
Israeli Acute Paralysis Virus Infection Leads to an Enhanced RNA Interference Response and Not Its Suppression in the Bumblebee Bombus terrestris


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bumblebee Rearing and Injections
2.2. Virus Production
2.3. dsRNA Synthesis
2.4. RNA Extraction, cDNA Synthesis and RT-qPCR
2.5. RNA Iinterference (RNAi) Efficiency Experiments, RNAi Gene Expression Levels and Pre-Infection Experiments
2.6. Statistical Analysis
2.7. High Definition Mass Spectrometry (HDMSE)
3. Results
3.1. Israeli Acute Paralysis Virus (IAPV) Genome Structure and HDMSE
3.2. Virus Distribution in Bumblebee Tissues
3.3. RNAi Efficiency in Bumblebee Tissues
3.4. Virus Infection Alters RNAi Efficiency
3.5. Expression Levels of Genes Involved in RNAi after Virus Infection
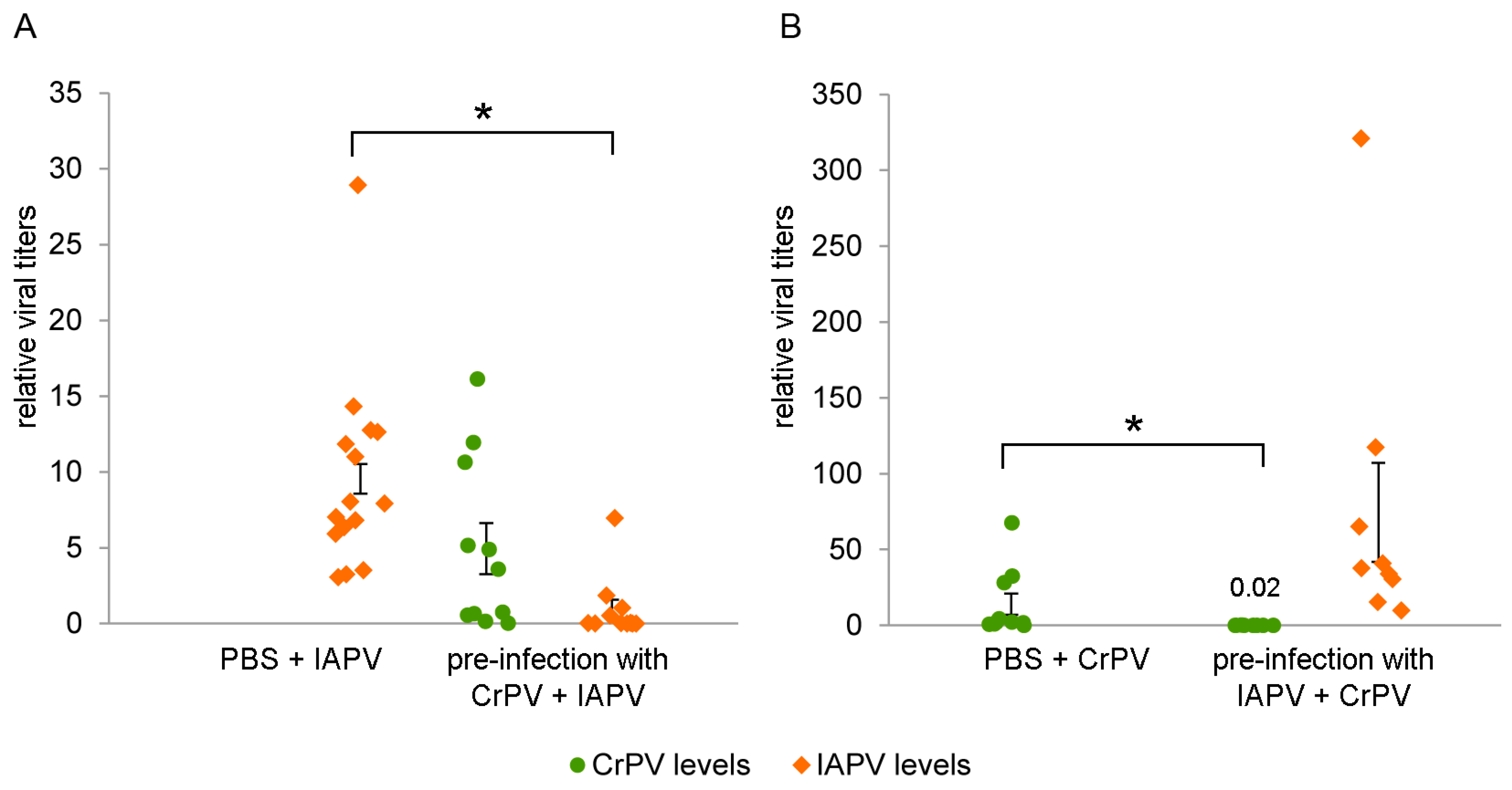
3.6. IAPV/CrPV Levels after Pre-Infection
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nasir, A.; Forterre, P.; Kim, K.M.; Caetano-Anollés, G. The distribution and impact of viral lineages in domains of life. Front. Microbiol. 2014, 5, 194. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.; Mohr, I. Viral subversion of the host protein synthesis machinery. Nat. Rev. Microbiol. 2011, 9, 860–875. [Google Scholar] [CrossRef] [PubMed]
- Samuel, C.E. Antiviral actions of interferons. Clin. Microbiol. Rev. 2001, 14, 778–809. [Google Scholar] [CrossRef] [PubMed]
- Maillard, P.V.; Ciaudo, C.; Marchais, A.; Li, Y.; Jay, F.; Ding, S.W.; Voinnet, O. Antiviral RNA interference in Mammalian cells. Science 2013, 342, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lu, J.; Han, Y.; Fan, X.; Ding, S.-W. RNA interference functions as an antiviral immunity mechanism in Mammals. Science 2013, 342, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Soosaar, J.L.M.; Burch-Smith, T.M.; Dinesh-Kumar, S.P. Mechanisms of plant resistance to viruses. Nat. Rev. Microbiol. 2005, 3, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Karlikow, M.; Goic, B.; Saleh, M.-C. RNAi and antiviral defense in Drosophila: Setting up a systemic immune response. Dev. Comp. Immunol. 2014, 42, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Van Mierlo, J.T.; van Cleef, K.W.R.; van Rij, R.P. Small silencing RNAs: Piecing together a viral genome. Cell Host Microbe 2010, 7, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Marques, J.T.; Wang, J.-P.; Wang, X.; de Oliveira, K.P.V.; Gao, C.; Aguiar, E.R.G.R.; Jafari, N.; Carthew, R.W. Functional specialization of the small interfering RNA pathway in response to virus infection. PLoS Pathog. 2013, 9, e1003579. [Google Scholar] [CrossRef]
- Obbard, D.J.; Gordon, K.H.J.; Buck, A.H.; Jiggins, F.M. The evolution of RNAi as a defence against viruses and transposable elements. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.-C.; Tassetto, M.; van Rij, R.P.; Goic, B.; Gausson, V.; Berry, B.; Jacquier, C.; Antoniewski, C.; Andino, R. Antiviral immunity in Drosophila requires systemic RNA interference spread. Nature 2009, 458, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Burgyán, J.; Havelda, Z. Viral suppressors of RNA silencing. Trends Plant Sci. 2011, 16, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Li, H.W.; Li, W.X.; Ding, S.W. Induction and suppression of RNA silencing by an animal virus. Science 2002, 296, 1319–1321. [Google Scholar] [CrossRef] [PubMed]
- Van Rij, R.P.; Saleh, M.C.; Berry, B.; Foo, C.; Houk, A.; Antoniewski, C.; Andino, R. The RNA silencing endonuclease Argonaute 2 mediates specific antiviral immunity in Drosophila melanogaster. Genes Dev. 2006, 20, 2985–2995. [Google Scholar] [CrossRef] [PubMed]
- Nayak, A.; Berry, B.; Tassetto, M.; Kunitomi, M.; Acevedo, A.; Deng, C.H.; Krutchinsky, A.; Gross, J.; Antoniewski, C.; Andino, R. Cricket paralysis virus antagonizes Argonaute 2 to modulate antiviral defense in Drosophila. Nat. Struct. Mol. Biol. 2010, 17, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Van Mierlo, J.T.; Bronkhorst, A.W.; Overheul, G.J.; Sadanandan, S.A.; Ekstrom, J.O.; Heestermans, M.; Hultmark, D.; Antoniewski, C.; van Rij, R.P. Convergent evolution of argonaute-2 slicer antagonism in two distinct insect RNA viruses. PLoS Pathog. 2012, 8, e1002872. [Google Scholar] [CrossRef] [PubMed]
- Schnettler, E.; Sterken, M.G.; Leung, J.Y.; Metz, S.W.; Geertsema, C.; Goldbach, R.W.; Vlak, J.M.; Kohl, A.; Khromykh, A.A.; Pijlman, G.P. Noncoding flavivirus RNA displays RNA interference suppressor activity in insect and Mammalian cells. J. Virol. 2012, 86, 13486–13500. [Google Scholar] [CrossRef] [PubMed]
- Van Cleef, K.W.R.; van Mierlo, J.T.; Miesen, P.; Overheul, G.J.; Fros, J.J.; Schuster, S.; Marklewitz, M.; Pijlman, G.P.; Junglen, S.; van Rij, R.P. Mosquito and Drosophila entomobirnaviruses suppress dsRNA- and siRNA-induced RNAi. Nucleic Acids Res. 2014, 42, 8732–8744. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Pettis, J.S.; Corona, M.; Chen, W.P.; Li, C.J.; Spivak, M.; Visscher, P.K.; de Grandi-Hoffman, G.; Boncristiani, H.; Zhao, Y.; et al. Israeli acute paralysis virus: Epidemiology, pathogenesis and implications for honey bee health. PLoS Pathog. 2014, 10, e1004261. [Google Scholar] [CrossRef] [PubMed]
- Sabath, N.; Price, N.; Graur, D. A potentially novel overlapping gene in the genomes of Israeli acute paralysis virus and its relatives. Virol. J. 2009, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.E.; Wang, Q.S.; Jan, E.; Atkins, J.F. Bioinformatic evidence for a stem-loop structure 5′-adjacent to the IGR-IRES and for an overlapping gene in the bee paralysis dicistroviruses. Virol. J. 2009, 6, 193. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Wang, Q.S.; Firth, A.E.; Chan, M.M.; Gouw, J.W.; Guarna, M.M.; Foster, L.J.; Atkins, J.F.; Jan, E. Alternative reading frame selection mediated by a tRNA-like domain of an internal ribosome entry site. Proc. Natl. Acad. Sci. USA 2012, 109, E630–E639. [Google Scholar] [CrossRef] [PubMed]
- Valles, S.M.; Sabath, N. No evidence for translation of pog, a predicted overlapping gene of Solenopsis invicta virus 1. Virus Genes 2012, 45, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A Metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Meeus, I.; de Miranda, J.R.; de Graaf, D.C.; Wäckers, F.; Smagghe, G. Effect of oral infection with Kashmir bee virus and Israeli acute paralysis virus on bumblebee (Bombus terrestris) reproductive success. J. Invertebr. Pathol. 2014, 121, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Maori, E.; Paldi, N.; Shafir, S.; Kalev, H.; Tsur, E.; Glick, E.; Sela, I. IAPV, a bee-affecting virus associated with Colony Collapse Disorder can be silenced by dsRNA ingestion. Insect Mol. Biol. 2009, 18, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Plus, N.; Croizier, G.; Reinganum, C.; Scotti, P.D. Cricket paralysis virus and Drosophila C virus: Serological analysis and comparison of capsid polypeptides and host range. J. Invertebr. Pathol. 1978, 31, 296–302. [Google Scholar] [CrossRef]
- Chao, Y.-C.; Young, S.Y., III; Kim, K.S. Characterization of a picornavirus isolated from Pseudoplusia includens (Lepidoptera: Noctuidae). J. Invertebr. Pathol. 1986, 47, 247–257. [Google Scholar] [CrossRef]
- Anderson, D.L.; Gibbs, A.J. Inapparent virus infections and their interactions in Pupae of the Honey Bee (Apis mellifera Linnaeus) in Australia. J. Gen. Virol. 1988, 69, 1617–1625. [Google Scholar] [CrossRef]
- Manley, R.; Boots, M.; Wilfert, L. Emerging viral disease risk to pollinating insects: Ecological, evolutionary and anthropogenic factors. J. Appl. Ecol. 2015, 52, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Tripp, J.; Dolezal, A.G.; Goblirsch, M.J.; Miller, W.A.; Toth, A.L.; Bonning, B.C. In vivo and in vitro infection dynamics of honey bee viruses. Sci. Rep. 2016, 6, 22265. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhao, Y.; Hammond, J.; Hsu, H.T.; Evans, J.; Feldlaufer, M. Multiple virus infections in the honey bee and genome divergence of honey bee viruses. J. Invertebr. Pathol. 2004, 87, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Y.; Jia, H.R.; Wang, Q.; Dai, P.L.; Diao, Q.Y.; Xu, S.F.; Wang, X.; Zhou, T. Multiple Virus Infections and the Characteristics of Chronic Bee Paralysis Virus in Diseased Honey Bees (Apis Mellifera L.) in China. J. Apic. Sci. 2015, 59, 95–106. [Google Scholar] [CrossRef]
- Niu, J.; Cappelle, K.; de Miranda, J.R.; Smagghe, G.; Meeus, I. Analysis of reference gene stability after Israeli acute paralysis virus infection in bumblebees Bombus terrestris. J. Invertebr. Pathol. 2014, 115, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Sguazza, G.H.; Reynaldi, F.J.; Galosi, C.M.; Pecoraro, M.R. Simultaneous detection of bee viruses by multiplex PCR. J. Virol. Methods 2013, 194, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Smagghe, G.; de Coninck, D.I.M.; van Nieuwerburgh, F.; Deforce, D.; Meeus, I. In vivo study of Dicer-2-mediated immune response of the small interfering RNA pathway upon systemic infections of virulent and avirulent viruses in Bombus terrestris. Insect Biochem. Mol. Biol. 2016, 70, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Luo, Y.; Lu, R.; Lau, N.; Lai, E.C.; Li, W.X.; Ding, S.W. Virus discovery by deep sequencing and assembly of virus-derived small silencing RNAs. Proc. Natl. Acad. Sci. USA 2010, 107, 1606–1611. [Google Scholar] [CrossRef] [PubMed]
- Hellemans, J.; Mortier, G.; de Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [PubMed]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Wan, H.; Li, J.; Jin, B.R. Molecular cloning and characterization of a short peptidoglycan recognition protein (PGRP-S) with antibacterial activity from the bumblebee Bombus ignitus. Dev. Comp. Immunol. 2010, 34, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Distler, U.; Kuharev, J.; Navarro, P.; Levin, Y.; Schild, H.; Tenzer, S. Drift time-specific collision energies enable deep-coverage data-independent acquisition proteomics. Nat. Methods 2014, 11, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Swevers, L.; Vanden Broeck, J.; Smagghe, G. The possible impact of persistent virus infection on the function of the RNAi machinery in insects: A hypothesis. Front. Physiol. 2013, 4, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaud, S.; Boncristiani, H.F.; Gouw, J.W.; Strand, M.K.; Pettis, J.; Rueppell, O.; Foster, L.J. Response of the honey bee (Apis mellifera) proteome to Israeli acute paralysis virus (IAPV) infection. Can. J. Zool. 2014, 93, 711–720. [Google Scholar] [CrossRef]
- Wang, Y.; Brent, C.S.; Fennern, E.; Amdam, G.V. Gustatory perception and fat body energy metabolism are jointly affected by vitellogenin and juvenile hormone in honey bees. PLoS Genet. 2012, 8, e1002779. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Grant, G.; Sabin, L.R.; Gordesky-Gold, B.; Yasunaga, A.; Tudor, M.; Cherry, S. Transcriptional pausing controls a rapid antiviral innate immune response in Drosophila. Cell Host Microbe 2012, 12, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Galbraith, D.A.; Yang, X.; Niño, E.L.; Yi, S.; Grozinger, C. Parallel epigenomic and transcriptomic responses to viral infection in honey bees (Apis mellifera). PLoS Pathog. 2015, 11, e1004713. [Google Scholar] [CrossRef] [PubMed]
- Girard, Y.A.; Klingler, K.A.; Higgs, S. West Nile virus dissemination and tissue tropisms in orally infected Culex pipiens quinquefasciatus. Vector Borne Zoonotic Dis. 2004, 4, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Salazar, M.I.; Richardson, J.H.; Sánchez-Vargas, I.; Olson, K.E.; Beaty, B.J. Dengue virus type 2: Replication and tropisms in orally infected Aedes aegypti mosquitoes. BMC Microbiol. 2007, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, J.P.; Kanost, M.R.; Trenczek, T. Biological mediators of insect immunity. Annu. Rev. Entomol. 1997, 42, 611–643. [Google Scholar] [CrossRef] [PubMed]
- Boisson, B.; Jacques, J.C.; Choumet, V.; Martin, E.; Xu, J.; Vernick, K.; Bourgouin, C. Gene silencing in mosquito salivary glands by RNAi. FEBS Lett. 2006, 580, 1988–1992. [Google Scholar] [CrossRef] [PubMed]
- Terenius, O.; Papanicolaou, A.; Garbutt, J.S.; Eleftherianos, I.; Huvenne, H.; Kanginakudru, S.; Albrechtsen, M.; An, C.; Aymeric, J.L.; Barthel, A.; et al. RNA interference in Lepidoptera: An overview of successful and unsuccessful studies and implications for experimental design. J. Insect Physiol. 2011, 57, 231–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynant, N.; Verlinden, H.; Breugelmans, B.; Simonet, G.; Vanden Broeck, J. Tissue-dependence and sensitivity of the systemic RNA interference response in the desert locust, Schistocerca gregaria. Insect Biochem. Mol. Biol. 2012, 42, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Jarosch, A.; Moritz, R.F. Systemic RNA-interference in the honeybee Apis mellifera: Tissue dependent uptake of fluorescent siRNA after intra-abdominal application observed by laser-scanning microscopy. J. Insect Physiol. 2011, 57, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Cai, Z.; Song, J.; Wu, Z.; Zhou, S. dsRNA uptake and persistence account for tissue-dependent susceptibility to RNA interference in the migratory locust. Locusta migratoria. Insect Mol. Biol. 2014, 23, 175–184. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′–3′) | Length (bp) | Eff. (%) |
|---|---|---|---|
| IAPV_q F | CCATGCCTGGCGATTCAC | 203 | 97–102 |
| IAPV_q R | CTGAATAATACTGTGCGTATC | ||
| CrPV_q F | AAACGCAAAAACAGCGAAAC | 110 | 103–104 |
| CrPV_q R | CACATCAAGCACCAAAGCAT | ||
| rpl23_q F | GGGAAAACCTGAACTTAGGAAAA | 143 | 86–99 |
| rpl23_q R | ACCCTTTCATTTCTCCCTTGTTA | ||
| ubi_q F | GGTATTTGGATGCCAGTGATTT | 129 | 94–96 |
| ubi_q R | ATGGGCATTTCTACCCCTTTTA | ||
| ppia_q F | TCGTAATGGAGTTGAGGAGTGA | 132 | 84–94 |
| ppia_q R | CTTGGCACATGAAGTTTGGAAT | ||
| dcr-2_q F | TGGTCAAAACATCAAGAACAACCA | 211 | 93–97 |
| dcr-2_q R | GATCGGGGCCATACGAACAT | ||
| ago-2_q F | CCGAATGTGGACAATGCTTA | 181 | 95–102 |
| ago-2_q R | AACGGGCAAAGGTGTGATTA | ||
| sid-1_q F | CGAGCCCATCAACGGTAGAA | 160 | 94–107 |
| sid-1_q R | CGAGCCAAATCACAAACGGA | ||
| ninaC_q F | GCGAAACCATCTGGAGGATA | 112 | 91–106 |
| ninaC_q R | ACTCTGTTAGCCGCATCGTT | ||
| egghead_q F | ACCGGAGGACTTAGTTGGAA | 122 | 93–97 |
| egghead_q R | TGCGGAAAGGAAAGAAATGT | ||
| GFP_T7 F | TAATACGACTCACTATAGGGTACGGCGTGCAGTGCT | 495 | / |
| GFP_T7 R | TAATACGACTCACTATAGGGTGATCGCGCTTCTCG | ||
| ppia_T7 F | TAATACGACTCACTATAGGGCACTGGTGGAAGGTCCATCT | 388 | / |
| ppia_T7 F | TAATACGACTCACTATAGGGAAGGGAAAATGGTGATGATTAGAA |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappelle, K.; Smagghe, G.; Dhaenens, M.; Meeus, I. Israeli Acute Paralysis Virus Infection Leads to an Enhanced RNA Interference Response and Not Its Suppression in the Bumblebee Bombus terrestris. Viruses 2016, 8, 334. https://0-doi-org.brum.beds.ac.uk/10.3390/v8120334
Cappelle K, Smagghe G, Dhaenens M, Meeus I. Israeli Acute Paralysis Virus Infection Leads to an Enhanced RNA Interference Response and Not Its Suppression in the Bumblebee Bombus terrestris. Viruses. 2016; 8(12):334. https://0-doi-org.brum.beds.ac.uk/10.3390/v8120334
Chicago/Turabian StyleCappelle, Kaat, Guy Smagghe, Maarten Dhaenens, and Ivan Meeus. 2016. "Israeli Acute Paralysis Virus Infection Leads to an Enhanced RNA Interference Response and Not Its Suppression in the Bumblebee Bombus terrestris" Viruses 8, no. 12: 334. https://0-doi-org.brum.beds.ac.uk/10.3390/v8120334