Volk, M.; Thorley-Lawson, D.A. EBV persistence in memory B cells in vivo. Immunity 1998, 9, 395–404. [Google Scholar] [CrossRef]
- Bechler, B.; Cogoli, A.; Cogoli-Greuter, M.; Muller, O.; Hunzinger, E.; Criswell, S.B. Activation of microcarrier-attached lymphocytes in microgravity. Biotech. Bioeng. 1992, 40, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Cogoli, A.; Cogoli-Greuter, M. Activation and proliferation of lymphocytes and other mammalian cells in microgravity. Adv. Space Biol. Med. 1997, 6, 33–79. [Google Scholar] [PubMed]
- Bem, R.A.; Domachowske, J.B.; Rosenberg, H.F. Animal models of human respiratory syncytial virus disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 301, L148–L156. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, F.; Xu, M.; Huang, K.; Zhong, W.; Cai, W.; Yin, Z.; Huang, S.; Deng, Z.; Wei, M.; et al. Description and clinical treatment of an early outbreak of severe acute respiratory syndrome (SARS) in Guangzhou, pr China. J. Med. Microbiol. 2003, 52, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Gretebeck, L.M.; Subbarao, K. Animal models for SARS and MERS coronaviruses. Curr. Opin. Virol. 2015, 13, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Lopman, B.A.; Steele, D.; Kirkwood, C.D.; Parashar, U.D. The vast and varied global burden of norovirus: Prospects for prevention and control. PLoS Med. 2016, 13, e1001999. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.M.; Widdowson, M.A.; Glass, R.I.; Akazawa, K.; Vinje, J.; Parashar, U.D. Systematic literature review of role of noroviruses in sporadic gastroenteritis. Emerg. Infect. Dis. 2008, 14, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the united states--major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Riddle, M.S.; Walker, R.I. Status of vaccine research and development for norovirus. Vaccine 2016, 34, 2895–2899. [Google Scholar] [CrossRef] [PubMed]
- Duizer, E.; Schwab, K.J.; Neill, F.H.; Atmar, R.L.; Koopmans, M.P.; Estes, M.K. Laboratory efforts to cultivate noroviruses. J. Gen. Virol. 2004, 85, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Hutson, A.M.; Atmar, R.L.; Marcus, D.M.; Estes, M.K. Norwalk virus-like particle hemagglutination by binding to H histo-blood group antigens. J. Virol. 2003, 77, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Lavappa, K.S. Survey of atcc stocks of human cell lines for hela contamination. In Vitro 1978, 14, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Nelson-Rees, W.A.; Daniels, D.W.; Flandermeyer, R.R. Cross-contamination of cells in culture. Science 1981, 212, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.K.; Watanabe, M.; Zhu, S.; Graves, C.L.; Keyes, L.R.; Grau, K.R.; Gonzalez-Hernandez, M.B.; Iovine, N.M.; Wobus, C.E.; Vinje, J.; et al. Enteric bacteria promote human and mouse norovirus infection of B cells. Science 2014, 346, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Neill, F.H.; Blutt, S.E.; Zeng, X.L.; Qu, L.; et al. Replication of human noroviruses in stem cell-derived human enteroids. Science 2016, 353, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Hober, D.; Sane, F. Enteroviral pathogenesis of type 1 diabetes. Discov. Med. 2010, 10, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Hober, D.; Sauter, P. Pathogenesis of type 1 diabetes mellitus: Interplay between enterovirus and host. Nat. Rev. Endocrinol. 2010, 6, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Hepatitis e. Available online: http://www.who.int/mediacentre/factsheets/fs280/en/ (accessed on 1 August 2016).
- Takahashi, M.; Tanaka, T.; Azuma, M.; Kusano, E.; Aikawa, T.; Shibayama, T.; Yazaki, Y.; Mizuo, H.; Inoue, J.; Okamoto, H. Prolonged fecal shedding of hepatitis E virus (HEV) during sporadic acute hepatitis E: Evaluation of infectivity of hev in fecal specimens in a cell culture system. J. Clin. Microbiol. 2007, 45, 3671–3679. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Takahashi, M.; Kusano, E.; Okamoto, H. Development and evaluation of an efficient cell-culture system for hepatitis E virus. J. Gen. Virol. 2007, 88, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Mohd Hanafiah, K.; Groeger, J.; Flaxman, A.D.; Wiersma, S.T. Global epidemiology of hepatitis C virus infection: New estimates of age-specific antibody to HCV seroprevalence. Hepatology 2013, 57, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Quayle, A.J. The innate and early immune response to pathogen challenge in the female genital tract and the pivotal role of epithelial cells. J. Reprod. Immunol. 2002, 57, 61–79. [Google Scholar] [CrossRef]
- LaMarca, H.L.; Ott, C.M.; Honer Zu Bentrup, K.; Leblanc, C.L.; Pierson, D.L.; Nelson, A.B.; Scandurro, A.B.; Whitley, G.S.; Nickerson, C.A.; Morris, C.A. Three-dimensional growth of extravillous cytotrophoblasts promotes differentiation and invasion. Placenta 2005, 26, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Winkle, S.M.; Throop, A.L.; Herbst-Kralovetz, M.M. IL-36gamma augments host defense and immune responses in human female reproductive tract epithelial cells. Front. Microbiol. 2016, 7, 955. [Google Scholar] [CrossRef] [PubMed]
- Garland, S.M.; Steben, M. Genital herpes. Best Pract. Res. Clin. Obstet. Gynaecol. 2014, 28, 1098–1110. [Google Scholar] [CrossRef] [PubMed]
- Wald, A.; Zeh, J.; Selke, S.; Warren, T.; Ryncarz, A.J.; Ashley, R.; Krieger, J.N.; Corey, L. Reactivation of genital herpes simplex virus type 2 infection in asymptomatic seropositive persons. N. Engl. J. Med. 2000, 342, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Gabay, C.; Towne, J.E. Regulation and function of interleukin-36 cytokines in homeostasis and pathological conditions. J. Leukoc. Biol. 2015, 97, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Gresnigt, M.S.; van de Veerdonk, F.L. Biology of IL-36 cytokines and their role in disease. Semin. Immunol. 2013, 25, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.A.; Mier-y-Teran-Romero, L.; Reefhuis, J.; Gilboa, S.M.; Hills, S.L. Zika and the risk of microcephaly. N. Engl. J. Med. 2016, 375, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Berkhout, B. With a little help from my enteric microbial friends. Front. Med. 2015, 2, 30. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.A.; Biesbroek, G.; Trzcinski, K.; Sanders, E.A.; Bogaert, D. Viral and bacterial interactions in the upper respiratory tract. PLoS Pathog. 2013, 9, e1003057. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V. Viruses and microbiome alterations. Ann. Am. Thorac. Soc. 2014, 11 Suppl 1, S57–S60. [Google Scholar] [CrossRef] [PubMed]
- Miura, T.; Sano, D.; Suenaga, A.; Yoshimura, T.; Fuzawa, M.; Nakagomi, T.; Nakagomi, O.; Okabe, S. Histo-blood group antigen-like substances of human enteric bacteria as specific adsorbents for human noroviruses. J. Virol. 2013, 87, 9441–9451. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, J.K.; Virgin, H.W. Viral immunity. Transkingdom control of viral infection and immunity in the mammalian intestine. Science 2016, 351, pii: aad5872. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.M.; Pfeiffer, J.K. Viruses and the microbiota. Ann. Rev. Virol. 2014, 1, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Doerflinger, S.Y.; Throop, A.L.; Herbst-Kralovetz, M.M. Bacteria in the vaginal microbiome alter the innate immune response and barrier properties of the human vaginal epithelia in a species-specific manner. J. Infect. Dis. 2014, 209, 1989–1999. [Google Scholar] [CrossRef] [PubMed]
- Herbst-Kralovetz, M.M.; Pyles, R.B.; Ratner, A.J.; Sycuro, L.K.; Mitchell, C. New systems for studying intercellular interactions in bacterial vaginosis. J. Infect. Dis. 2016, 214, S6–S13. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.J.; O’Shaughnessy, T.J.; Kelly, J.; Ma, W. Neural stem cell differentiation in a cell-collagen-bioreactor culture system. Brain Res. Dev. Brain Res. 2004, 153, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Zhu, B.; Zhang, Y.; Jin, Y. Tissue-engineered nerve constructs under a microgravity system for peripheral nerve regeneration. Tissue Eng. Part A 2015, 21, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Fitzgerald, W.; Liu, Q.Y.; O’Shaughnessy, T.J.; Maric, D.; Lin, H.J.; Alkon, D.L.; Barker, J.L. Cns stem and progenitor cell differentiation into functional neuronal circuits in three-dimensional collagen gels. Exp. Neurol. 2004, 190, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Cerwinka, W.H.; Sharp, S.M.; Boyan, B.D.; Zhau, H.E.; Chung, L.W.; Yates, C. Differentiation of human mesenchymal stem cell spheroids under microgravity conditions. Cell Regen. 2012, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Rungarunlert, S.; Ferreira, J.N.; Dinnyes, A. Novel bioreactor platform for scalable cardiomyogenic differentiation from pluripotent stem cell-derived embryoid bodies. Methods Mol. Biol. 2016, 1502, 169–179. [Google Scholar]
- Teo, A.; Mantalaris, A.; Song, K.; Lim, M. A novel perfused rotary bioreactor for cardiomyogenesis of embryonic stem cells. Biotechnol. Lett. 2014, 36, 947–960. [Google Scholar] [CrossRef] [PubMed]
- Vidyasekar, P.; Shyamsunder, P.; Sahoo, S.K.; Verma, R.S. Scaffold-free and scaffold-assisted 3D culture enhances differentiation of bone marrow stromal cells. In Vitro Cell. Dev. Biol. Anim. 2016, 52, 204–217. [Google Scholar] [CrossRef] [PubMed]



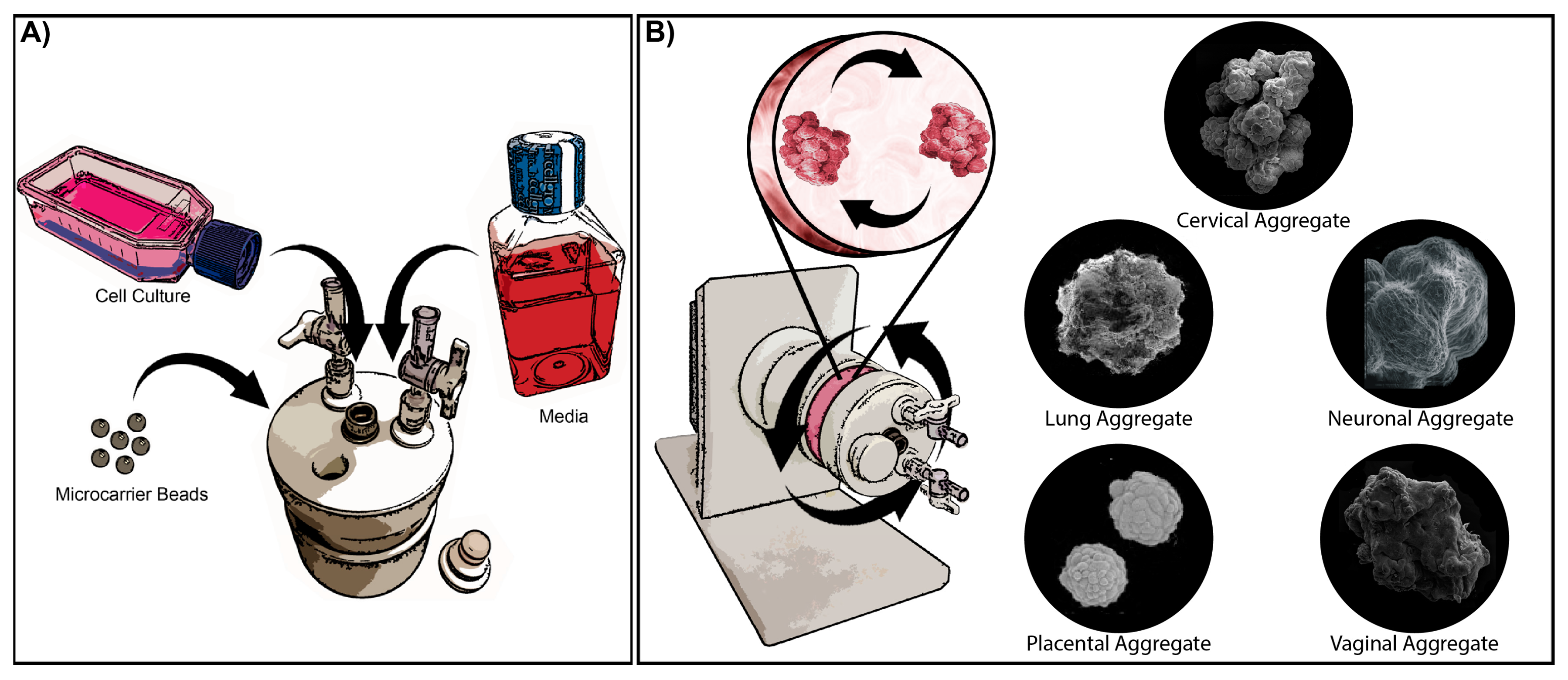{kind=link}
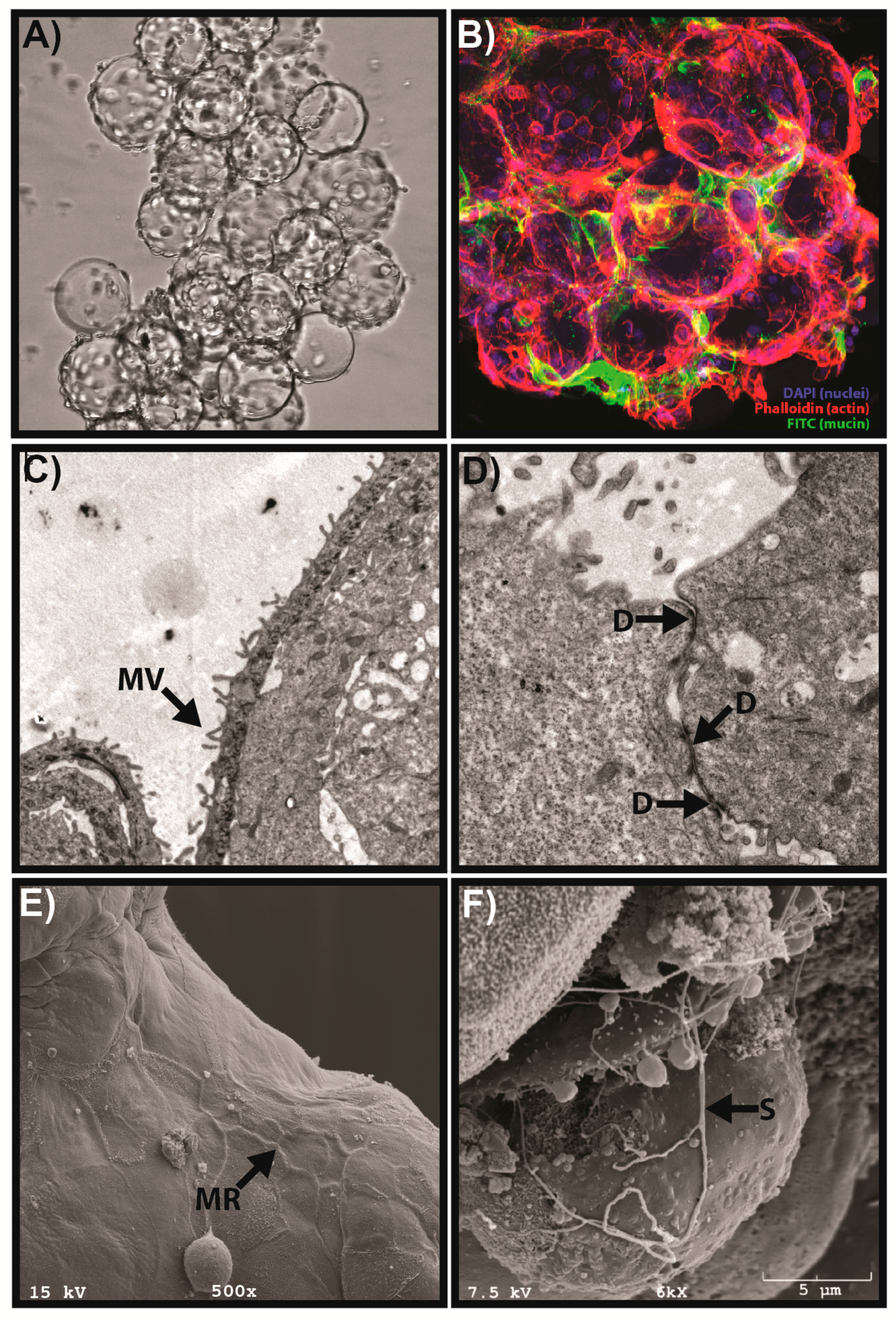{kind=link}
| Tissue Model | Cell Lines | Virus | Virus Replication | Host Response | Reference |
|---|---|---|---|---|---|
| Neuronal | NHNP | VZV | Productive infection | No CPE | [31] |
| Tonsil | Primary cells | HIV | Productive infection | Lymphocyte migration tracked | [32] |
| Lymphoid | P3HR-1 | EBV | Suppression of EBV reactivation | ND | [33] |
| P3HR-1 Daudi Ramos | EBV | Suppression of EBV reactivation | ND | [34] | |
| BJAB Raji | EBV | Suppression of EBV reactivation | Microgravity and radiation increased DNA damage in EBV positive cells | [35] | |
| Lung | HBTC BEAS-2B | SARS-CoV | No SARS-CoV replication detected | Vacuolization, mitochondria loss and chromatin alterations | [36] |
| HBTC BEAS-2B | RSV | Productive infection | Signs of cellular damage; mucus produced in 3D aggregates | [37] | |
| HBTC BEAS-2B | RSV PIV3 | Productive infection | Cytokine profile in 3D aggregates was similar to human airways from RSV- and PIV3-infected patients | [30] | |
| Liver | Huh7 | HCV | Productive infection | Expression and localization of TJ proteins enhance HCV infection | [38] |
| PLC/PRF/5 | HEV | Productive infection | No CPE | [39] | |
| Small Intestine | INT-407 | HuNoV | Increase in viral RNA copies detected | Vacuolization, shortening of apical microvilli, cell detachment from bead | [40] |
| No HuNoV replication detected | Clumping and detachment of cells from microcarrier beads | [41] | |||
| No HuNoV replication detected | No CPE | [42,43] | |||
| Colon | Caco-2 | HuNoV | Increase in HuNoV RNA copies detected | Shortening of apical microvilli and formation of vacuoles | [44] |
| No HuNoV replication detected | No CPE | [42,43] | |||
| CVB | Productive infection | Increased expression of proliferation and differentiation genes | [24] | ||
| Placenta | TBPC | HCMV | Productive infection | ND | [45] |
| JEG-3 | VSV | No VSV replication detected | Resistance to infection mimics in vivo response to infection | [26] | |
| Vagina | V19I | HSV-1 HSV-2 | Productive infection | Mucosal epithelial barrier features mimic in vivo characteristics, aggregates are more resistant to HSV infection | [28] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gardner, J.K.; Herbst-Kralovetz, M.M. Three-Dimensional Rotating Wall Vessel-Derived Cell Culture Models for Studying Virus-Host Interactions. Viruses 2016, 8, 304. https://0-doi-org.brum.beds.ac.uk/10.3390/v8110304
Gardner JK, Herbst-Kralovetz MM. Three-Dimensional Rotating Wall Vessel-Derived Cell Culture Models for Studying Virus-Host Interactions. Viruses. 2016; 8(11):304. https://0-doi-org.brum.beds.ac.uk/10.3390/v8110304
Chicago/Turabian StyleGardner, Jameson K., and Melissa M. Herbst-Kralovetz. 2016. "Three-Dimensional Rotating Wall Vessel-Derived Cell Culture Models for Studying Virus-Host Interactions" Viruses 8, no. 11: 304. https://0-doi-org.brum.beds.ac.uk/10.3390/v8110304