
Long Terminal Repeat Circular DNA as Markers of Active Viral Replication of Human T Lymphotropic Virus-1 in Vivo

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. In-Vivo Studies
2.3. DNA Extraction and Amplification
2.4. PCR Reaction Protocol
2.5. Quantification of LTR DNA Circles and HTLV-1 PVL
2.6. Controls
2.7. LTR DNA Localisation
2.8. Reverse Transcriptase Inhibitory Study
2.9. Cell Entry Inhibitor Study
2.10. Statistical Analysis
3. Results
3.1. Detection, Localisation, Quantification of LTR DNA Circles
3.2. Biomarker for Persistent RT Usage
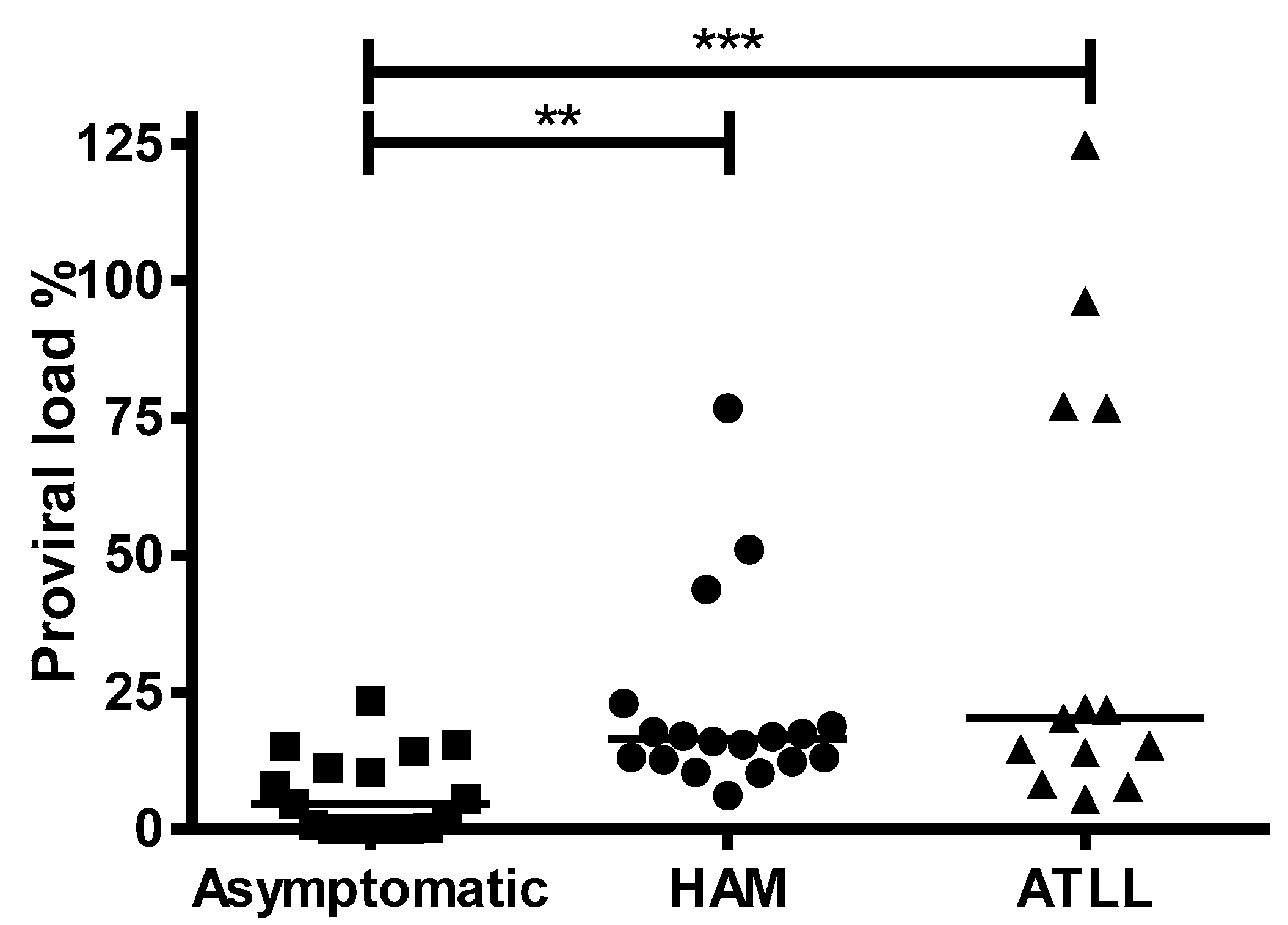
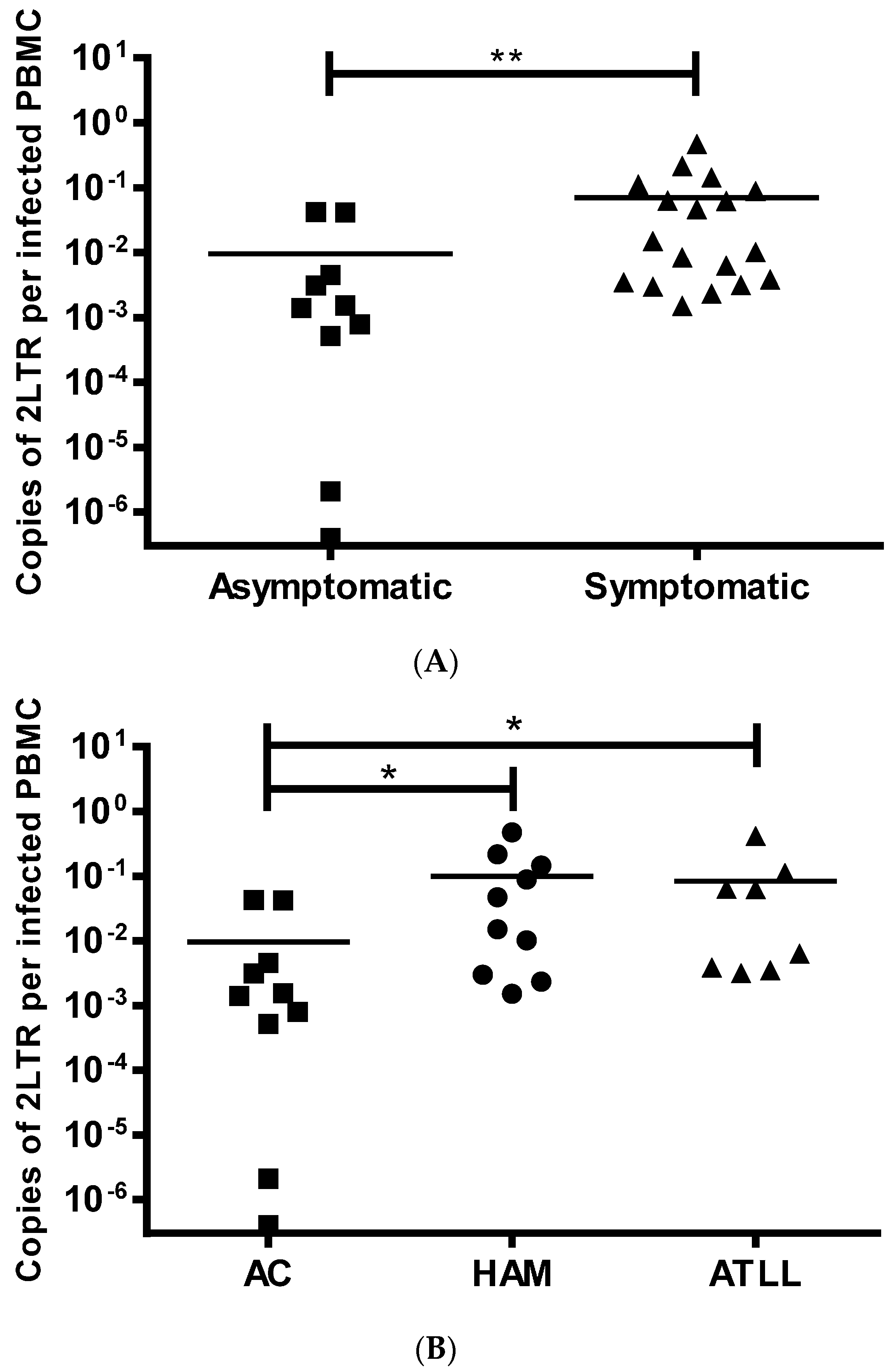
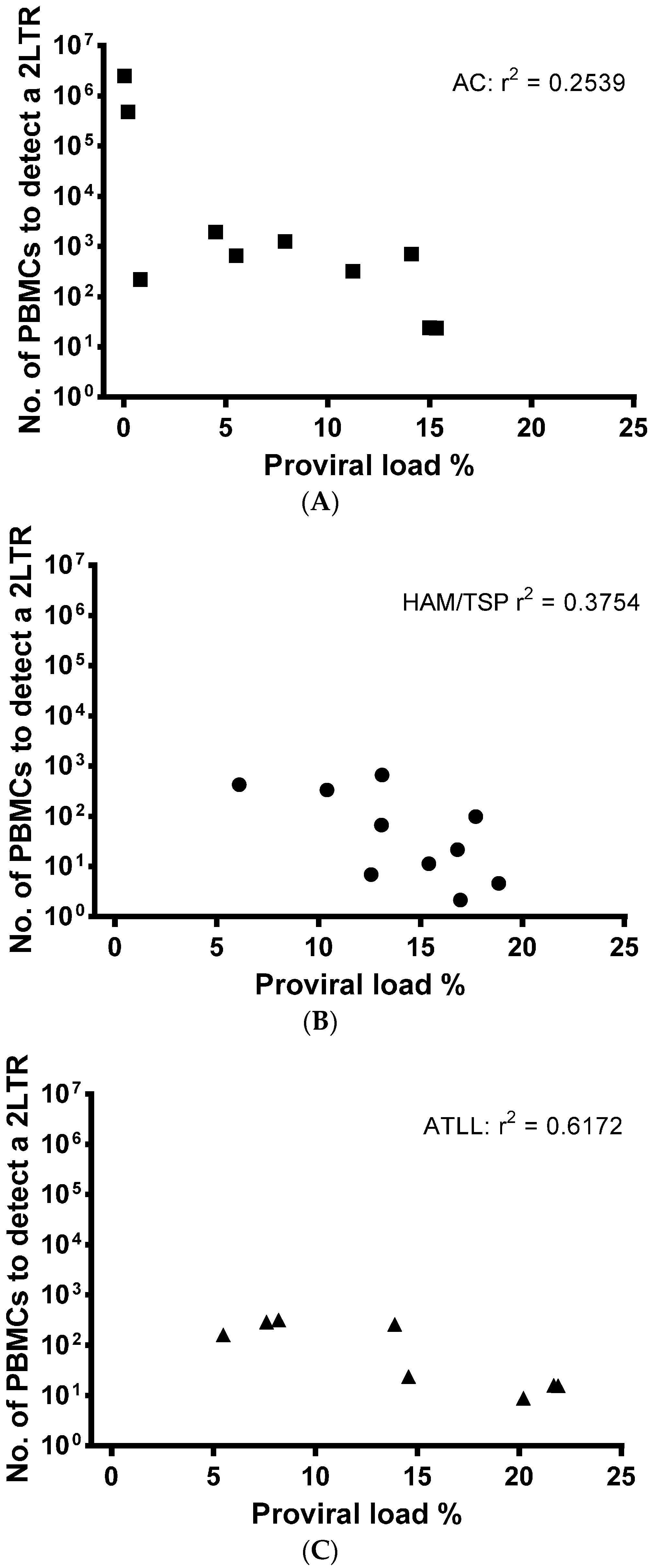
3.3. LTR DNA Circles in HTLV-1 Infected Patients
3.4. Biomarker for HTLV Disease Progression
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflict of Interest
References
- Hinuma, Y.; Nagata, K.; Hanaoka, M.; Nakai, M.; Matsumoto, T.; Kinoshita, K.I.; Shirakawa, S.; Miyoshi, I. Adult T-cell leukemia: antigen in an ATL cell line and detection of antibodies to the antigen in human sera. Proc. Natl. Acad. Sci. USA 1981, 78, 6476–6480. [Google Scholar] [CrossRef] [PubMed]
- Poiesz, B.J.; Ruscetti, F.W.; Gazdar, A.F.; Bunn, P.A.; Minna, J.D.; Gallo, R.C. Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc. Natl. Acad. Sci. USA 1980, 77, 7415–7419. [Google Scholar] [CrossRef] [PubMed]
- Osame, M.; Usuku, K.; Izumo, S.; Ijichi, N.; Amitani, H.; Igata, A.; Matsumoto, M.; Tara, M. HTLV-I associated myelopathy, a new clinical entity. Lancet 1986, 1, 1031–1032. [Google Scholar] [CrossRef]
- Gessain, A.; Barin, F.; Vernant, J.C.; Gout, O.; Maurs, L.; Calender, A.; de The, G. Antibodies to human T-lymphotropic virus type-I in patients with tropical spastic paraparesis. Lancet 1985, 2, 407–410. [Google Scholar] [CrossRef]
- Seiki, M.; Hattori, S.; Hirayama, Y.; Yoshida, M. Human adult T-cell leukemia virus: complete nucleotide sequence of the provirus genome integrated in leukemia cell DNA. Proc. Natl. Acad. Sci. USA 1983, 80, 3618–3622. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.O.; Bowerman, B.; Varmus, H.E.; Bishop, J.M. Retroviral integration: structure of the initial covalent product and its precursor, and a role for the viral IN protein. Proc. Natl. Acad. Sci. USA 1989, 86, 2525–2529. [Google Scholar] [CrossRef] [PubMed]
- Shank, P.R.; Hughes, S.H.; Kung, H.J.; Majors, J.E.; Quintrell, N.; Guntaka, R.V.; Bishop, J.M.; Varmus, H.E. Mapping unintegrated avian sarcoma virus DNA: termini of linear DNA bear 300 nucleotides present once or twice in two species of circular DNA. Cell 1978, 15, 1383–1395. [Google Scholar] [CrossRef]
- Gianni, A.M.; Weinberg, R.A. Partially single-stranded form of free Moloney viral DNA. Nature 1975, 255, 646–648. [Google Scholar] [CrossRef] [PubMed]
- Dina, D.; Benz, E.W., Jr. Structure of murine sarcoma virus DNA replicative intermediates synthesized in vitro. J. Virol. 1980, 33, 377–389. [Google Scholar] [PubMed]
- Gilboa, E.; Goff, S.; Shields, A.; Yoshimura, F.; Mitra, S.; Baltimore, D. In vitro synthesis of a 9 kbp terminally redundant DNA carrying the infectivity of Moloney murine leukemia virus. Cell 1979, 16, 863–874. [Google Scholar] [CrossRef]
- Swanstrom, R.; DeLorbe, W.J.; Bishop, J.M.; Varmus, H.E. Nucleotide sequence of cloned unintegrated avian sarcoma virus DNA: viral DNA contains direct and inverted repeats similar to those in transposable elements. Proc. Natl. Acad. Sci. USA 1981, 78, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Katz, R.A.; Omer, C.A.; Weis, J.H.; Mitsialis, S.A.; Faras, A.J.; Guntaka, R.V. Restriction endonuclease and nucleotide sequence analyses of molecularly cloned unintegrated avian tumor virus DNA: structure of large terminal repeats in circle junctions. J. Virol. 1982, 42, 346–351. [Google Scholar] [PubMed]
- Farnet, C.M.; Haseltine, W.A. Circularization of human immunodeficiency virus type 1 DNA in vitro. J. Virol. 1991, 65, 6942–6952. [Google Scholar] [PubMed]
- Pauza, C.D.; Galindo, J.E.; Richman, D.D. Reinfection results in accumulation of unintegrated viral DNA in cytopathic and persistent human immunodeficiency virus type 1 infection of CEM cells. J. Exp. Med. 1990, 172, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Robinson, H.L.; Zinkus, D.M. Accumulation of human immunodeficiency virus type 1 DNA in T cells: results of multiple infection events. J. Virol. 1990, 64, 4836–4841. [Google Scholar] [PubMed]
- Chun, T.W.; Carruth, L.; Finzi, D.; Shen, X.; DiGiuseppe, J.A.; Taylor, H.; Hermankova, M.; Chadwick, K.; Margolick, J.; Quinn, T.C.; et al. Quantification of latent tissue reservoirs and total body viral load in HIV-1 infection. Nature 1997, 387, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, M.E.; Teo, I.; Greenough, T.; Sharova, N.; Luzuriaga, K.; Sullivan, J.L.; Bucy, R.P.; Kostrikis, L.G.; Haase, A.; Veryard, C.; et al. Persistence of episomal HIV-1 infection intermediates in patients on highly active anti-retroviral therapy. Nat. Med. 2000, 6, 76–81. [Google Scholar] [PubMed]
- Zennou, V.; Petit, C.; Guetard, D.; Nerhbass, U.; Montagnier, L.; Charneau, P. HIV-1 genome nuclear import is mediated by a central DNA flap. Cell 2000, 101, 173–185. [Google Scholar] [CrossRef]
- Buzon, M.J.; Massanella, M.; Llibre, J.M.; Esteve, A.; Dahl, V.; Puertas, M.C.; Gatell, J.M.; Domingo, P.; Paredes, R.; Sharkey, M.; et al. HIV-1 replication and immune dynamics are affected by raltegravir intensification of HAART-suppressed subjects. Nat. Med. 2010, 16, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, A.; Barez, P.Y.; Hamaidia, M.; Gazon, H.; de Brogniez, A.; Perike, S.; Gillet, N.; Willems, L. Modes of Human T Cell Leukemia Virus Type 1 Transmission, Replication and Persistence. Viruses 2015, 7, 3603–3624. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.L.; Maldonado, J.O.; Mueller, J.D.; Zhang, W.; Mansky, L.M. Molecular Studies of HTLV-1 Replication: An Update. Viruses 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.S.; Petrow-Sadowski, C.; Huang, Y.K.; Bertolette, D.C.; Ruscetti, F.W. Cell-free HTLV-1 infects dendritic cells leading to transmission and transformation of CD4(+) T cells. Nat. Med. 2008, 14, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, M.; Jeang, K.T. Human T-cell leukaemia virus type 1 (HTLV-1) infectivity and cellular transformation. Nature reviews. Cancer 2007, 7, 270–280. [Google Scholar] [PubMed]
- Alais, S.; Mahieux, R.; Dutartre, H. Viral Source-Independent High Susceptibility of Dendritic Cells to Human T-Cell Leukemia Virus Type 1 Infection Compared to That of T Lymphocytes. J. Virol. 2015, 89, 10580–10590. [Google Scholar] [CrossRef] [PubMed]
- Demontis, M.A.; Sadiq, M.T.; Golz, S.; Taylor, G.P. HTLV-1 viral RNA is detected rarely in plasma of HTLV-1 infected subjects. J Med Virol 2015, 87, 2130–2134. [Google Scholar] [CrossRef] [PubMed]
- Derse, D.; Mikovits, J.; Waters, D.; Brining, S.; Ruscetti, F. Examining the molecular genetics of HTLV-I with an infectious molecular clone of the virus and permissive cell culture systems. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1996, 12, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.F.; Vandamme, A.M.; Kazadi, K.; Carton, H.; Desmyter, J.; Goubau, P. Familial transmission and minimal sequence variability of human T-lymphotropic virus type I (HTLV-I) in Zaire. AIDS Res. Hum. Retroviruses. 1994, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Cook, L.B.; Melamed, A.; Demontis, M.A.; Laydon, D.J.; Fox, J.M.; Tosswill, J.H.; de Freitas, D.; Price, A.D.; Medcalf, J.F.; Martin, F.; et al. Rapid dissemination of human T-lymphotropic virus type 1 during primary infection in transplant recipients. Retrovirology 2016, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Demontis, M.A.; Hilburn, S.; Taylor, G.P. Human T cell lymphotropic virus type 1 viral load variability and long-term trends in asymptomatic carriers and in patients with human T cell lymphotropic virus type 1-related diseases. AIDS Res. Hum. Retrovir. 2013, 29, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.A.; Lloyd, P.A.; McDonald, S.; Wykoff, J.; Derse, D. Susceptibility of human T cell leukemia virus type I to nucleoside reverse transcriptase inhibitors. J. Infect. Dis. 2003, 188, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Ratner, L. Adult T cell leukemia lymphoma. Front. Biosci. 2004, 9, 2852–2859. [Google Scholar] [CrossRef] [PubMed]
- Macchi, B.; Balestrieri, E.; Ascolani, A.; Hilburn, S.; Martin, F.; Mastino, A.; Taylor, G.P. Susceptibility of primary HTLV-1 isolates from patients with HTLV-1-associated myelopathy to reverse transcriptase inhibitors. Viruses 2011, 3, 469–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevino, A.; Parra, P.; Bar-Magen, T.; Garrido, C.; de Mendoza, C.; Soriano, V. Antiviral effect of raltegravir on HTLV-1 carriers. J. Antimicrob. Chemother. 2012, 67, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Matutes, E.; Taylor, G.P.; Cavenagh, J.; Pagliuca, A.; Bareford, D.; Domingo, A.; Hamblin, M.; Kelsey, S.; Mir, N.; Reilly, J.T. Interferon alpha and zidovudine therapy in adult T-cell leukaemia lymphoma: response and outcome in 15 patients. Br. J. Haematol. 2001, 113, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Taylor, G.P.; Goon, P.; Furukawa, Y.; Green, H.; Barfield, A.; Mosley, A.; Nose, H.; Babiker, A.; Rudge, P.; Usuku, K.; et al. Zidovudine plus lamivudine in Human T-Lymphotropic Virus type-I-associated myelopathy: a randomised trial. Retrovirology 2006, 3, 63. [Google Scholar] [CrossRef] [PubMed]
- Afonso, P.V.; Mekaouche, M.; Mortreux, F.; Toulza, F.; Moriceau, A.; Wattel, E.; Gessain, A.; Bangham, C.R.; Dubreuil, G.; Plumelle, Y.; et al. Highly active antiretroviral treatment against STLV-1 infection combining reverse transcriptase and HDAC inhibitors. Blood 2010, 116, 3802–3808. [Google Scholar] [CrossRef] [PubMed]
- Nejmeddine, M.; Negi, V.S.; Mukherjee, S.; Tanaka, Y.; Orth, K.; Taylor, G.P.; Bangham, C.R. HTLV-1-Tax and ICAM-1 act on T-cell signal pathways to polarize the microtubule-organizing center at the virological synapse. Blood 2009, 114, 1016–1025. [Google Scholar] [CrossRef] [PubMed]
- Igakura, T.; Stinchcombe, J.C.; Goon, P.K.; Taylor, G.P.; Weber, J.N.; Griffiths, G.M.; Tanaka, Y.; Osame, M.; Bangham, C.R. Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science 2003, 299, 1713–1716. [Google Scholar] [CrossRef] [PubMed]
- Nejmeddine, M.; Barnard, A.L.; Tanaka, Y.; Taylor, G.P.; Bangham, C.R. Human T-lymphotropic virus, type 1, tax protein triggers microtubule reorientation in the virological synapse. J. Biol. Chem. 2005, 280, 29653–29660. [Google Scholar] [CrossRef] [PubMed]
- Majorovits, E.; Nejmeddine, M.; Tanaka, Y.; Taylor, G.P.; Fuller, S.D.; Bangham, C.R. Human T-lymphotropic virus-1 visualized at the virological synapse by electron tomography. PLoS ONE 2008, 3, e2251. [Google Scholar] [CrossRef] [PubMed]
- Pais-Correia, A.M.; Sachse, M.; Guadagnini, S.; Robbiati, V.; Lasserre, R.; Gessain, A.; Gout, O.; Alcover, A.; Thoulouze, M.I. Biofilm-like extracellular viral assemblies mediate HTLV-1 cell-to-cell transmission at virological synapses. Nat. Med. 2010, 16, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Overbaugh, J.; Bangham, C.R. Selection forces and constraints on retroviral sequence variation. Science 2001, 292, 1106–1109. [Google Scholar] [CrossRef] [PubMed]
- Tosswill, J.H.; Taylor, G.P.; Clewley, J.P.; Weber, J.N. Quantification of proviral DNA load in human T-cell leukaemia virus type I infections. J. Virol. Methods 1998, 75, 21–26. [Google Scholar] [CrossRef]
- Koressaar, T.R.M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.; Remm, M.; Rozen, S. Primer3 - new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Goon, P.K.; Hanon, E.; Igakura, T.; Tanaka, Y.; Weber, J.N.; Taylor, G.P.; Bangham, C.R. High frequencies of Th1-type CD4(+) T cells specific to HTLV-1 Env and Tax proteins in patients with HTLV-1-associated myelopathy/tropical spastic paraparesis. Blood 2002, 99, 3335–3341. [Google Scholar] [CrossRef] [PubMed]
- Pinon, J.D.; Kelly, S.M.; Price, N.C.; Flanagan, J.U.; Brighty, D.W. An antiviral peptide targets a coiled-coil domain of the human T-cell leukemia virus envelope glycoprotein. J. Virol. 2003, 77, 3281–3290. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.M.; Hilburn, S.; Cook, L.B.; Demontis, M.A.; Rowan, A.G.; Martin, F.; Taylor, G.P. Circularised 1 and 2 LTR DNA circles are present in freshly- and chronically-infected cell lines and patient PBMCs, indicating ongoing reverse transcriptase usage. Retrovirology 2011, 8 (Suppl 1), A51. [Google Scholar] [CrossRef]
- Salahuddin, S.Z.; Markham, P.D.; Wong-Staal, F.; Franchini, G.; Kalyanaraman, V.S.; Gallo, R.C. Restricted expression of human T-cell leukemia--lymphoma virus (HTLV) in transformed human umbilical cord blood lymphocytes. Virology 1983, 129, 51–64. [Google Scholar] [CrossRef]
- Harada, S.; Koyanagi, Y.; Yamamoto, N. Infection of HTLV-III/LAV in HTLV-I-carrying cells MT-2 and MT-4 and application in a plaque assay. Science 1985, 229, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.; Fedina, A.; Youshya, S.; Taylor, G.P. A 15-year prospective longitudinal study of disease progression in patients with HTLV-1 associated myelopathy in the UK. J. Neurol. Neurosurg. Psychiatry 2010, 81, 1336–1340. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, K.; Yonemoto, R.; Yoshikura, H. Epigenetic control and reintegration of extrachromosomal proviral DNA in HL60 cells chronically infected with human T cell leukaemia virus type 1. J. Gen. Virol. 1988, 69 (Pt 3), 651–658. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Besansky, N.J.; Rudolph, D.; Nutman, T.B.; Folks, T.M.; Lal, R.B. Unintegrated two-long terminal repeat circular human T lymphotropic virus DNA accumulation during chronic HTLV infection. AIDS Res. Hum. Retrovir. 1993, 9, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Yoder, K.E.; Fishel, R. PCR-based detection is unable to consistently distinguish HIV 1LTR circles. J. Virol. Methods 2006, 138, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Macatonia, S.E.; Cruickshank, J.K.; Rudge, P.; Knight, S.C. Dendritic cells from patients with tropical spastic paraparesis are infected with HTLV-1 and stimulate autologous lymphocyte proliferation. AIDS Res. Hum. Retrovir. 1992, 8, 1699–1706. [Google Scholar] [CrossRef] [PubMed]
- Hishizawa, M.; Imada, K.; Kitawaki, T.; Ueda, M.; Kadowaki, N.; Uchiyama, T. Depletion and impaired interferon-alpha-producing capacity of blood plasmacytoid dendritic cells in human T-cell leukaemia virus type I-infected individuals. Br. J. Haematol. 2004, 125, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Pauza, C.D.; Galindo, J. Persistent human immunodeficiency virus type 1 infection of monoblastoid cells leads to accumulation of self-integrated viral DNA and to production of defective virions. J. Virol. 1989, 63, 3700–3707. [Google Scholar] [PubMed]
- Kattan, T.; MacNamara, A.; Rowan, A.G.; Nose, H.; Mosley, A.J.; Tanaka, Y.; Taylor, G.P.; Asquith, B.; Bangham, C.R. The avidity and lytic efficiency of the CTL response to HTLV-1. J. Immunol. 2009, 182, 5723–5729. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Marsh, J.W. Early transcription from nonintegrated DNA in human immunodeficiency virus infection. J. Virol. 2003, 77, 10376–10382. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Marsh, J.W. Gene transcription in HIV infection. Microbes Infect. 2003, 5, 1023–1027. [Google Scholar] [CrossRef]
- Wu, Y.; Marsh, J.W. Selective transcription and modulation of resting T cell activity by preintegrated HIV DNA. Science 2001, 293, 1503–1506. [Google Scholar] [CrossRef] [PubMed]
- Poon, B.; Chen, I.S. Human immunodeficiency virus type 1 (HIV-1) Vpr enhances expression from unintegrated HIV-1 DNA. J. Virol. 2003, 77, 3962–3972. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.Y.; Mikovits, J.A.; Bagni, R.; Petrow-Sadowski, C.L.; Ruscetti, F.W. Infection of lymphoid cells by integration-defective human immunodeficiency virus type 1 increases de novo methylation. J. Virol. 2001, 75, 9753–9761. [Google Scholar] [CrossRef] [PubMed]
- Cara, A.; Cereseto, A.; Lori, F.; Reitz, M.S., Jr. HIV-1 protein expression from synthetic circles of DNA mimicking the extrachromosomal forms of viral DNA. J. Biol. Chem. 1996, 271, 5393–5397. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primer/Probe | Sequence | Product Size |
|---|---|---|
| 1LTR probe | CTCTCACACGGCCTCATACAGTACTCTTCCTTTCATAGTTTACATCTCCTGTTTGAATTATTCCCTAGGCAATGGGCCAAATCTTTTCCCGTAGCGCTAG | 100 bp |
| 1LTR FWD | CTCTCACACGGCCTCATACA | 940 bp |
| 1LTR REV | CTAGCGCTACGGGAAAAGAT | |
| 1LTR nested probe | CACCAACATCCCCATTTCTCGTACTCTTCCTTTCATAGTTTACATCTCCTGTTTGAATTATTCCCTAGGCAATGGGCCAAACTGGCAATGGGCCAAATCT | 100 bp |
| 1LTR nested FWD | CACCAACATCCCCATTTCTC | 840 bp |
| 1LTR nested REV | AGATTTGGCCCATTGC | |
| 2LTR probe | ATGAGCCCCAAATATCCCCCGGGGGTACTCTTCCTTTCATAGTTTACATCTCCTGTTAGTTGAATTATTCCCTAGGCAGTTCTGCGCCGTTACAGATCGA | 100 bp |
| 2LTR FWD | ATGAGCCCCAAATATCCCCCGGGG | 611 bp |
| 2LTR REV | TCGATCTGTAACGGCGCAGAAC | |
| 2LTR nested probe | AGCCACCGGGAACCACCCATGTACTCTTCCTTTCATAGTTTACATTTGTTTGAATTATTCCCTAGGCAATGGGCCAGGTCGAGACCGGGCCTTTGTC | 100 bp |
| 2LTR nested FWD | AGCCACCGGGAACCACCCAT | 307 bp |
| 2LTR nested REV | GACAAAGGCCCGGTCTCGACCT |
| Cell Line | Limit of Detection in 1 μg DNA | Ratio of 1:2 LTR | |
|---|---|---|---|
| 1LTR | 2LTR | ||
| MT-2 | 8.79 copies | 14.67 copies | 1:1.7 |
| HuT 102 | 5.51 copies | 11.03 copies | 1:2 |
| Patient | VL% (Range) | 1LTR | 1LTR% |
|---|---|---|---|
| HBK | 129 | + | 0 |
| HBU | 3.94 | + | 0 |
| HCT | 0.08 | + | 0.0023 |
| HEF | 2.39 | + | 0 |
| HT | 23.4 | - | 0 |
| HX | 0.02 | - | 0 |
| ACs mean | 26.5 (0.02–129) | ||
| TAF | 76.8 | + | 0.0048 |
| TAN | 16 | + | 0.0048 |
| TAQ | 12.3 | + | 0.0023 |
| TAS | 17.4 | + | 0.00048 |
| TBW | 12.8 | + | 0.0023 |
| TBX | 13.28 | + | 0.000048 |
| TBR | 22.9 | + | 0 |
| TBJ | 51 | - | 0 |
| TBU | 10.2 | - | 0 |
| TCD | 43.7 | - | 0 |
| HAM mean | 27.6 (10–77) | ||
| LER | 76.75 | + | 0.00023 |
| LEV* | 124.7 | + | 0.11547 |
| LEY | 77.1 | + | 0.0048 |
| LP4 | 96.3 | - | 0 |
| ATLL mean | 94 (77–125) |
| Patient | VL% (Range) | 2LTR | 2LTR% |
|---|---|---|---|
| HBU | 4.5 | + | 0.0005184 |
| HBZ | 5.5 | + | 0.0015246 |
| HBF | 7.9 | + | 0.00079 |
| HBE | 14.1 | + | 0.00141 |
| HAO | 0.21 | + | 0.0000021 |
| HBK | 10.3 | Nested + | |
| HBX | 11.23 | + | 0.003112956 |
| HCH | 15 | + | 0.04158 |
| HDH | 0.04 | + | 0.0000004 |
| HDO | 0.01 | - | Undetectable |
| HDY | 0.06 | Nested + | |
| HFE | 15.33 | + | 0.04249476 |
| HT-UV1 | 0.82 | + | 0.00454608 |
| ACs mean | 6.54 (0.01–15) | ||
| TBP | 6.1 | + | 0.0023424 |
| TBW | 13.1 | + | 0.00150912 |
| TBG | 16.8 | + | 0.0465696 |
| TBA | 17.7 | + | 0.0101952 |
| TAL | 15.4 | + | 0.088704 |
| TAT | 10.4 | + | 0.0029952 |
| TBZ | 18.83 | + | 0.2169216 |
| TCJ | 16.95 | + | 0.469854 |
| TCP | 13.07 | + | 0.01505664 |
| TCQ | 12.57 | + | 0.1448064 |
| HAM mean | 14.09 (6–19) | ||
| LEU | 7.6 | + | 0.00347776 |
| LGH | 13.88 | + | 0.003847536 |
| LGA | 21.68 | + | 0.0624384 |
| LFE | 15.2 | + | 0.210672 |
| TBX | 14.57 | + | 0.00839232 |
| LFK * | 8.2 | + | 0.0031488 |
| LFA * | 21.9 | + | 0.063072 |
| LFP * | 5.48 | + | 0.00631296 |
| ATLL mean | 13.56 (5–22) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fox, J.M.; Hilburn, S.; Demontis, M.-A.; Brighty, D.W.; Rios Grassi, M.F.; Galvão-Castro, B.; Taylor, G.P.; Martin, F. Long Terminal Repeat Circular DNA as Markers of Active Viral Replication of Human T Lymphotropic Virus-1 in Vivo. Viruses 2016, 8, 80. https://0-doi-org.brum.beds.ac.uk/10.3390/v8030080
Fox JM, Hilburn S, Demontis M-A, Brighty DW, Rios Grassi MF, Galvão-Castro B, Taylor GP, Martin F. Long Terminal Repeat Circular DNA as Markers of Active Viral Replication of Human T Lymphotropic Virus-1 in Vivo. Viruses. 2016; 8(3):80. https://0-doi-org.brum.beds.ac.uk/10.3390/v8030080
Chicago/Turabian StyleFox, James M, Silva Hilburn, Maria-Antonietta Demontis, David W Brighty, Maria Fernanda Rios Grassi, Bernardo Galvão-Castro, Graham P Taylor, and Fabiola Martin. 2016. "Long Terminal Repeat Circular DNA as Markers of Active Viral Replication of Human T Lymphotropic Virus-1 in Vivo" Viruses 8, no. 3: 80. https://0-doi-org.brum.beds.ac.uk/10.3390/v8030080