
Recent Advances of Nanotechnology-Facilitated Bacteria-Based Drug and Gene Delivery Systems for Cancer Treatment

 ,
,
Abstract
:1. Introduction
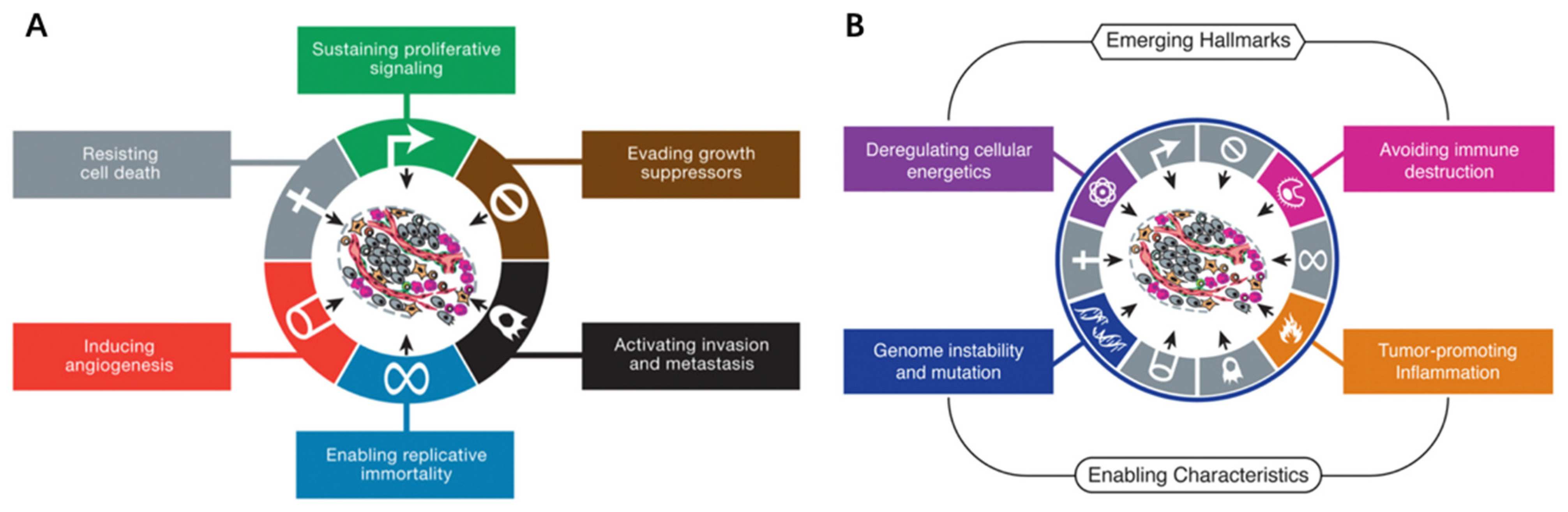
2. Cancer Hallmarks and Targeted Therapy
3. Bacteria, an Old Player against Cancer
3.1. Development of Bacterial Therapy against Cancer
3.2. Main Mechanisms of Bacterial Therapy
3.2.1. Tumor-Targeting Mechanisms
3.2.2. Therapeutic Mechanisms
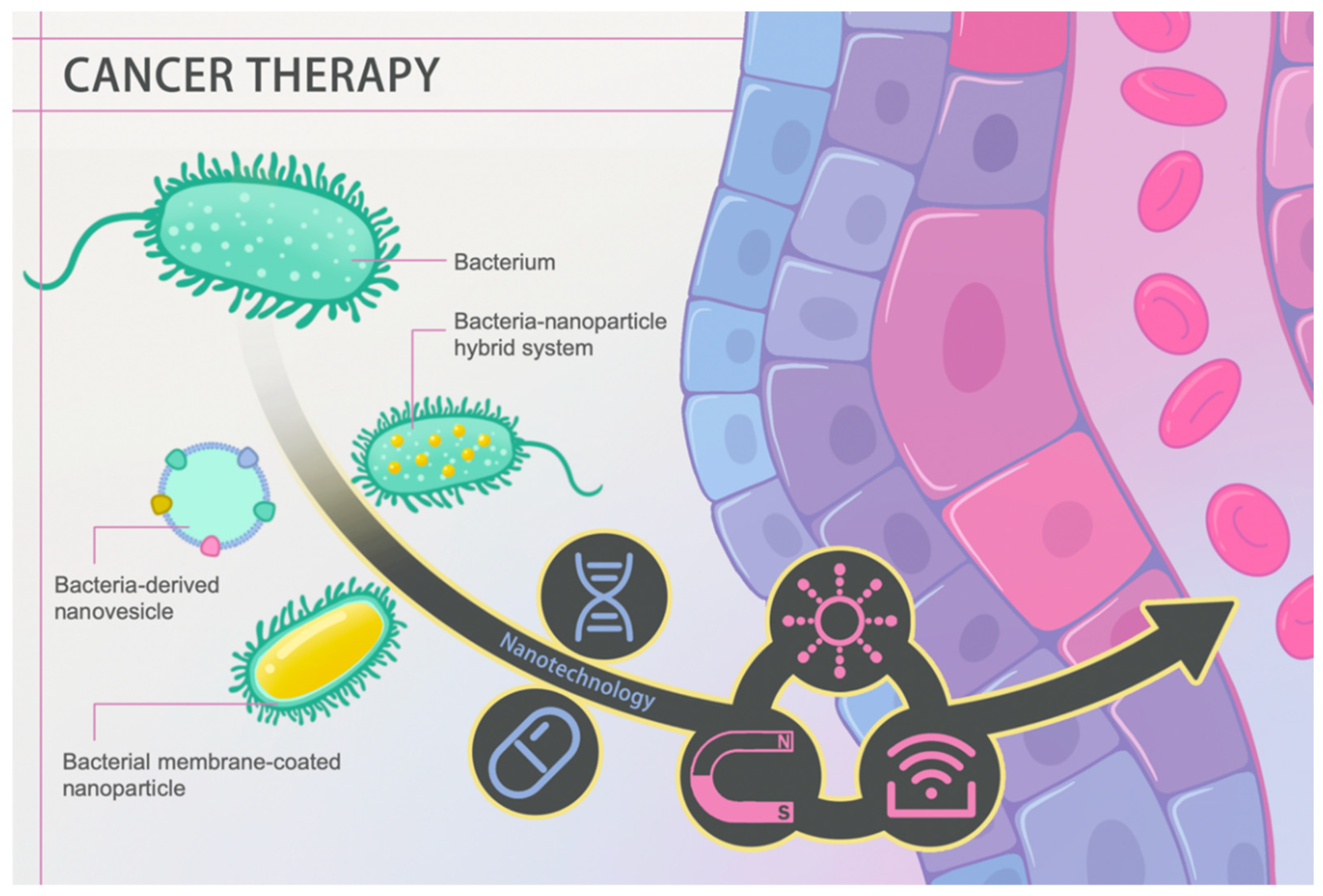
4. A New Role for the Old Player
4.1. Bacterial Membrane-Based Nanoformulations against Cancer
4.1.1. Bacteria-Derived Nanovesicles as Drug and Gene Delivery Systems
4.1.2. Other Functional Properties of Bacteria-Derived Nanovesicles
4.2. Bacteria–Nanoparticle Hybrid System
4.2.1. Drug and Gene Delivery
4.2.2. Other Functional Properties
5. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Nagai, H.; Kim, Y.H. Cancer Prevention from the Perspective of Global Cancer Burden Patterns. J. Thorac. Dis. 2017, 9, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S. Cisplatin: The First Metal Based Anticancer Drug. Bioorganic Chem. 2019, 88, 102925. [Google Scholar] [CrossRef] [PubMed]
- Minton, N.P. Clostridia in Cancer Therapy. Nat. Rev. Microbiol. 2003, 1, 237–242. [Google Scholar] [CrossRef]
- Faguet, G.B. A Brief History of Cancer: Age-Old Milestones Underlying Our Current Knowledge Database: A Brief History of Cancer. Int. J. Cancer 2015, 136, 2022–2036. [Google Scholar] [CrossRef] [PubMed]
- Falzone, L.; Salomone, S.; Libra, M. Evolution of Cancer Pharmacological Treatments at the Turn of the Third Millennium. Front. Pharmacol. 2018, 9, 1300. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yu, J.; Lu, X.; He, X. Nanoparticle Systems Reduce Systemic Toxicity in Cancer Treatment. Nanomedicine 2016, 11, 103–106. [Google Scholar] [CrossRef]
- Scripture, D.; Figg, W.D.; Sparreboom, A. Paclitaxel chemotherapy: From empiricism to a mechanism-based formulation strategy. Ther Clin Risk Manag. 2005, 1, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Venditto, V.J.; Simanek, E.E. Cancer Therapies Utilizing the Camptothecins: A Review of the in vivo Literature. Mol. Pharm. 2010, 7, 307–349. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Anand, P.; Kunnumakara, A.B.; Sundaram, C.; Harikumar, K.B.; Tharakan, S.T.; Lai, O.S.; Sung, B.; Aggarwal, B.B. Cancer Is a Preventable Disease That Requires Major Lifestyle Changes. Pharm Res. 2008, 25, 2097–2116. [Google Scholar] [CrossRef]
- Arruebo, M.; Vilaboa, N.; Sáez-Gutierrez, B.; Lambea, J.; Tres, A.; Valladares, M.; González-Fernández, Á. Assessment of the Evolution of Cancer Treatment Therapies. Cancers 2011, 3, 3279–3330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, N.; Iqbal, N. Human Epidermal Growth Factor Receptor 2 (HER2) in Cancers: Overexpression and Therapeutic Implications. Mol. Biol. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bedard, P.L.; Hyman, D.M.; Davids, M.S.; Siu, L.L. Small Molecules, Big Impact: 20 Years of Targeted Therapy in Oncology. Lancet 2020, 395, 1078–1088. [Google Scholar] [CrossRef]
- Bozic, I.; Allen, B.; Nowak, M.A. Dynamics of Targeted Cancer Therapy. Trends Mol. Med. 2012, 18, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, D.E.; Minna, J.D. ALK Inhibition for Non-Small Cell Lung Cancer: From Discovery to Therapy in Record Time. Cancer Cell 2010, 18, 548–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.K.; Menezes, M.E.; Bhatia, S.; Wang, X.-Y.; Emdad, L.; Sarkar, D.; Fisher, P.B. Gene Therapies for Cancer: Strategies, Challenges and Successes: Gene therapies for cancer. J. Cell. Physiol. 2015, 230, 259–271. [Google Scholar] [CrossRef] [Green Version]
- Lacouture, M.; Sibaud, V. Toxic Side Effects of Targeted Therapies and Immunotherapies Affecting the Skin, Oral Mucosa, Hair, and Nails. Am. J. Clin. Derm. 2018, 19, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Raoult, D.; Mouffok, N.; Bitam, I.; Piarroux, R.; Drancourt, M. Plague: History and Contemporary Analysis. J. Infect. 2013, 66, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Pawelek, J.M.; Low, K.B.; Bermudes, D. Bacteria as Tumour-Targeting Vectors. Lancet Oncol. 2003, 4, 548–556. [Google Scholar] [CrossRef]
- Nauts, H.C.; Swift, W.E. The Treatment of Malignant Tumors by Bacterial Toxins as Developed by the Late William I3. Coley, M.D., Reviewed in the Light of Modern Research. Cancer Res. 1946, 6, 205–216. [Google Scholar] [PubMed]
- Guallar-Garrido, S.; Julián, E. Bacillus Calmette-Guérin (BCG) Therapy for Bladder Cancer: An Update. ITT 2020, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Clairmont, C.; Lee, K.C.; Pike, J.; Ittensohn, M.; Low, K.B.; Pawelek, J.; Bermudes, D.; Brecher, S.M.; Margitich, D.; Turnier, J.; et al. Biodistribution and Genetic Stability of the Novel Antitumor Agent VNP20009, a Genetically Modified Strain of Salmonella typhimurium. J. Infect Dis. 2000, 181, 1996–2002. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.B.; Freeze, B.S.; LaMarche, M.J.; Sager, J.; Kinzler, K.W.; Vogelstein, B. Discodermolide Analogues as the Chemical Component of Combination Bacteriolytic Therapy. Bioorg. Med. Chem. Lett. 2005, 15, 3623–3626. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.C.; Clarke, E.J.; Arkin, A.P.; Voigt, C.A. Environmentally Controlled Invasion of Cancer Cells by Engineered Bacteria. J. Mol. Biol. 2006, 355, 619–627. [Google Scholar] [CrossRef]
- Yu, B.; Yang, M.; Shi, L.; Yao, Y.; Jiang, Q.; Li, X.; Tang, L.-H.; Zheng, B.-J.; Yuen, K.-Y.; Smith, D.K.; et al. Explicit Hypoxia Targeting with Tumor Suppression by Creating an “Obligate” Anaerobic Salmonella Typhimurium Strain. Sci. Rep. 2012, 2, 436. [Google Scholar] [CrossRef] [Green Version]
- Orange, M.; Reuter, U.; Hobohm, U. Coley’s Lessons Remembered: Augmenting Mistletoe Therapy. Integr. Cancer 2016, 15, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Han, R.; Evanoff, D.P.; Chen, X. Interleukin-10 Inhibits Mycobacterium Bovis Bacillus Calmette-Guérin (BCG)-Induced Macrophage Cytotoxicity against Bladder Cancer Cells: IL-10 in BCG-Induced Macrophage Cytotoxicity. Clin. Exp. Immunol. 2010, 160, 359–368. [Google Scholar] [CrossRef]
- Yuksel, Z.S.; Buber, E.; Kocagoz, T.; Alp, A.; Saribas, Z.; Acan, N.L. Mycobacterial Strains That Stimulate the Immune System Most Efficiently as Candidates for the Treatment of Bladder Cancer. J. Mol. Microbiol. Biotechnol. 2011, 20, 24–28. [Google Scholar] [CrossRef]
- Forbes, N.S. Engineering the Perfect (Bacterial) Cancer Therapy. Nat. Rev. Cancer 2010, 10, 785–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Yang, M.; Ma, H.; Li, X.; Tan, X.; Li, S.; Yang, Z.; Hoffman, R.M. Targeted Therapy with a Salmonella Typhimurium Leucine-Arginine Auxotroph Cures Orthotopic Human Breast Tumors in Nude Mice. Cancer Res. 2006, 66, 7647–7652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mircetic, J.; Dietrich, A.; Paszkowski-Rogacz, M.; Krause, M.; Buchholz, F. Development of a Genetic Sensor That Eliminates P53 Deficient Cells. Nat. Commun. 2017, 8, 1463. [Google Scholar] [CrossRef] [Green Version]
- Ruder, W.C.; Lu, T.; Collins, J.J. Synthetic Biology Moving into the Clinic. Science 2011, 333, 1248–1252. [Google Scholar] [CrossRef] [Green Version]
- Pawelek, J.M.; Low, K.B.; Bermudes, D. Tumor-Targeted Salmonella as a Novel Anticancer Vector. Cancer Res. 1997, 57, 4537–4544. [Google Scholar] [PubMed]
- Chandra, D.; Jahangir, A.; Quispe-Tintaya, W.; Einstein, M.H.; Gravekamp, C. Myeloid-Derived Suppressor Cells Have a Central Role in Attenuated Listeria Monocytogenes-Based Immunotherapy against Metastatic Breast Cancer in Young and Old Mice. Br. J. Cancer 2013, 108, 2281–2290. [Google Scholar] [CrossRef] [Green Version]
- Heldin, C.-H.; Rubin, K.; Pietras, K.; Östman, A. High Interstitial Fluid Pressure—An Obstacle in Cancer Therapy. Nat. Rev. Cancer 2004, 4, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Duong, M.T.-Q.; Qin, Y.; You, S.-H.; Min, J.-J. Bacteria-Cancer Interactions: Bacteria-Based Cancer Therapy. Exp. Mol. Med. 2019, 51, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kasinskas, R.W.; Forbes, N.S. Salmonella Typhimurium Specifically Chemotax and Proliferate in Heterogeneous Tumor Tissue in Vitro. Biotechnol. Bioeng. 2006, 94, 710–721. [Google Scholar] [CrossRef]
- Kasinskas, R.W.; Forbes, N.S. Salmonella Typhimurium Lacking Ribose Chemoreceptors Localize in Tumor Quiescence and Induce Apoptosis. Cancer Res. 2007, 67, 3201–3209. [Google Scholar] [CrossRef] [Green Version]
- Forbes, N.S.; Munn, L.L.; Fukumura, D.; Jain, R.K. Sparse Initial Entrapment of Systemically Injected Salmonella Typhimurium Leads to Heterogeneous Accumulation within Tumors. Cancer Res. 2003, 63, 5188–5193. [Google Scholar]
- Uchugonova, A.; Zhang, Y.; Salz, R.; Liu, F.; Suetsugu, A.; Zhang, L.; Koenig, K.; Hoffman, R.M.; Zhao, M. Imaging the Different Mechanisms of Prostate Cancer Cell- Killing by Tumor-Targeting Salmonella Typhimurium A1-R. Anticancer. Res. 2015, 5, 5225–5229. [Google Scholar]
- Lai, X.-H.; Arencibia, I.; Johansson, A.; Wai, S.N.; Oscarsson, J.; Kalfas, S.; Sundqvist, K.-G.; Mizunoe, Y.; Sjöstedt, A.; Uhlin, B.E. Cytocidal and Apoptotic Effects of the ClyA Protein from Escherichia Coli on Primary and Cultured Monocytes and Macrophages. Infect. Immun. 2000, 68, 4363–4367. [Google Scholar] [CrossRef] [Green Version]
- Vannini, F.; Kashfi, K.; Nath, N. The Dual Role of INOS in Cancer. Redox Biol. 2015, 6, 334–343. [Google Scholar] [CrossRef] [Green Version]
- Charles, G. The Role of Nitric Oxide in Cancer. Cell Res. 2002, 12, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-E.; Phan, T.X.; Nguyen, V.H.; Dinh-Vu, H.-V.; Zheng, J.H.; Yun, M.; Park, S.-G.; Hong, Y.; Choy, H.E.; Szardenings, M.; et al. Salmonella Typhimurium Suppresses Tumor Growth via the Pro-Inflammatory Cytokine Interleukin-1β. Theranostics 2015, 5, 1328–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saccheri, F.; Pozzi, C.; Avogadri, F.; Barozzi, S.; Faretta, M.; Fusi, P.; Rescigno, M. Bacteria-Induced Gap Junctions in Tumors Favor Antigen Cross-Presentation and Antitumor Immunity. Sci. Transl. Med. 2010, 2, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Berraondo, P.; Sanmamed, M.F.; Ochoa, M.C.; Etxeberria, I.; Aznar, M.A.; Pérez-Gracia, J.L.; Rodríguez-Ruiz, M.E.; Ponz-Sarvise, M.; Castañón, E.; Melero, I. Cytokines in Clinical Cancer Immunotherapy. Br. J. Cancer 2019, 120, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Phan, T.X.; Nguyen, V.H.; Duong, M.T.-Q.; Hong, Y.; Choy, H.E.; Min, J.-J. Activation of Inflammasome by Attenuated Salmonella Typhimurium in Bacteria-Mediated Cancer Therapy: Bacteriotherapy and Inflammasome. Microbiol. Immunol. 2015, 59, 664–675. [Google Scholar] [CrossRef] [Green Version]
- Tsujimoto, H.; Uchida, T.; Efron, P.A.; Scumpia, P.O.; Verma, A.; Matsumoto, T.; Tschoeke, S.K.; Ungaro, R.F.; Ono, S.; Seki, S.; et al. Flagellin Enhances NK Cell Proliferation and Activation Directly and through Dendritic Cell-NK Cell Interactions. J. Leukoc. Biol. 2005, 78, 888–897. [Google Scholar] [CrossRef]
- Kupz, A.; Curtiss, R.; Bedoui, S.; Strugnell, R.A. In Vivo IFN-γ Secretion by NK Cells in Response to Salmonella Typhimurium Requires NLRC4 Inflammasomes. PLoS ONE 2014, 9, e97418. [Google Scholar] [CrossRef]
- Kramer, M.G.; Masner, M.; Ferreira, F.A.; Hoffman, R.M. Bacterial Therapy of Cancer: Promises, Limitations, and Insights for Future Directions. Front. Microbiol. 2018, 9, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhtar, M.; Bilal, M.; Rahdar, A.; Barani, M.; Arshad, R.; Behl, T.; Brisc, C.; Banica, F.; Bungau, S. Nanomaterials for Diagnosis and Treatment of Brain Cancer: Recent Updates. Chemosensors 2020, 8, 117. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering Precision Nanoparticles for Drug Delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef]
- Chen, F.; Cai, W. Nanomedicine for Targeted Photothermal Cancer Therapy: Where Are We Now? Nanomedicine 2015, 10, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paszko, E.; Ehrhardt, C.; Senge, M.O.; Kelleher, D.P.; Reynolds, J.V. Nanodrug Applications in Photodynamic Therapy. Photodiagnosis Photodyn. Ther. 2011, 8, 14–29. [Google Scholar] [CrossRef]
- Kobayashi, T. Cancer Hyperthermia Using Magnetic Nanoparticles. Biotechnol. J. 2011, 6, 1342–1347. [Google Scholar] [CrossRef]
- Wang, S.; Gao, J.; Li, M.; Wang, L.; Wang, Z. A Facile Approach for Development of a Vaccine Made of Bacterial Double-Layered Membrane Vesicles (DMVs). Biomaterials 2018, 187, 28–38. [Google Scholar] [CrossRef]
- Pérez-Cruz, C.; Delgado, L.; López-Iglesias, C.; Mercade, E. Outer-Inner Membrane Vesicles Naturally Secreted by Gram-Negative Pathogenic Bacteria. PLoS ONE 2015, 10, e0116896. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the Wall: Extracellular Vesicles in Gram-Positive Bacteria, Mycobacteria and Fungi. Nat. Rev. Microbiol. 2015, 13, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Pillai, J. Bacterial Membrane Vesicles as Novel Nanosystems for Drug Delivery. IJN 2017, 12, 6329–6341. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Cárcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive Cell Lysis as a Mechanism for the Biogenesis of Bacterial Membrane Vesicles and Biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Liu, J. Bacteria and Bacterial Derivatives as Drug Carriers for Cancer Therapy. J. Control. Release 2020, 326, 396–407. [Google Scholar] [CrossRef]
- Bitto, N.; Kaparakis-Liaskos, M. The Therapeutic Benefit of Bacterial Membrane Vesicles. IJMS 2017, 18, 1287. [Google Scholar] [CrossRef] [Green Version]
- Chronopoulos, A.; Kalluri, R. Emerging Role of Bacterial Extracellular Vesicles in Cancer. Oncogene 2020, 39, 6951–6960. [Google Scholar] [CrossRef]
- MacDiarmid, J.A.; Mugridge, N.B.; Weiss, J.C.; Phillips, L.; Burn, A.L.; Paulin, R.P.; Haasdyk, J.E.; Dickson, K.-A.; Brahmbhatt, V.N.; Pattison, S.T.; et al. Bacterially Derived 400 Nm Particles for Encapsulation and Cancer Cell Targeting of Chemotherapeutics. Cancer Cell 2007, 11, 431–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDiarmid, J.A.; Amaro-Mugridge, N.B.; Madrid-Weiss, J.; Sedliarou, I.; Wetzel, S.; Kochar, K.; Brahmbhatt, V.N.; Phillips, L.; Pattison, S.T.; Petti, C.; et al. Sequential Treatment of Drug-Resistant Tumors with Targeted Minicells Containing SiRNA or a Cytotoxic Drug. Nat. Biotechnol. 2009, 27, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.Y.; Park, H.T.; Dinh, N.T.H.; Choi, S.J.; Lee, J.; Kim, J.H.; Lee, S.-W.; Gho, Y.S. Bacterial Outer Membrane Vesicles Suppress Tumor by Interferon-γ-Mediated Antitumor Response. Nat. Commun. 2017, 8, 626. [Google Scholar] [CrossRef]
- Qing, S.; Lyu, C.; Zhu, L.; Pan, C.; Wang, S.; Li, F.; Wang, J.; Yue, H.; Gao, X.; Jia, R.; et al. Biomineralized Bacterial Outer Membrane Vesicles Potentiate Safe and Efficient Tumor Microenvironment Reprogramming for Anticancer Therapy. Adv. Mater. 2020, 32, 2002085. [Google Scholar] [CrossRef] [PubMed]
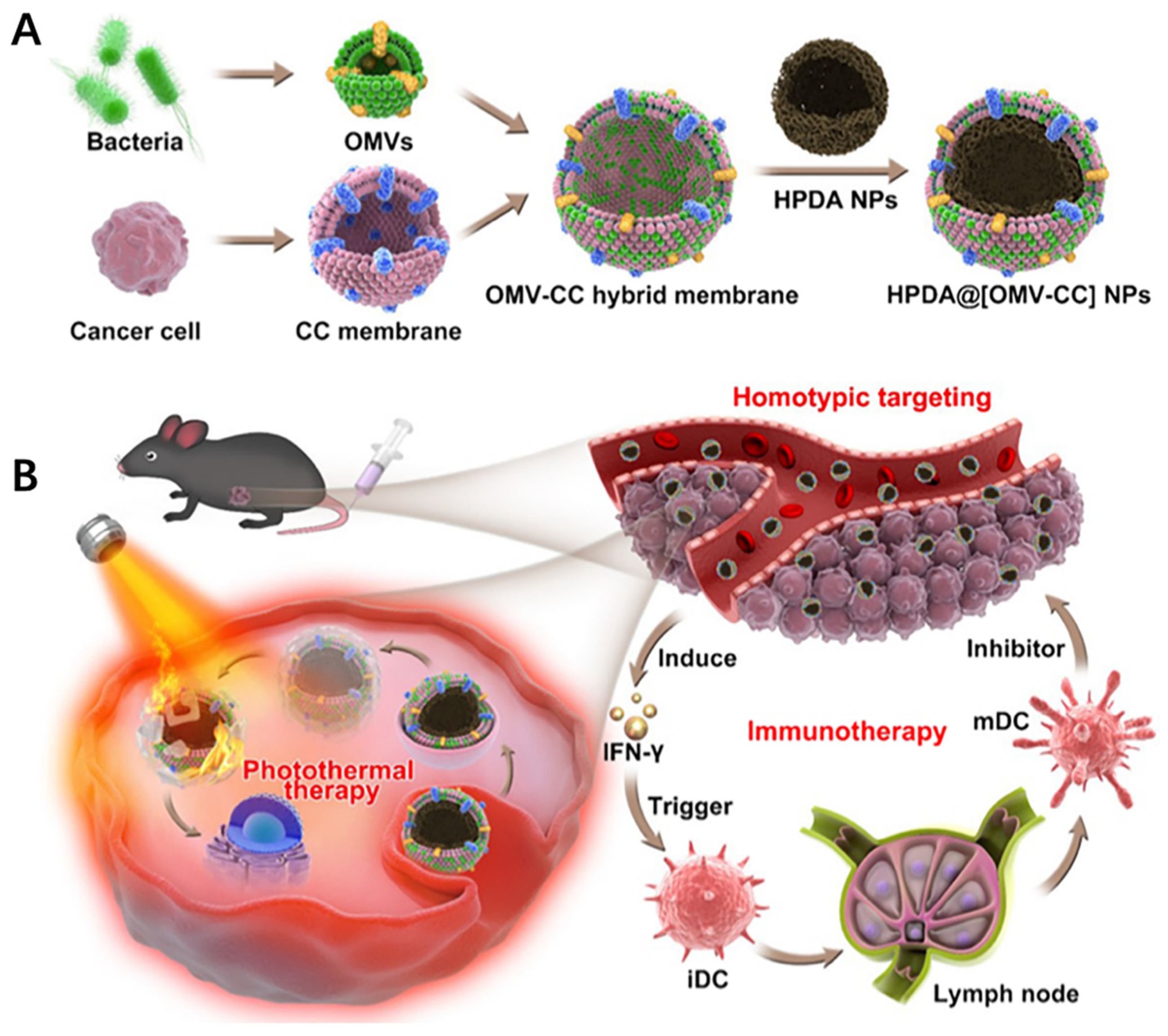
- Wang, D.; Liu, C.; You, S.; Zhang, K.; Li, M.; Cao, Y.; Wang, C.; Dong, H.; Zhang, X. Bacterial Vesicle-Cancer Cell Hybrid Membrane-Coated Nanoparticles for Tumor Specific Immune Activation and Photothermal Therapy. ACS Appl. Mater. Interfaces 2020, 12, 41138–41147. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.B.; Ye, M.; Carlson, P.M.; Jaquish, A.; Zangl, L.; Ma, B.; Wang, Y.; Arthur, I.; Xie, R.; Brown, R.J.; et al. Development of an In Situ Cancer Vaccine via Combinational Radiation and Bacterial-Membrane-Coated Nanoparticles. Adv. Mater. 2019, 31, 1902626. [Google Scholar] [CrossRef]
- Chen, Q.; Bai, H.; Wu, W.; Huang, G.; Li, Y.; Wu, M.; Tang, G.; Ping, Y. Bioengineering Bacterial Vesicle-Coated Polymeric Nanomedicine for Enhanced Cancer Immunotherapy and Metastasis Prevention. Nano Lett. 2020, 20, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Aly, R.G.; El-Enbaawy, M.I.; Abd El-Rahman, S.S.; Ata, N.S. Antineoplastic Activity of Salmonella Typhimurium Outer Membrane Nanovesicles. Exp. Cell Res. 2021, 399, 112423. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.Y.; Dinh, N.T.H.; Park, H.T.; Choi, S.J.; Hong, K.; Gho, Y.S. Bacterial Protoplast-Derived Nanovesicles for Tumor Targeted Delivery of Chemotherapeutics. Biomaterials 2017, 113, 68–79. [Google Scholar] [CrossRef]
- Gao, J.; Wang, S.; Dong, X.; Wang, Z. RGD-Expressed Bacterial Membrane-Derived Nanovesicles Enhance Cancer Therapy via Multiple Tumorous Targeting. Theranostics 2021, 11, 3301–3316. [Google Scholar] [CrossRef]
- Gujrati, V.; Kim, S.; Kim, S.-H.; Min, J.J.; Choy, H.E.; Kim, S.C.; Jon, S. Bioengineered Bacterial Outer Membrane Vesicles as Cell-Specific Drug-Delivery Vehicles for Cancer Therapy. ACS Nano 2014, 8, 1525–1537. [Google Scholar] [CrossRef]
- Peng, L.-H.; Wang, M.-Z.; Chu, Y.; Zhang, L.; Niu, J.; Shao, H.-T.; Yuan, T.-J.; Jiang, Z.-H.; Gao, J.-Q.; Ning, X.-H. Engineering Bacterial Outer Membrane Vesicles as Transdermal Nanoplatforms for Photo-TRAIL–Programmed Therapy against Melanoma. Sci. Adv. 2020, 6, eaba2735. [Google Scholar] [CrossRef]
- Huang, W.; Shu, C.; Hua, L.; Zhao, Y.; Xie, H.; Qi, J.; Gao, F.; Gao, R.; Chen, Y.; Zhang, Q.; et al. Modified Bacterial Outer Membrane Vesicles Induce Autoantibodies for Tumor Therapy. Acta Biomater. 2020, 108, 300–312. [Google Scholar] [CrossRef]
- Suh, S.; Jo, A.; Traore, M.A.; Zhan, Y.; Coutermarsh-Ott, S.L.; Ringel-Scaia, V.M.; Allen, I.C.; Davis, R.M.; Behkam, B. Nanoscale Bacteria-Enabled Autonomous Drug Delivery System (NanoBEADS) Enhances Intratumoral Transport of Nanomedicine. Adv. Sci. 2019, 6, 1801309. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Xu, D.; Gao, X.; Xiong, J.; Jiang, B.; Zhang, Y.; Wang, Y.; Tang, Y.; Chen, C.; Qiao, H.; et al. Nanoparticles Conjugated with Bacteria Targeting Tumors for Precision Imaging and Therapy. Biochem. Biophys. Res. Commun. 2019, 514, 1147–1153. [Google Scholar] [CrossRef] [PubMed]
- Durymanov, M.; Reineke, J. Non-Viral Delivery of Nucleic Acids: Insight into Mechanisms of Overcoming Intracellular Barriers. Front. Pharmacol. 2018, 9, 971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, A.B.; Chen, M.; Chen, C.-K.; Pfeifer, B.A.; Jones, C.H. Overcoming Gene-Delivery Hurdles: Physiological Considerations for Nonviral Vectors. Trends Biotechnol. 2016, 34, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Gisbert-Garzarán, M.; Lozano, D.; Matsumoto, K.; Komatsu, A.; Manzano, M.; Tamanoi, F.; Vallet-Regí, M. Designing Mesoporous Silica Nanoparticles to Overcome Biological Barriers by Incorporating Targeting and Endosomal Escape. ACS Appl. Mater. Interfaces. 2021, 13, 9656–9666. [Google Scholar] [CrossRef]
- Akin, D.; Sturgis, J.; Ragheb, K.; Sherman, D.; Burkholder, K.; Robinson, J. Paul.; Bhunia, A.K.; Mohammed, S.; Bashir, R. Bacteria-Mediated Delivery of Nanoparticles and Cargo into Cells. Nat. Nanotech. 2007, 2, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Celli, J.P.; Spring, B.Q.; Rizvi, I.; Evans, C.L.; Samkoe, K.S.; Verma, S.; Pogue, B.W.; Hasan, T. Imaging and Photodynamic Therapy: Mechanisms, Monitoring, and Optimization. Chem. Rev. 2010, 110, 2795–2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
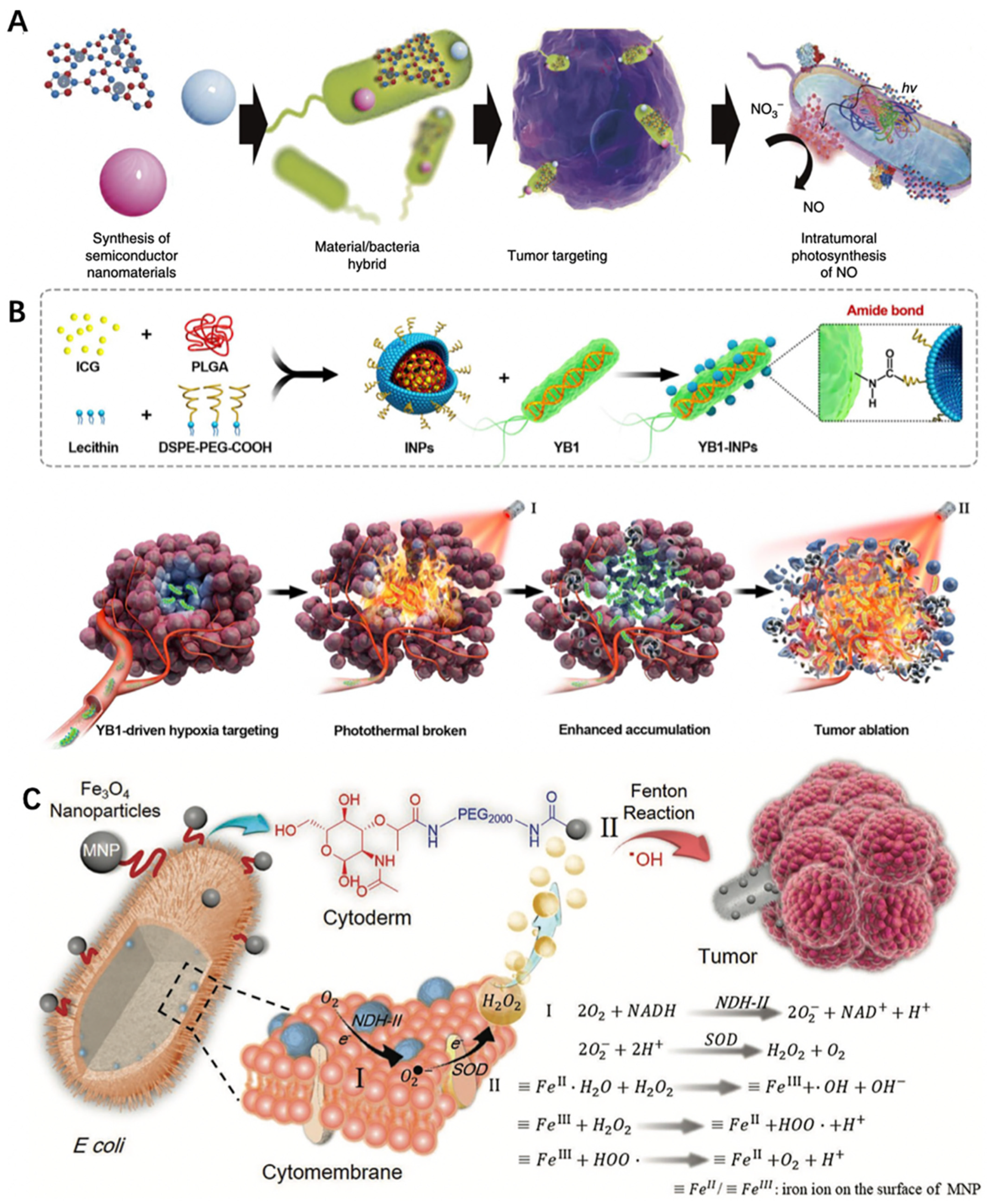
- Zheng, D.-W.; Chen, Y.; Li, Z.-H.; Xu, L.; Li, C.-X.; Li, B.; Fan, J.-X.; Cheng, S.-X.; Zhang, X.-Z. Optically-Controlled Bacterial Metabolite for Cancer Therapy. Nat. Commun. 2018, 9, 1680. [Google Scholar] [CrossRef]
- Chen, F.; Zang, Z.; Chen, Z.; Cui, L.; Chang, Z.; Ma, A.; Yin, T.; Liang, R.; Han, Y.; Wu, Z.; et al. Nanophotosensitizer-Engineered Salmonella Bacteria with Hypoxia Targeting and Photothermal-Assisted Mutual Bioaccumulation for Solid Tumor Therapy. Biomaterials 2019, 214, 119226. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Peng, M.; Wang, H.; Zheng, H.; Liu, Z.; Li, C.; Wang, X.; Liu, X.; Cheng, S.; Zhang, X. Engineered Bacterial Bioreactor for Tumor Therapy via Fenton-Like Reaction with Localized H2O2 Generation. Adv. Mater. 2019, 31, 1808278. [Google Scholar] [CrossRef] [PubMed]
- Han, H.S.; Choi, K.Y. Advances in Nanomaterial-Mediated Photothermal Cancer Therapies: Toward Clinical Applications. Biomedicines 2021, 9, 305. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Shen, X.; Gao, X.; Zhao, Y. Simultaneous Enzyme Mimicking and Chemical Reduction Mechanisms for Nanoceria as a Bio-Antioxidant: A Catalytic Model Bridging Computations and Experiments for Nanozymes. Nanoscale 2019, 11, 13289–13299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jin, Y.; Cui, H.; Yan, X.; Fan, K. Nanozyme-Based Catalytic Theranostics. RSC Adv. 2020, 10, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Zhuang, J.; Nie, L.; Zhang, J.; Zhang, Y.; Gu, N.; Wang, T.; Feng, J.; Yang, D.; Perrett, S.; et al. Intrinsic Peroxidase-like Activity of Ferromagnetic Nanoparticles. Nat. Nanotech. 2007, 2, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Chen, Y.; Shi, J. Nanocatalytic Medicine. Adv. Mater. 2019, 31, 1901778. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Yang, W.; Shao, Z.; Zhao, Y. Genetically Modified Bacteria for Targeted Phototherapy of Tumor. Biomaterials 2021, 272, 120809. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, J.; Wang, X.; Fan, J.; Liu, X.; Li, B.; Han, Z.; Cheng, S.; Zhang, X. Inhibition of Tumor Progression through the Coupling of Bacterial Respiration with Tumor Metabolism. Angew. Chem. Int. Ed. 2020, 59, 21562–21570. [Google Scholar] [CrossRef]
- Liu, L.; He, H.; Luo, Z.; Zhou, H.; Liang, R.; Pan, H.; Ma, Y.; Cai, L. In Situ Photocatalyzed Oxygen Generation with Photosynthetic Bacteria to Enable Robust Immunogenic Photodynamic Therapy in Triple-Negative Breast Cancer. Adv. Funct. Mater. 2020, 30, 1910176. [Google Scholar] [CrossRef]
- Felfoul, O.; Mohammadi, M.; Taherkhani, S.; de Lanauze, D.; Zhong Xu, Y.; Loghin, D.; Essa, S.; Jancik, S.; Houle, D.; Lafleur, M.; et al. Magneto-Aerotactic Bacteria Deliver Drug-Containing Nanoliposomes to Tumour Hypoxic Regions. Nat. Nanotech. 2016, 11, 941–947. [Google Scholar] [CrossRef]
- Nedeva, C.; Menassa, J.; Puthalakath, H. Sepsis: Inflammation Is a Necessary Evil. Front. Cell Dev. Biol. 2019, 7, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Bacteria | Cancer Type | Brief Description | Ref. |
|---|---|---|---|---|
| 1868 | Streptococcus pyogenes | Sarcoma | First use of bacteria in cancer treatment | [20] |
| 1891 | Streptococcus pyogenes | Malignant sarcoma | Coley’s toxins | [21] |
| 1989 | Mycobacterium bovis | Bladder cancer | Bacillus Calmette–Guerin vaccine (BCG) approved by the FDA | [22] |
| 2000 | Salmonella typhimurium VNP20009 | Solid tumor | Deletion of the purI and msbB genes which reduce the virulence and the risk of septic shock | [23] |
| 2005 | Clostridium novyi-NT | HCT116 colorectal cancer | Combination of bacterial therapy and traditional drug therapy | [24] |
| 2006 | Escherichia coli | HeLa, HepG2, and U2OS cell lines | Characterization of invasin from Yersinia pseudotuberculosis as an output module | [25] |
| 2011 | Salmonella Typhimurium SL7207 | Colorectal carcinoma | Engineered to survive only in anaerobic conditions without otherwise affecting its functions | [26] |
| Membrane Source | Cancer Type | Membrane Type | Cargo | Efficacy | Ref. |
|---|---|---|---|---|---|
| Salmonella | B16F10 and 4T1 tumors | OMV | Tegafur@F127 nanomicelles |
| [71] |
| Ehrlich ascites carcinoma (EAC) | OMV | Paclitaxel |
| [72] | |
| Escherichia coli | Human lung carcinoma A459 cells | Protoplast-derived nanovesicles | Doxorubicin |
| [73] |
| B16F10 tumor | DMV | Doxorubicin |
| [74] | |
| HER2-overexpressing HCC1954 cells | OMV | siRNA |
| [75] | |
| CT26 and 4T1 tumors | OMV | ICG |
| [68] | |
| B16F10 tumor | OMV | ICG |
| [76] | |
| TC-1 and B16F10 tumors | OMV | BFGF |
| [77] |
| Bacterium | Cancer Type | Nanoparticle | Cargo | Efficacy/Therapeutic Mechanism | Ref. |
|---|---|---|---|---|---|
| S. typhimurium VNP20009 | 4T1 tumor | PLGA | / | Remarkable (up to 100-fold) enhancement of nanoparticle retention and distribution in solid tumors | [78] |
| Bifidobacterium longum | MDA-MB-231 breast tumor | PLGA | Low-boiling-point perfluorohexane (PFH) | Combination of diagnostic and therapeutic efficacyRealization of high-intensity focused ultrasound therapy against cancer | [79] |
| L. monocytogenes | MCF-7, HT29, KB, HepG-2 cancer cells | Polystyrene nanoparticles | GFP-encoding plasmid DNA | High resistance toward the acidic endosome environment and intracellular enzymes and successful delivery of genes into the nucleus | [83] |
| Escherichia coli | 4T1 and CT26 tumors | Carbon nitride (C3N4) semiconductor nanomaterials | / | Achievement of approximately 80% tumor regression superior than with E. coli alone (~20%) | [85] |
| Salmonella typhimurium YB1 | MB49 tumor | PLGA | ICG | Highly efficient photothermal ability to eradicate established solid tumors without relapse | [86] |
| Escherichia coli MG1655 | CT26 tumor | Magnetic Fe3O4 nanoparticles | / | Achievement of effective tumor colonization and realization of a self-supplied therapeutic Fenton-like reaction to cure cancer without an additional H2O2 source | [87] |
| Escherichia coli | HOS, MG63, and U2OS cancer cells | Polydopamine nanoparticles | Ce6 | An ability to provide catalase and convert endogenic hydrogen peroxide into oxygen for subsequent photodynamic therapy | [93] |
| Shewanella oneidensis MR-1 | CT26 tumor | Manganese dioxide nanoflowers | / | MnO2 serves as electron acceptor, tumor metabolite lactic acid performs as an electron donor, resulting in continuous consumption of lactic acid in cancer cells | [94] |
| Synechococcus 7942 | 4T1 tumor | Human serum albumin nanoparticles | ICG |
In situ photocatalyzed oxygen generation enabling robust immunogenic PDT against tumor growth and metastasis | [95] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, C.; Ji, Z.; Ma, J.; Ding, Z.; Shen, J.; Wang, Q. Recent Advances of Nanotechnology-Facilitated Bacteria-Based Drug and Gene Delivery Systems for Cancer Treatment. Pharmaceutics 2021, 13, 940. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13070940
Zhu C, Ji Z, Ma J, Ding Z, Shen J, Wang Q. Recent Advances of Nanotechnology-Facilitated Bacteria-Based Drug and Gene Delivery Systems for Cancer Treatment. Pharmaceutics. 2021; 13(7):940. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13070940
Chicago/Turabian StyleZhu, Chaojie, Zhiheng Ji, Junkai Ma, Zhijie Ding, Jie Shen, and Qiwen Wang. 2021. "Recent Advances of Nanotechnology-Facilitated Bacteria-Based Drug and Gene Delivery Systems for Cancer Treatment" Pharmaceutics 13, no. 7: 940. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13070940