
Highly Potent GalNAc-Conjugated Tiny LNA Anti-miRNA-122 Antisense Oligonucleotides

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design and Synthesis of ASOs
2.2. Methods for 3′ Labeling of Oligonucleotides with Alexa Fluor 647 Dye
2.3. Animal Study
2.4. Serum Chemistry
2.5. RNA Isolation from Mice Liver
2.6. Quantitative Real-Time PCR
2.7. Evaluation of miR-122 Inhibition Using SplintR® Ligase
2.8. Evaluation of Inhibition via Mobility Shift Assay
2.9. In Vivo/Ex Vivo Studies
2.10. Statistical Analysis
3. Results and Discussions
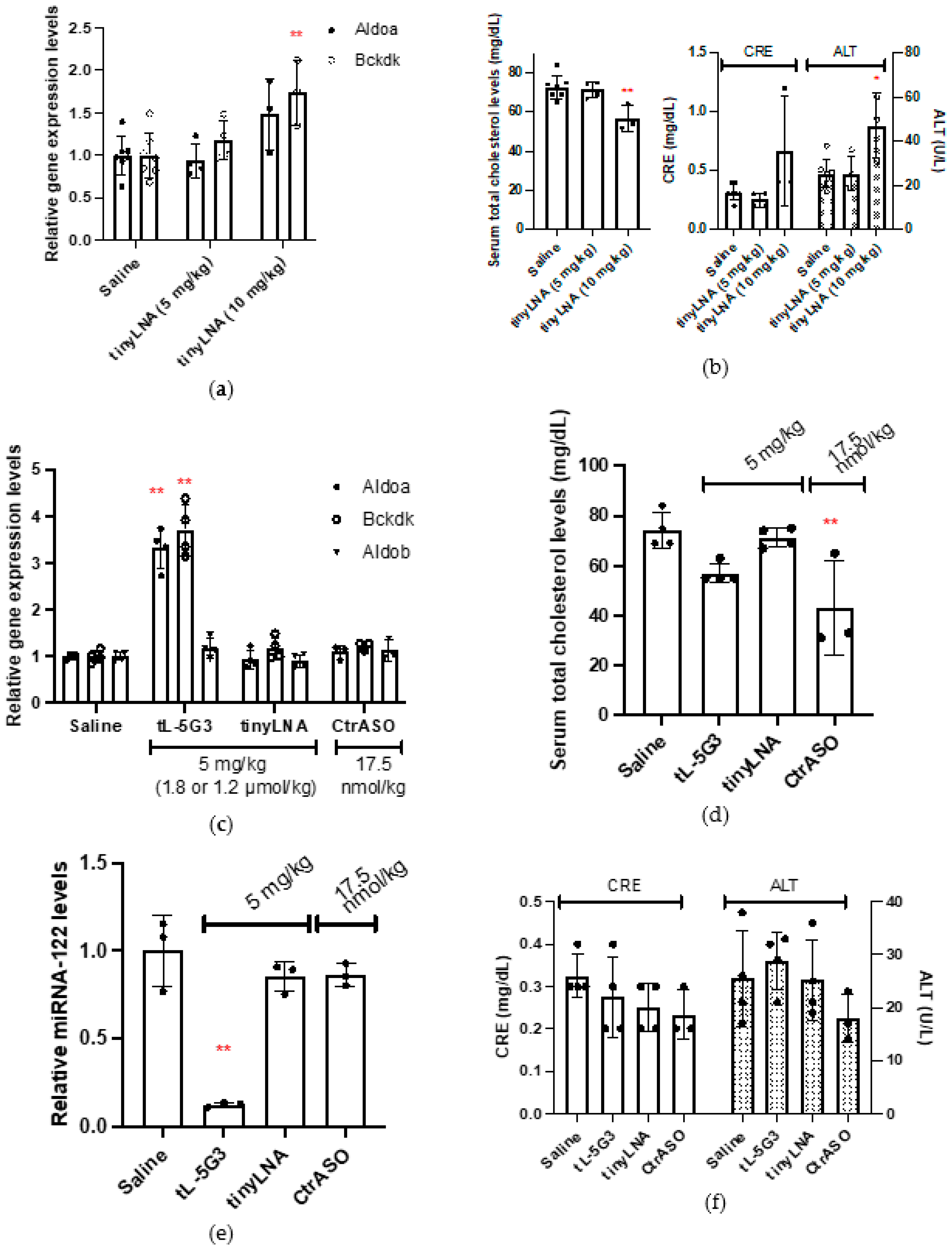
3.1. Effect of GalNAc Conjugation on In Vivo Activity of Tiny LNA
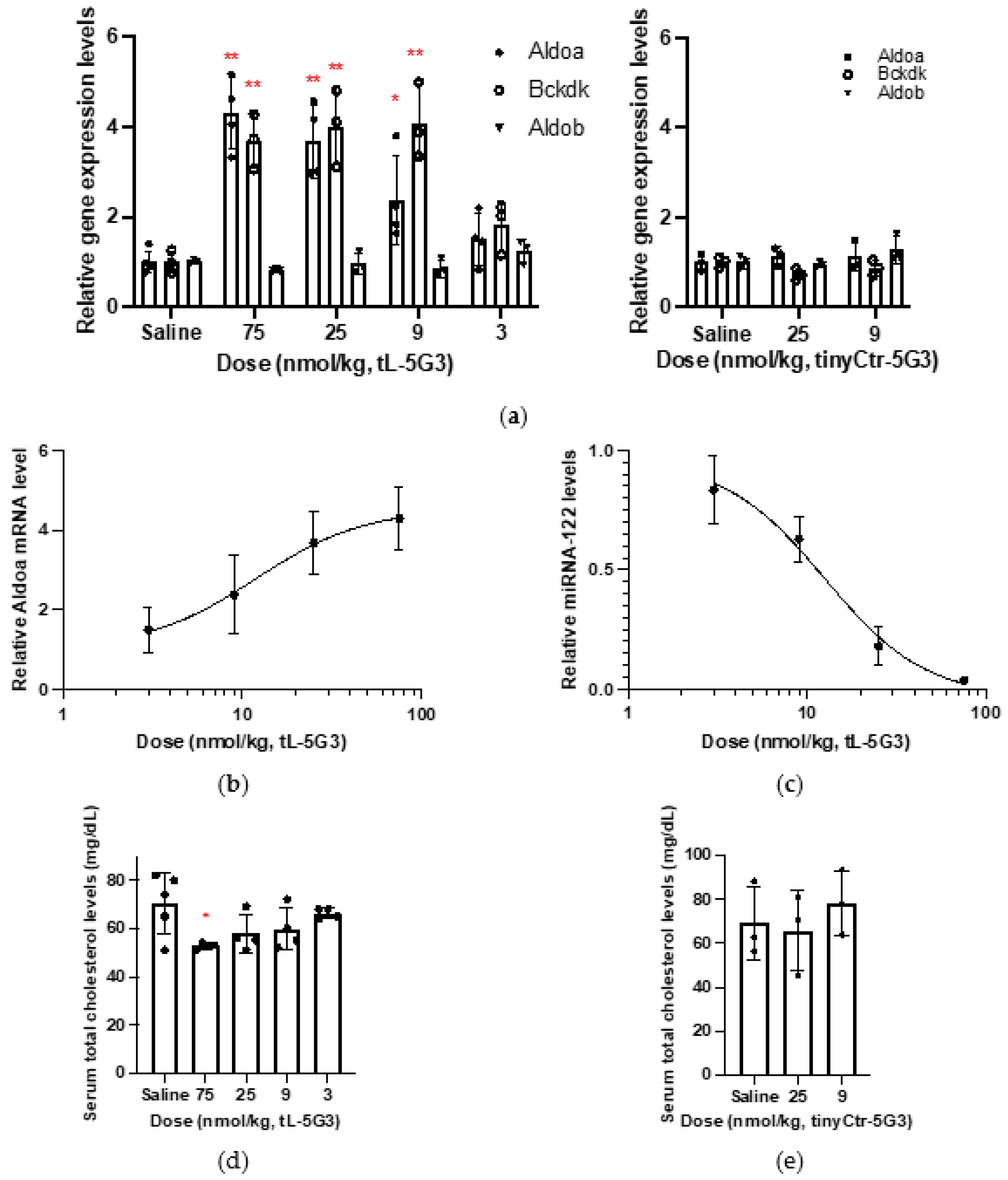
3.2. Evaluation of Dose Responsiveness of In Vivo Activity of GalNAc-Conjugated Tiny LNAs
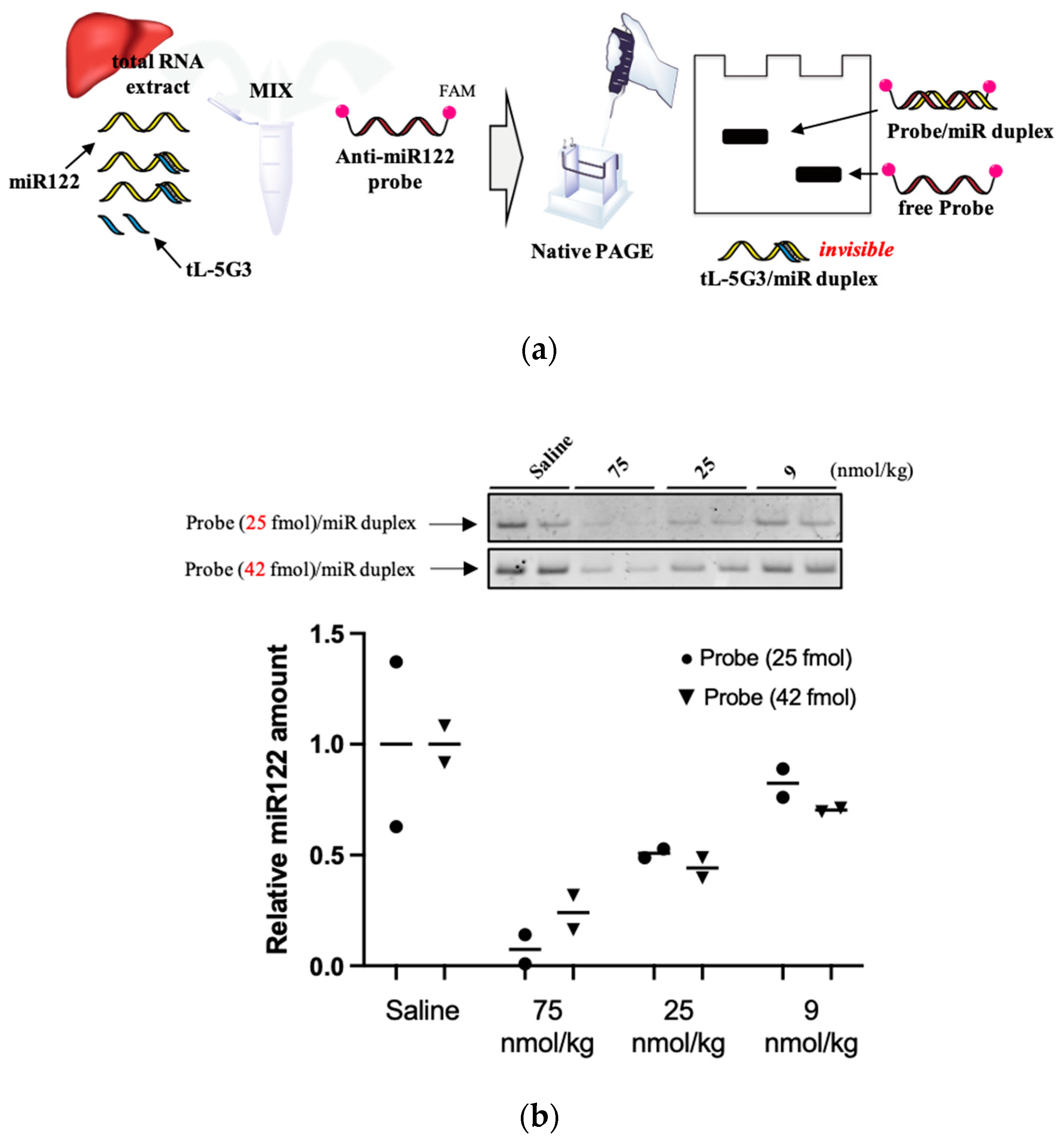
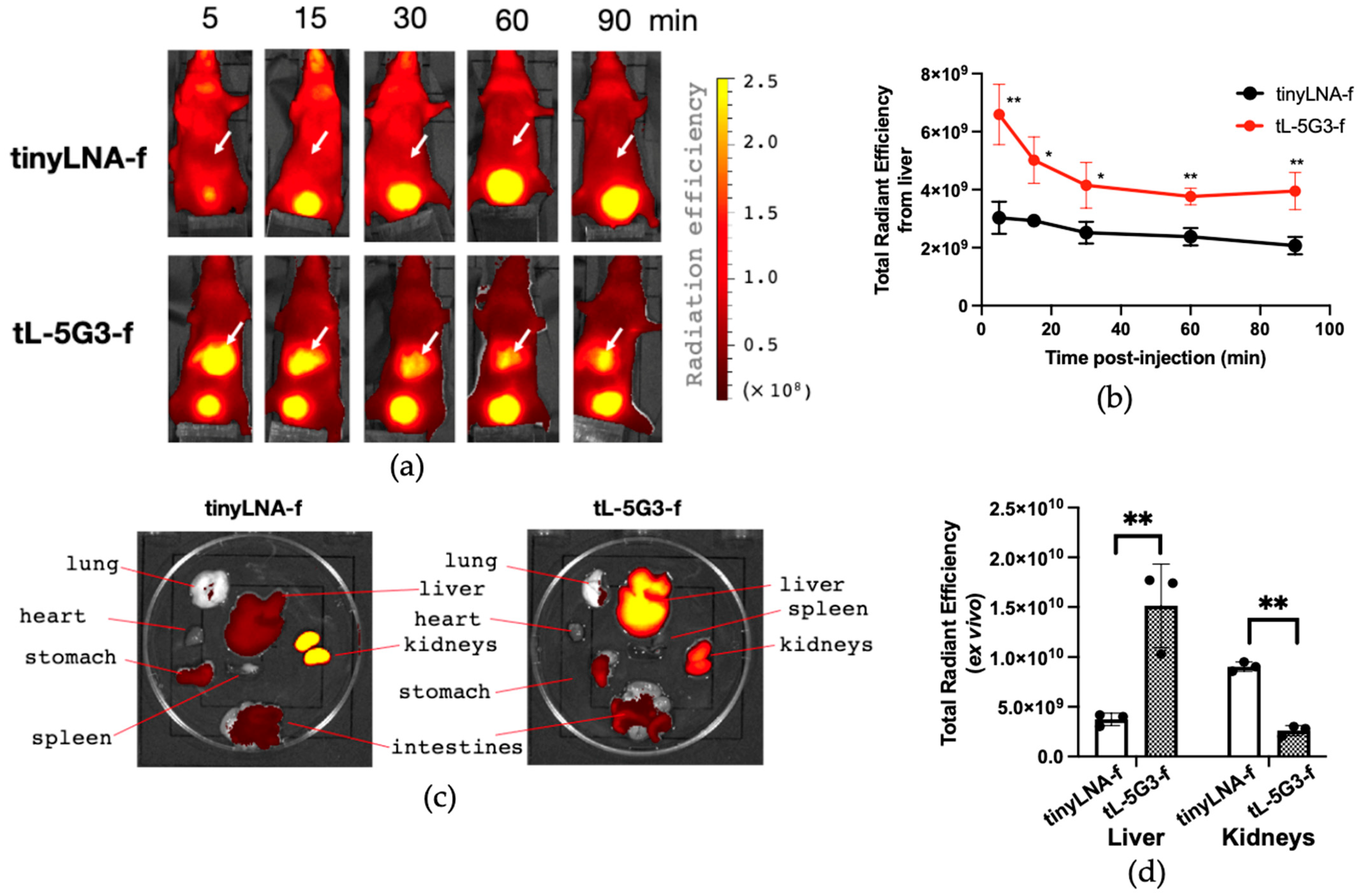
3.3. Visualization of miRNA-122 in Liver
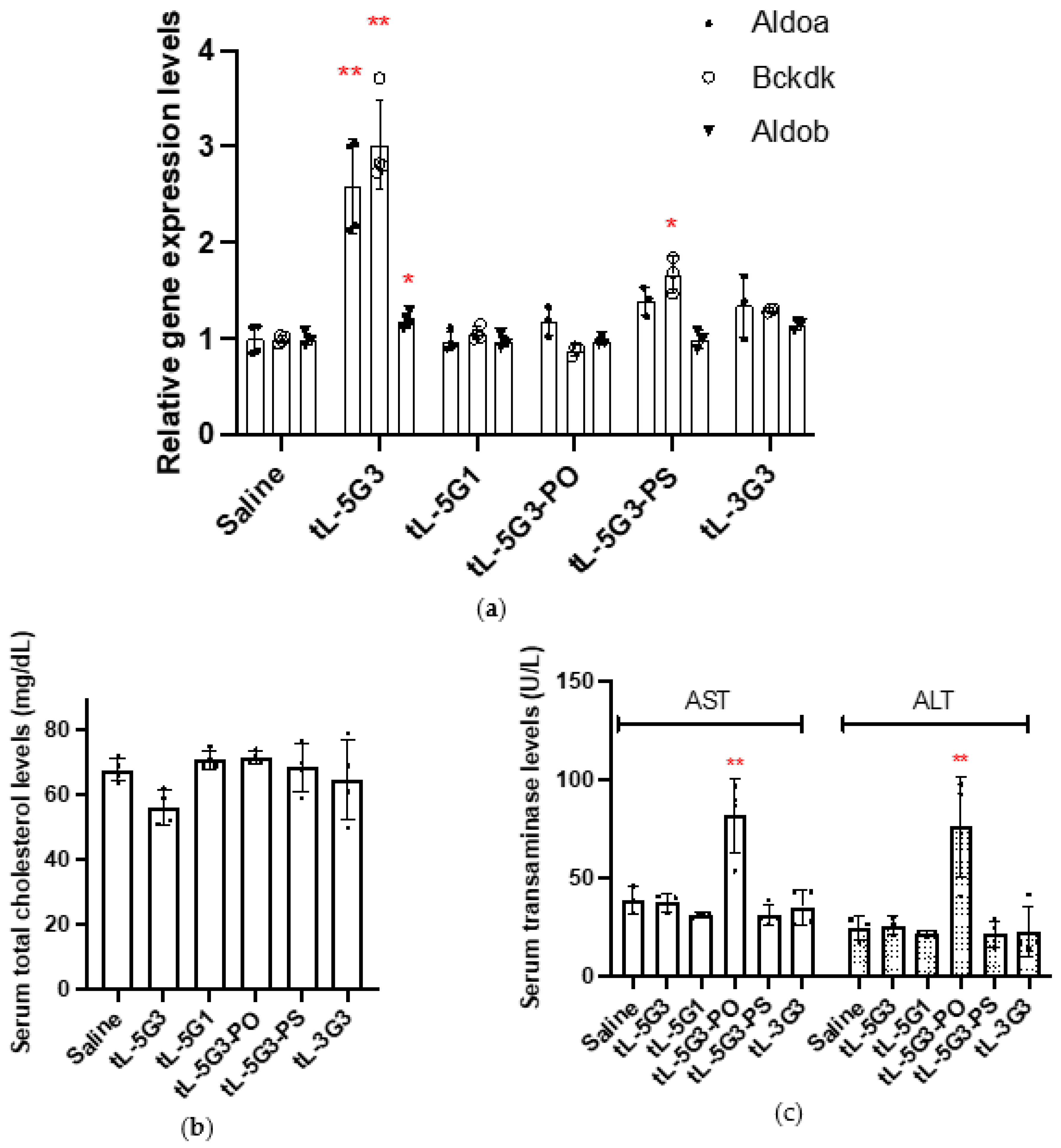
3.4. Configuration–Activity Study of GalNAc-Conjugated Tiny LNA
3.5. Biodistribution Study of GalNAc-Conjugated Tiny LNA
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Baumann, V.; Winkler, J. miRNA-based therapies: Strategies and delivery platforms for oligonucleotide and non-oligonucleotide agents. Future Med. Chem. 2014, 6, 1967–1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, A.M.; Heneghan, H.M.; Miller, N.; Kerin, M.J. The Therapeutic Potential of MicroRNAs: Disease Modulators and Drug Targets. Pharm. Res. 2011, 28, 3016–3029. [Google Scholar] [CrossRef]
- Esau, C.; Davis, S.; Murray, S.F.; Yu, X.X.; Pandey, S.K.; Pear, M.; Watts, L.; Booten, S.L.; Graham, M.; McKay, R.; et al. miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab. 2006, 3, 87–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krützfeldt, J.; Rajewsky, N.; Braich, R.; Rajeev, K.G.; Tuschl, T.; Manoharan, M.; Stoffel, M. Silencing of microRNAs in vivo with ‘antagomirs’. Nat. Cell Biol. 2005, 438, 685–689. [Google Scholar] [CrossRef]
- Jopling, C.L.; Schütz, S.; Sarnow, P. Position-Dependent Function for a Tandem MicroRNA miR-122-Binding Site Located in the Hepatitis C Virus RNA Genome. Cell Host Microbe 2008, 4, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Henke, J.I.; Goergen, D.; Zheng, J.; Song, Y.; Schüttler, C.G.; Fehr, C.; Jünemann, C.; Niepmann, M. microRNA-122 stimulates translation of hepatitis C virus RNA. EMBO J. 2008, 27, 3300–3310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuhara, T.; Matsuura, Y. Role of miR-122 and lipid metabolism in HCV infection. J. Gastroenterol. 2012, 48, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Jopling, C.L.; Yi, M.; Lancaster, A.M.; Lemon, S.M.; Sarnow, P. Modulation of Hepatitis C Virus RNA Abundance by a Liver-Specific MicroRNA. Science 2005, 309, 1577–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajan, S.; Hutvagner, G. RNA-Based Therapeutics: From Antisense Oligonucleotides to miRNAs. Cells 2020, 9, 137. [Google Scholar] [CrossRef] [Green Version]
- Elmén, J.; Lindow, M.; Schütz, S.; Lawrence, M.; Petri, A.; Obad, S.; Lindholm, M.W.; Hedtjärn, M.; Hansen, H.F.; Berger, U.; et al. LNA-mediated microRNA silencing in non-human primates. Nat. Cell Biol. 2008, 452, 896–899. [Google Scholar] [CrossRef]
- Kojima, S.; Gatfield, D.; Esau, C.C.; Green, C.B. MicroRNA-122 Modulates the Rhythmic Expression Profile of the Circadian Deadenylase Nocturnin in Mouse Liver. PLoS ONE 2010, 5, e11264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stelma, F.; Van Der Ree, M.H.M.H.; Sinnige, M.J.; Brown, A.; Swadling, L.; De Vree, J.M.L.; Willemse, S.B.; Van Der Valk, M.; Grint, P.; Neben, S.; et al. Immune phenotype and function of natural killer and T cells in chronic hepatitis C patients who received a single dose of anti-MicroRNA-122, RG-101. Hepatology 2017, 66, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshioka, K.; Kunieda, T.; Asami, Y.; Guo, H.; Miyata, H.; Yoshida-Tanaka, K.; Sujino, Y.; Piao, W.; Kuwahara, H.; Nishina, K.; et al. Highly efficient silencing of microRNA by heteroduplex oligonucleotides. Nucleic Acids Res. 2019, 47, 7321–7332. [Google Scholar] [CrossRef] [Green Version]
- Obad, S.; Dos Santos, C.O.; Petri, A.; Heidenblad, M.; Broom, O.; Ruse, C.; Fu, C.; Lindow, M.; Stenvang, J.; Straarup, E.M.; et al. Silencing of microRNA families by seed-targeting tiny LNAs. Nat. Genet. 2011, 43, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Rottiers, V.; Obad, S.; Petri, A.; McGarrah, R.; Lindholm, M.W.; Black, J.C.; Sinha, S.; Goody, R.J.; Lawrence, M.S.; Delemos, A.S.; et al. Pharmacological Inhibition of a MicroRNA Family in Nonhuman Primates by a Seed-Targeting 8-Mer AntimiR. Sci. Transl. Med. 2013, 5, 212ra162. [Google Scholar] [CrossRef] [Green Version]
- Debacker, A.J.; Voutila, J.; Catley, M.; Blakey, D.; Habib, N. Delivery of Oligonucleotides to the Liver with GalNAc: From Research to Registered Therapeutic Drug. Mol. Ther. 2020, 28, 1759–1771. [Google Scholar] [CrossRef]
- Yamamoto, T.; Sawamura, M.; Wada, F.; Harada-Shiba, M.; Obika, S. Serial incorporation of a monovalent GalNAc phosphoramidite unit into hepatocyte-targeting antisense oligonucleotides. Bioorganic Med. Chem. 2016, 24, 26–32. [Google Scholar] [CrossRef]
- Rajeev, K.G.; Nair, J.K.; Jayaraman, M.; Charisse, K.; Taneja, N.; O’Shea, J.; Willoughby, J.L.S.; Yucius, K.; Nguyen, T.; Shulga-Morskaya, S.; et al. Hepatocyte-Specific Delivery of siRNAs Conjugated to Novel Non-nucleosidic TrivalentN-Acetylgalactosamine Elicits Robust Gene Silencing in Vivo. ChemBioChem 2015, 16, 903–908. [Google Scholar] [CrossRef]
- Matsuda, S.; Keiser, K.; Nair, J.K.; Charisse, K.; Manoharan, R.M.; Kretschmer, P.; Peng, C.G.; Kel’In, A.V.; Kandasamy, P.; Willoughby, J.L.; et al. siRNA Conjugates Carrying Sequentially Assembled Trivalent N-Acetylgalactosamine Linked Through Nucleosides Elicit Robust Gene Silencing In Vivo in Hepatocytes. ACS Chem. Biol. 2015, 10, 1181–1187. [Google Scholar] [CrossRef]
- Yamamoto, T.; Sawamura, M.; Terada, C.; Kashiwada, K.; Wada, F.; Yamayoshi, A.; Obika, S.; Harada-Shiba, M. Effect of modular conjugation strategy forN-acetylgalactosamine-targeted antisense oligonucleotides. Nucleosides Nucleotides Nucleic Acids 2020, 39, 109–118. [Google Scholar] [CrossRef]
- Wada, F.; Yamamoto, T.; Ueda, T.; Sawamura, M.; Wada, S.; Harada-Shiba, M.; Obika, S. Cholesterol–GalNAc Dual Conjugation Strategy for Reducing Renal Distribution of Antisense Oligonucleotides. Nucleic Acid Ther. 2018, 28, 50–57. [Google Scholar] [CrossRef]
- Yamamoto, T.; Terada, C.; Kashiwada, K.; Yamayoshi, A.; Harada-Shiba, M.; Obika, S. Synthesis of MonovalentN-Acetylgalactosamine Phosphoramidite for Liver-Targeting Oligonucleotides. Curr. Protoc. Nucleic Acid Chem. 2019, 78, e99. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Vaud, S.; Zhelkovsky, A.M.; Posfai, J.; McReynolds, L.A. Sensitive and specific miRNA detection method using SplintR Ligase. Nucleic Acids Res. 2016, 44, e116. [Google Scholar] [CrossRef] [PubMed]
- Elmén, J.; Lindow, M.; Silahtaroglu, A.; Bak, M.; Christensen, M.; Lind-Thomsen, A.; Hedtjärn, M.; Hansen, J.B.; Hansen, H.F.; Straarup, E.M.; et al. Antagonism of microRNA-122 in mice by systemically administered LNA-antimiR leads to up-regulation of a large set of predicted target mRNAs in the liver. Nucleic Acids Res. 2007, 36, 1153–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sewing, S.; Gubler, M.; Gérard, R.; Avignon, B.; Mueller, Y.; Braendli-Baiocco, A.; Odin, M.; Moisan, A. GalNAc Conjugation Attenuates the Cytotoxicity of Antisense Oligonucleotide Drugs in Renal Tubular Cells. Mol. Ther. Nucleic Acids 2019, 14, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Janas, M.M.; Schlegel, M.K.; Harbison, C.E.; Yilmaz, V.O.; Jiang, Y.; Parmar, R.; Zlatev, I.; Castoreno, A.; Xu, H.; Shulga-Morskaya, S.; et al. Selection of GalNAc-conjugated siRNAs with limited off-target-driven rat hepatotoxicity. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Prakash, T.P.; Graham, M.J.; Yu, J.; Carty, R.; Low, A.; Chappell, A.; Schmidt, K.; Zhao, C.; Aghajan, M.; Murray, H.F.; et al. Targeted delivery of antisense oligonucleotides to hepatocytes using triantennary N-acetyl galactosamine improves potency 10-fold in mice. Nucleic Acids Res. 2014, 42, 8796–8807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Post, N.; Yu, R.; Greenlee, S.; Gaus, H.; Hurh, E.; Matson, J.; Wang, Y. Metabolism and Disposition of Volanesorsen, a 2′-O-(2 methoxyethyl) Antisense Oligonucleotide, Across Species. Drug Metab. Dispos. 2019, 47, 1164–1173. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.A.; Geary, R.S.; Levin, A.A. Plasma Protein Binding of an Antisense Oligonucleotide Targeting Human ICAM-1 (ISIS 2302). Oligonucleotides 2006, 16, 169–180. [Google Scholar] [CrossRef]
- Kilanowska, A.; Studzińska, S. In vivoandin vitrostudies of antisense oligonucleotides—A review. RSC Adv. 2020, 10, 34501–34516. [Google Scholar] [CrossRef]
- Huang, Y.; Zou, Q.; Wang, S.P.; Tang, S.M.; Zhang, G.Z.; Shen, X.J. The discovery approaches and detection methods of microRNAs. Mol. Biol. Rep. 2011, 38, 4125–4135. [Google Scholar] [CrossRef] [PubMed]
- Østergaard, M.E.; Yu, J.; Kinberger, G.A.; Wan, W.B.; Migawa, M.T.; Vasquez, G.; Schmidt, K.; Gaus, H.J.; Murray, H.M.; Low, A.; et al. Efficient Synthesis and Biological Evaluation of 5′-GalNAc Conjugated Antisense Oligonucleotides. Bioconjugate Chem. 2015, 26, 1451–1455. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.-P.; Kent, K.; Bird, J.; Fishback, J.; Froehler, B. Modified deoxyoligonucleotides stable to exonuclease degradation in serum. Nucleic Acids Res. 1991, 19, 747–750. [Google Scholar] [CrossRef] [Green Version]
- Burel, S.A.; Hart, C.E.; Cauntay, P.; Hsiao, J.; Machemer, T.; Katz, M.; Watt, A.T.; Bui, H.-H.; Younis, H.; Sabripour, M.; et al. Hepatotoxicity of high affinity gapmer antisense oligonucleotides is mediated by RNase H1 dependent promiscuous reduction of very long pre-mRNA transcripts. Nucleic Acids Res. 2016, 44, 2093–2109. [Google Scholar] [CrossRef] [Green Version]
- Swayze, E.E.; Siwkowski, A.M.; Wancewicz, E.V.; Migawa, M.T.; Wyrzykiewicz, T.K.; Hung, G.; Monia, B.P.; Bennett, C.F. Antisense oligonucleotides containing locked nucleic acid improve potency but cause significant hepatotoxicity in animals. Nucleic Acids Res. 2006, 35, 687–700. [Google Scholar] [CrossRef]
- Hagedorn, P.H.; Yakimov, V.; Ottosen, S.; Kammler, S.; Nielsen, N.F.; Høg, A.M.; Hedtjärn, M.; Meldgaard, M.; Møller, M.R.; Ørum, H.; et al. Hepatotoxic Potential of Therapeutic Oligonucleotides Can Be Predicted from Their Sequence and Modification Pattern. Nucleic Acid Ther. 2013, 23, 302–310. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Sequence (5′ to 3′) |
|---|---|
| CtrASO (a) (apoB-targeting cholesterol-lowering positive control) | HºHºHºG^C^a^t^t^g^g^t^a^t^T^C^A |
| tinyLNA | C^A^C^A^C^T^C^C |
| tL-5G3 | HºHºHºC^A^C^A^C^T^C^C |
| tinyCtr-5G3 (b) | HºHºHºT^C^A^T^A^C^T^A |
| tL-5G1 | HºC^A^C^A^C^T^C^C |
| tL-5G3-PO | HºHºHºCºAºCºAºCºTºCºC |
| tL-5G3-PS | H^H^H^C^A^C^A^C^T^C^C |
| tL-3G3 | C^A^C^A^C^T^C^CºHºHºH |
| Anti-miR122-probe | 5′-FºaºcºAºaºaºCºaºcºCºaºtºTºgºtº CºaºcºAºcºtºCºcºaºF-3′ |
| tinyLNA-f(b) | C^A^C^A^C^T^C^C^f |
| tL-5G3-f(b) | HºHºHºC^A^C^A^C^T^C^C^f |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamoto, T.; Mukai, Y.; Wada, F.; Terada, C.; Kayaba, Y.; Oh, K.; Yamayoshi, A.; Obika, S.; Harada–Shiba, M. Highly Potent GalNAc-Conjugated Tiny LNA Anti-miRNA-122 Antisense Oligonucleotides. Pharmaceutics 2021, 13, 817. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13060817
Yamamoto T, Mukai Y, Wada F, Terada C, Kayaba Y, Oh K, Yamayoshi A, Obika S, Harada–Shiba M. Highly Potent GalNAc-Conjugated Tiny LNA Anti-miRNA-122 Antisense Oligonucleotides. Pharmaceutics. 2021; 13(6):817. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13060817
Chicago/Turabian StyleYamamoto, Tsuyoshi, Yahiro Mukai, Fumito Wada, Chisato Terada, Yukina Kayaba, Kaho Oh, Asako Yamayoshi, Satoshi Obika, and Mariko Harada–Shiba. 2021. "Highly Potent GalNAc-Conjugated Tiny LNA Anti-miRNA-122 Antisense Oligonucleotides" Pharmaceutics 13, no. 6: 817. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13060817