
Pleiotropic Effects of Atorvastatin Result in a Downregulation of the Carboxypeptidase U System (CPU, TAFIa, CPB2) in a Mouse Model of Advanced Atherosclerosis

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Study Protocol
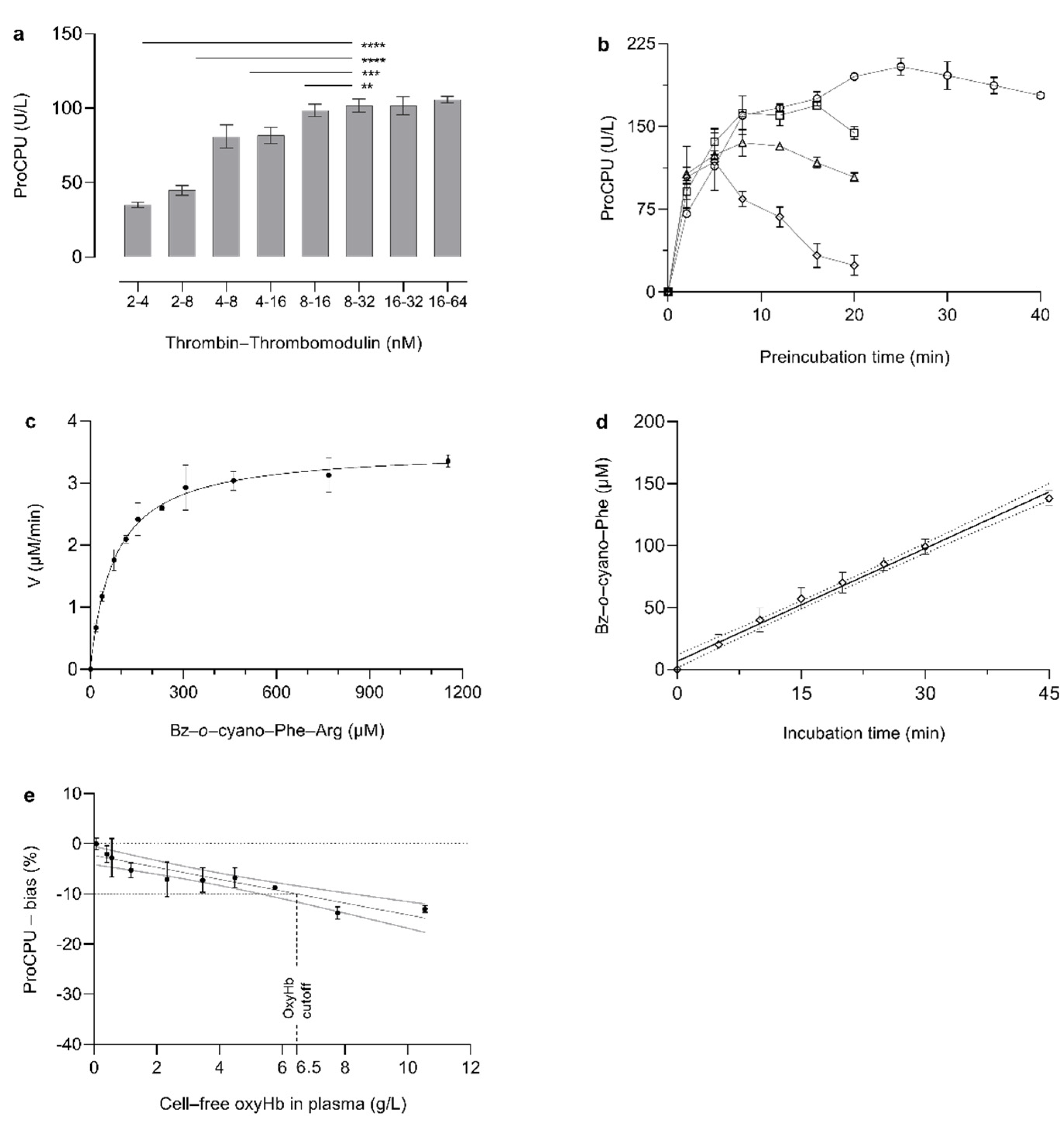
2.2. Activity Assay for the Measurement of ProCPU in Plasma of Mice
2.3. Measurement of Total Plasma Cholesterol, CRP and Blood Immune Cells
2.4. Statistical Analysis
3. Results and Discussion
3.1. Activity Assay for the Measurement of ProCPU in Mice
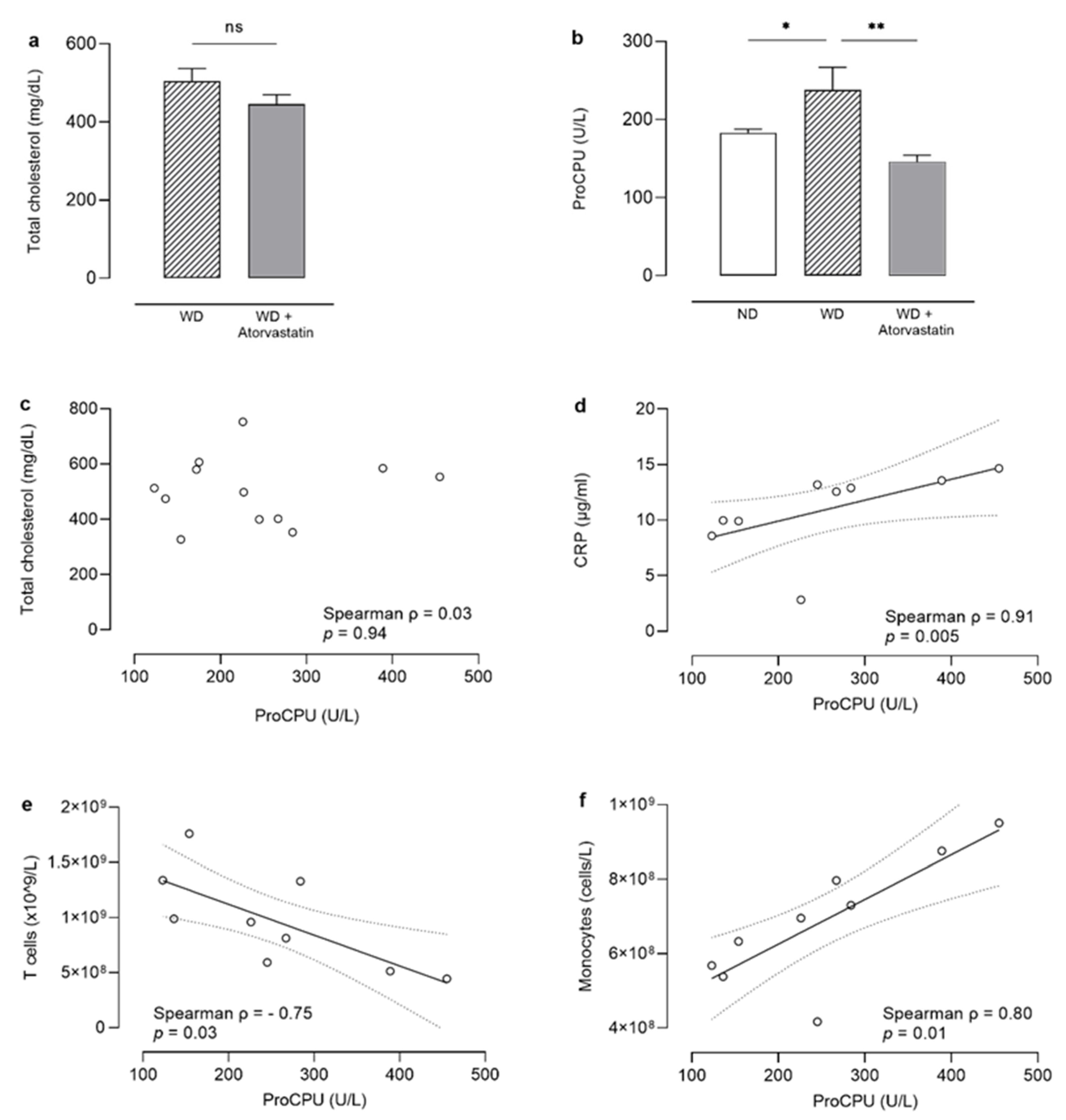
3.2. Total Plasma Cholesterol
3.3. ProCPU Decrease in Atorvastatin-Treated Mice on a Western-Diet Is Cholesterol-Independent
3.4. Inflammation and Blood Immune Cells
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hendriks, D.; Wang, W.; Scharpé, S.; Lommaert, M.P.; van Sande, M. Purification and characterization of a new arginine carboxypeptidase in human serum. BBA Gen. Subj. 1990, 1034, 86–92. [Google Scholar] [CrossRef]
- Heylen, E. An update on the role of carboxypeptidase U (TAFIa) in fibrinolysis. Front. Biosci. 2011, 16, 2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leurs, J.; Hendriks, D. Carboxypeptidase U (TAFIa): A metallocarboxypeptidase with a distinct role in haemostasis and a possible risk factor for thrombotic disease. Thromb. Haemost. 2005, 94, 471–487. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.D.; Lazoura, E.; Okada, N.; Okada, H. Inactivation of C3a and C5a octapeptides by carboxypeptidase R and carboxypeptidase N. Microbiol. Immunol. 2002, 46, 131–134. [Google Scholar] [CrossRef]
- Oesterle, A.; Laufs, U.; Liao, J.K. Pleiotropic Effects of Statins on the Cardiovascular System. Circ. Res. 2017, 120, 229–243. [Google Scholar] [CrossRef] [Green Version]
- Cappelletti, R.M. Statins Therapy: Effects on Plasma Fibrinogen Levels and Fibrinolysis. J. Nutr. Disord. Ther. 2013, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bruni, F.; Pasqui, A.L.; Pastorelli, M.; Bova, G.; Di Renzo, M.; Cercigani, M.; Leo, A.; Auteri, A.; Puccetti, L. Effect of atorvastatin on different fibrinolyis mechanisms in hypercholesterolemic subjects. Int. J. Cardiol. 2004, 95, 269–274. [Google Scholar] [CrossRef]
- Claesen, K.; Mertens, J.C.; Basir, S.; De Belder, S.; Maes, J.; Bosmans, J.; Stoffelen, H.; De Meester, I.; Hendriks, D. Effect of Statin Therapy on the Carboxypeptidase U (CPU, TAFIa, CPB2) System in Patients With Hyperlipidemia: A Proof-of-Concept Observational Study. Clin. Ther. 2021, 43, 908–916. [Google Scholar] [CrossRef]
- Emini Veseli, B.; Perrotta, P.; De Meyer, G.R.A.; Roth, L.; Van der Donckt, C.; Martinet, W.; De Meyer, G.R.Y. Animal models of atherosclerosis. Eur. J. Pharmacol. 2017, 816, 3–13. [Google Scholar] [CrossRef]
- Roth, L.; Rombouts, M.; Schrijvers, D.M.; Martinet, W.; De Meyer, G.R.Y. Cholesterol-independent effects of atorvastatin prevent cardiovascular morbidity and mortality in a mouse model of atherosclerotic plaque rupture. Vascul. Pharmacol. 2016, 80, 50–58. [Google Scholar] [CrossRef]
- Heylen, E.; Van Goethem, S.; Willemse, J.; Olsson, T.; Augustyns, K.; Hendriks, D. Development of a sensitive and selective assay for the determination of procarboxypeptidase U (thrombin-activatable fibrinolysis inhibitor) in plasma. Anal. Biochem. 2010, 396, 152–154. [Google Scholar] [CrossRef]
- Mertens, J.C.; Claesen, K.; Leenaerts, D.; Sim, Y.; Lambeir, A.M.; Hendriks, D. Inhibition of the procarboxypeptidase U (proCPU, TAFI, proCPB2) system due to hemolysis. J. Thromb. Haemost. 2019, 17, 878–884. [Google Scholar] [CrossRef]
- Kahn, S.E.; Watkins, B.F.; Bermes, E.W. An evaluation of a spectrophotometric scanning technique for measurement of plasma hemoglobin. Ann. Clin. Lab. Sci. 1981, 11, 126–131. [Google Scholar]
- Bougarne, N.; Weyers, B.; Desmet, S.J.; Deckers, J.; Ray, D.W.; Staels, B.; De Bosscher, K. Molecular actions of PPARα in lipid metabolism and inflammation. Endocr. Rev. 2018, 39, 760–802. [Google Scholar] [CrossRef] [Green Version]
- Yoon, M. The role of PPARα in lipid metabolism and obesity: Focusing on the effects of estrogen on PPARα actions. Pharmacol. Res. 2009, 60, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Kilicarslan, A.; Yavuz, B.; Guven, G.S.; Atalar, E.; Sahiner, L.; Beyazit, Y.; Kekilli, M.; Ozer, N.; Oz, G.; Haznedaroglu, I.C.; et al. Fenofibrate improves endothelial function and decreases thrombin-activatable fibrinolysis inhibitor concentration in metabolic syndrome. Blood Coagul. Fibrinolysis 2008, 19, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Saotome, D.; Takada, K.; Sugimoto, K.; Sasaki, T.; Ishii, H. Peroxisome proliferator-activated receptor-alpha agonists repress expression of thrombin-activatable fibrinolysis inhibitor by decreasing transcript stability. Thromb. Haemost. 2012, 108, 74–85. [Google Scholar] [CrossRef]
- Paumelle, R.; Staels, B. Cross-talk Between Statins and PPARα in Cardiovascular Diseases: Clinical Evidence and Basic Mechanisms. Trends Cardiovasc. Med. 2008, 18, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Paumelle, R.; Staels, B. Peroxisome proliferator-activated receptors mediate pleiotropic actions of statins. Circ. Res. 2007, 100, 1394–1395. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, K.; Myśliwiec, M.; Pawlak, D. Kynurenine pathway—A new link between endothelial dysfunction and carotid atherosclerosis in chronic kidney disease patients. Adv. Med. Sci. 2010, 55, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.H.H.; Garand, M.; Zagorac, B.; Schadinger, S.L.; Scipione, C.; Koschinsky, M.L.; Boffa, M.B. Identification of human thrombin-activatable fibrinolysis inhibitor in vascular and inflammatory cells. Thromb. Haemost. 2011, 105, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Mosnier, L.O.; Meijers, J.C.M.; Bouma, B.N. Regulation of fibrinolysis in plasma by TAFI and protein C is dependent on the concentration of thrombomodulin. Thromb. Haemost. 2001, 85, 5–11. [Google Scholar] [PubMed]
- Jönsson Rylander, A.C.; Lindgren, A.; Deinum, J.; Bergström, G.M.L.; Böttcher, G.; Kalies, I.; Wåhlander, K. Fibrinolysis inhibitors in plaque stability: A morphological association of PAI-1 and TAFI in advanced carotid plaque. J. Thromb. Haemost. 2017, 15, 758–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claesen, K.; Mertens, J.C.; Leenaerts, D.; Hendriks, D. Carboxypeptidase U (CPU, TAFIa, CPB2) in thromboembolic disease: What do we know three decades after its discovery? Int. J. Mol. Sci. 2021, 22, 883. [Google Scholar] [CrossRef] [PubMed]
- Komnenov, D.; Scipione, C.A.; Bazzi, Z.A.; Garabon, J.J.W.; Koschinsky, M.L.; Boffa, M.B. Pro-inflammatory cytokines reduce human TAFI expression via tristetraprolin-mediated mRNA destabilisation and decreased binding of HuR. Thromb. Haemost. 2015, 114, 337–349. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameter | WD | WD + Atorvastatin | p-Value |
|---|---|---|---|
| CRP (µg/mL) | 10.9 (3.6) | 7.4 (2.6) | 0.001 |
| Leukocytes (109/L) | 6.02 (2.42) | 6.77 (2.10) | 0.35 |
| Monocytes (109/L) | 0.62 (0.18) | 0.37 (0.26) | 0.01 |
| Neutrophils (109/L) | 1.54 (1.11) | 0.78 (0.55) | 0.08 |
| Dendritic cells (109/L) | 0.062 (0.057) | 0.087 (0.035) | 0.16 |
| T cells (109/L) | 0.88 (0.24) | 1.49 (0.33) | 0.01 |
| NK cells (109/L) | 0.51 (0.22) | 0.49 (0.17) | 0.93 |
| NKT cells (109/L) | 0.063 (0.041) | 0.066 (0.028) | 0.49 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Claesen, K.; Roth, L.; Mertens, J.C.; Hermans, K.; Sim, Y.; Hendriks, D. Pleiotropic Effects of Atorvastatin Result in a Downregulation of the Carboxypeptidase U System (CPU, TAFIa, CPB2) in a Mouse Model of Advanced Atherosclerosis. Pharmaceutics 2021, 13, 1731. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13101731
Claesen K, Roth L, Mertens JC, Hermans K, Sim Y, Hendriks D. Pleiotropic Effects of Atorvastatin Result in a Downregulation of the Carboxypeptidase U System (CPU, TAFIa, CPB2) in a Mouse Model of Advanced Atherosclerosis. Pharmaceutics. 2021; 13(10):1731. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13101731
Chicago/Turabian StyleClaesen, Karen, Lynn Roth, Joachim C. Mertens, Karlijn Hermans, Yani Sim, and Dirk Hendriks. 2021. "Pleiotropic Effects of Atorvastatin Result in a Downregulation of the Carboxypeptidase U System (CPU, TAFIa, CPB2) in a Mouse Model of Advanced Atherosclerosis" Pharmaceutics 13, no. 10: 1731. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13101731