
Comparison of Pharmacokinetics and Anti-Pulmonary Fibrosis-Related Effects of Sulforaphane and Sulforaphane N-acetylcysteine

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
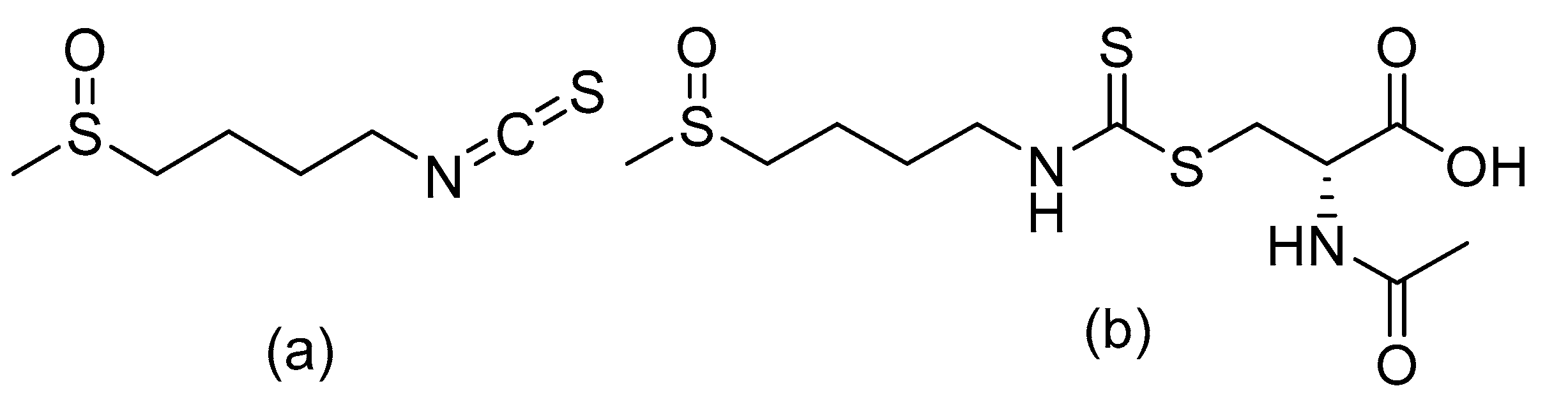
2.1. Chemicals and Reagents
2.2. In Vitro Anti-Lung Fibrosis Effects
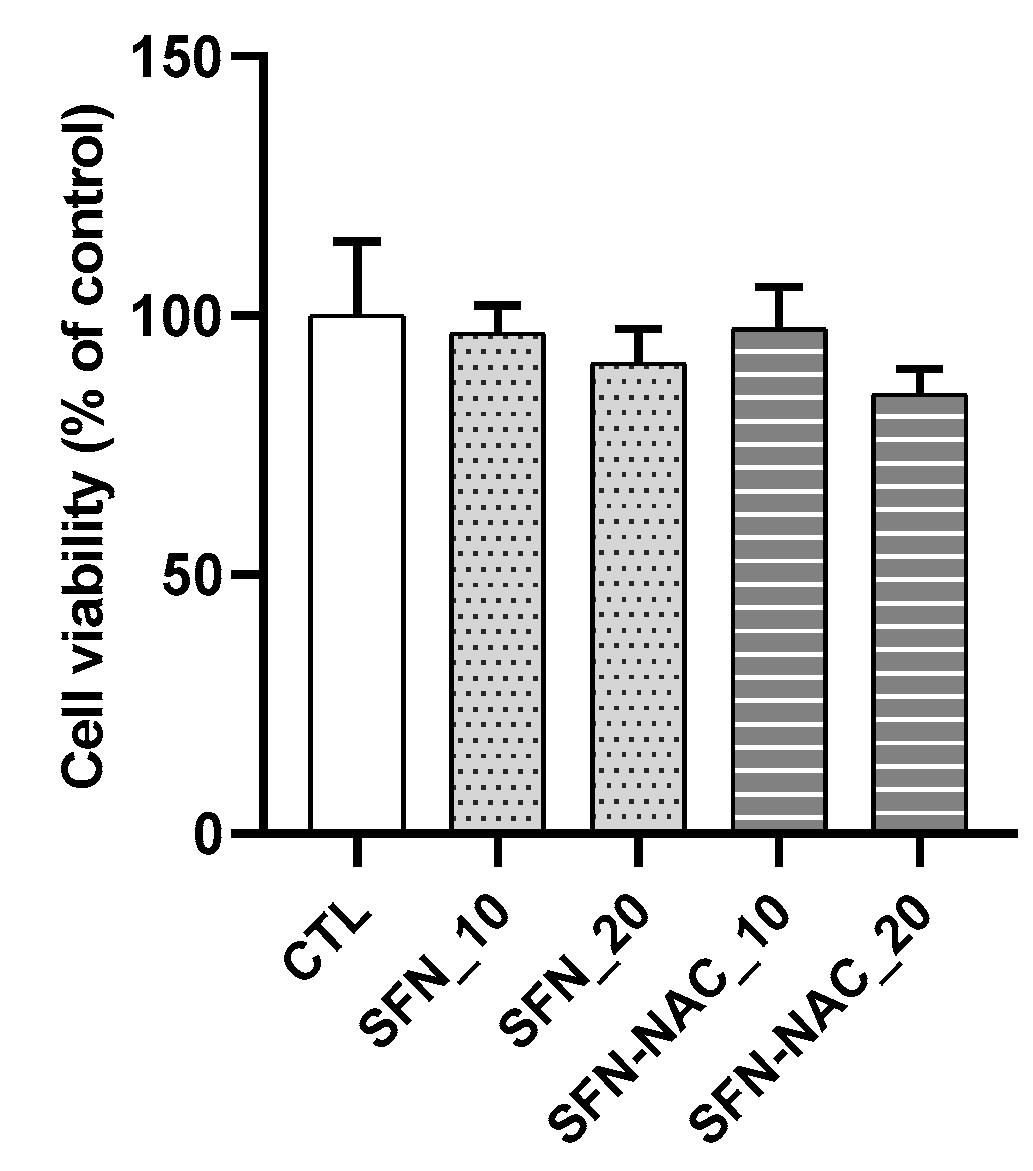
2.2.1. Cell Viability Assay
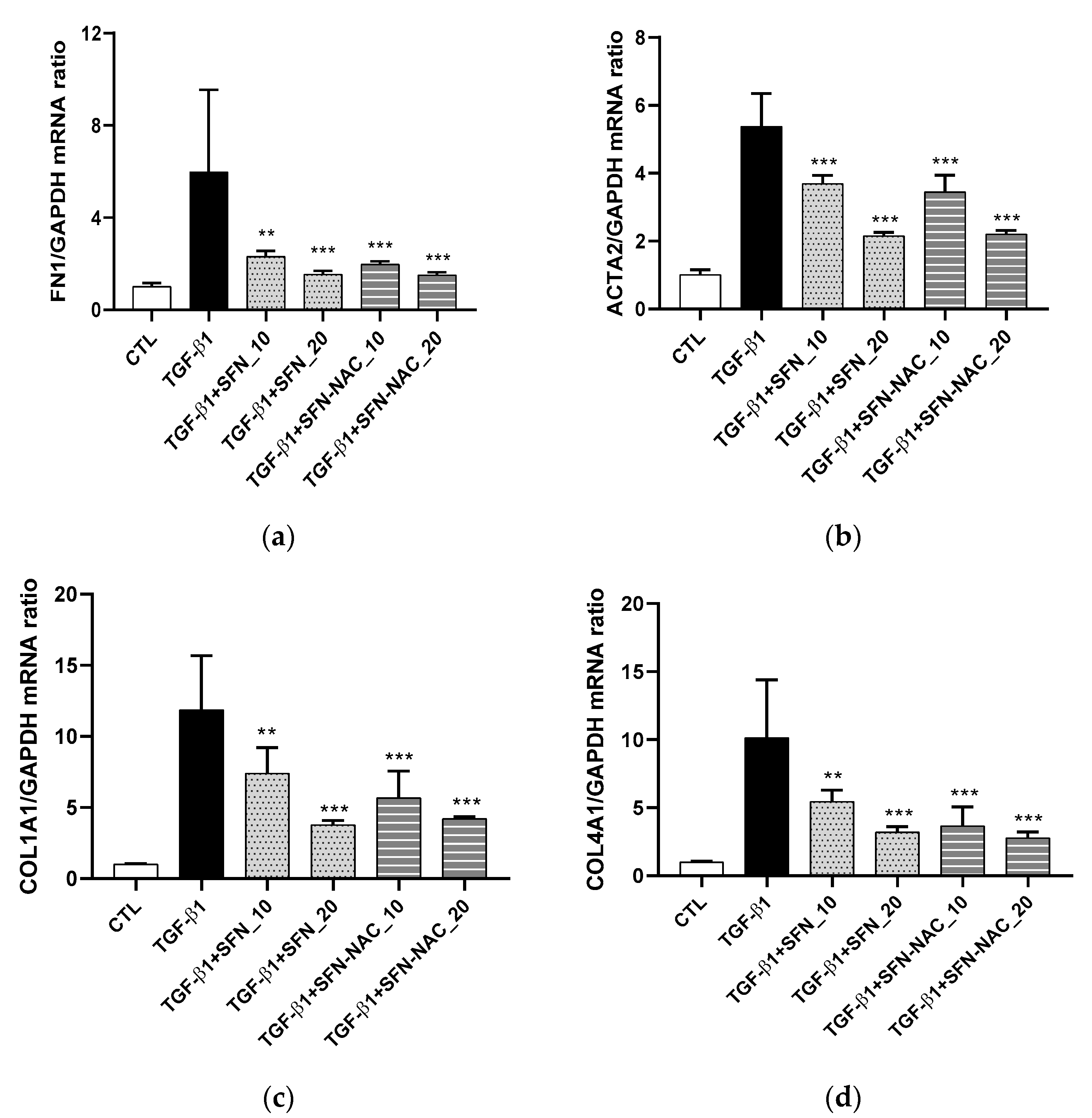
2.2.2. Quantitative Reverse Transcription-PCR (qRT-PCR) Analysis
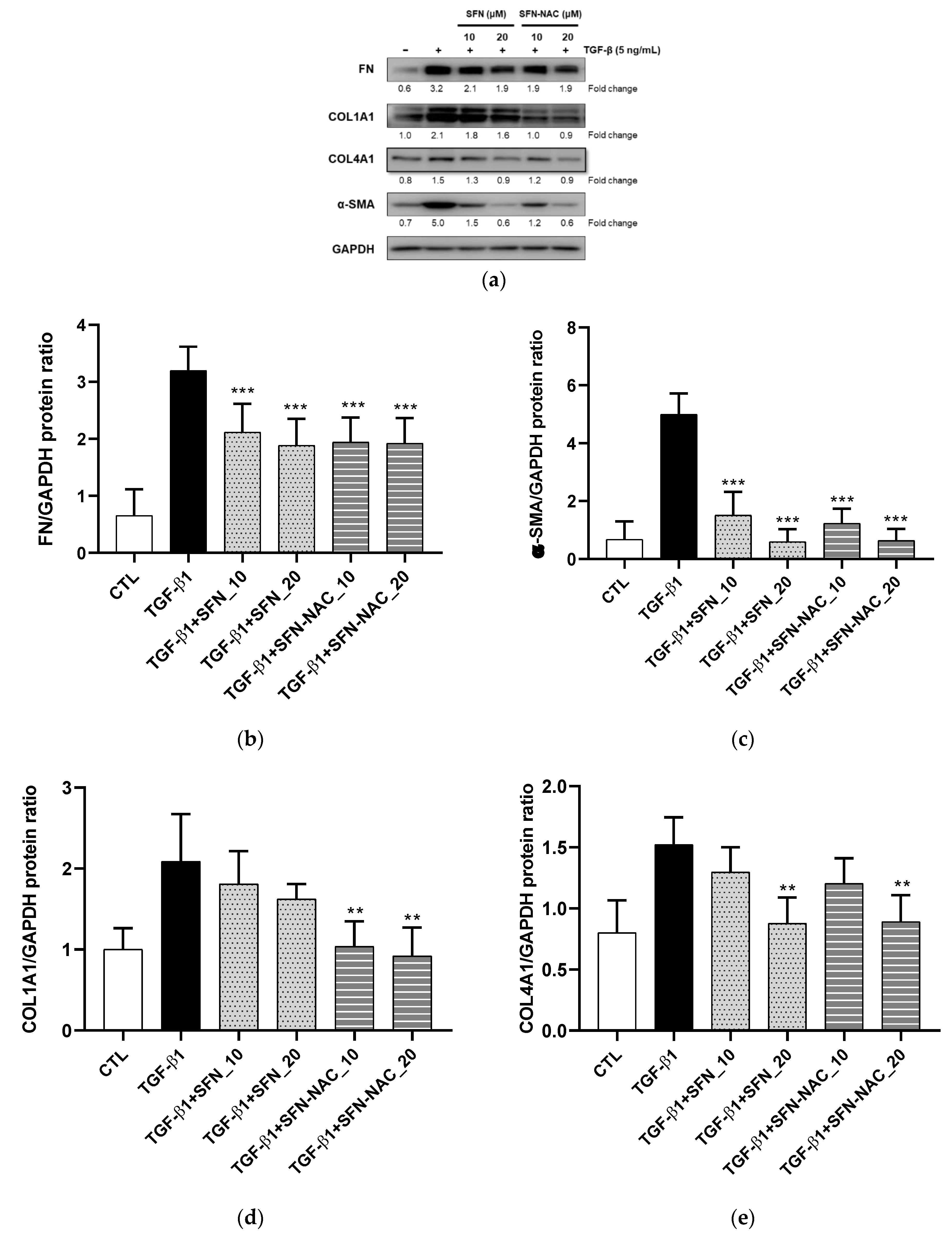
2.2.3. Western Blot Analysis
2.3. In Vitro Pharmacokinetic Study
2.3.1. Plasma Stability Assay
2.3.2. Liver Microsomal Metabolic Stability
2.3.3. Apparent Permeability for Direction of Absorption in Caco-2 Cells
2.4. In Vivo Pharmacokinetic Study at a Micro-Dose Range
2.4.1. Animal Study
2.4.2. Pharmacokinetic Analysis
2.5. LC-MS/MS Analysis
2.6. Statistical Analyses
3. Results
3.1. In Vitro Anti-Fibrosis Effects of SFN and SFN-NAC
3.2. In Vitro Pharmacokinetic Properties
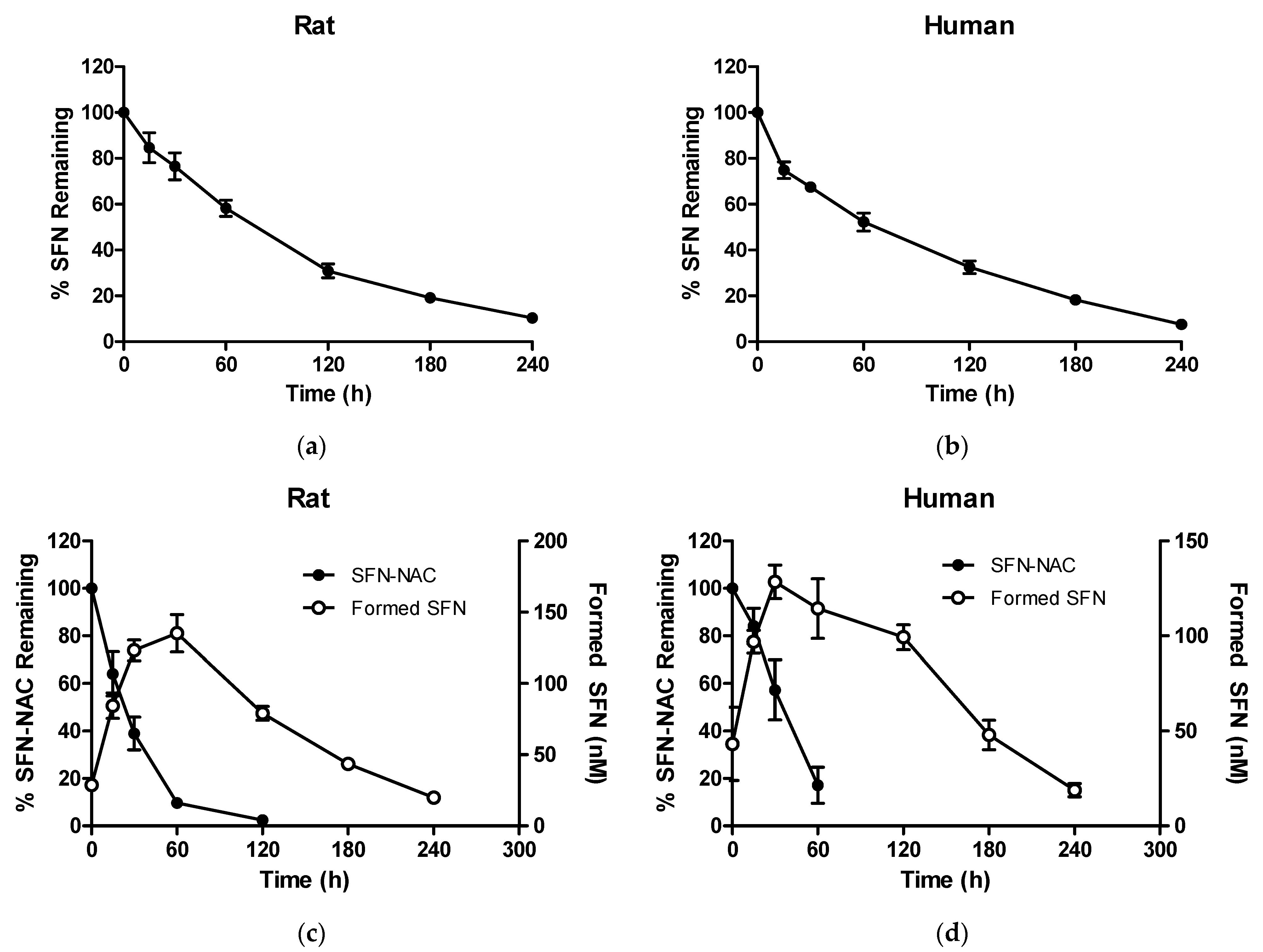
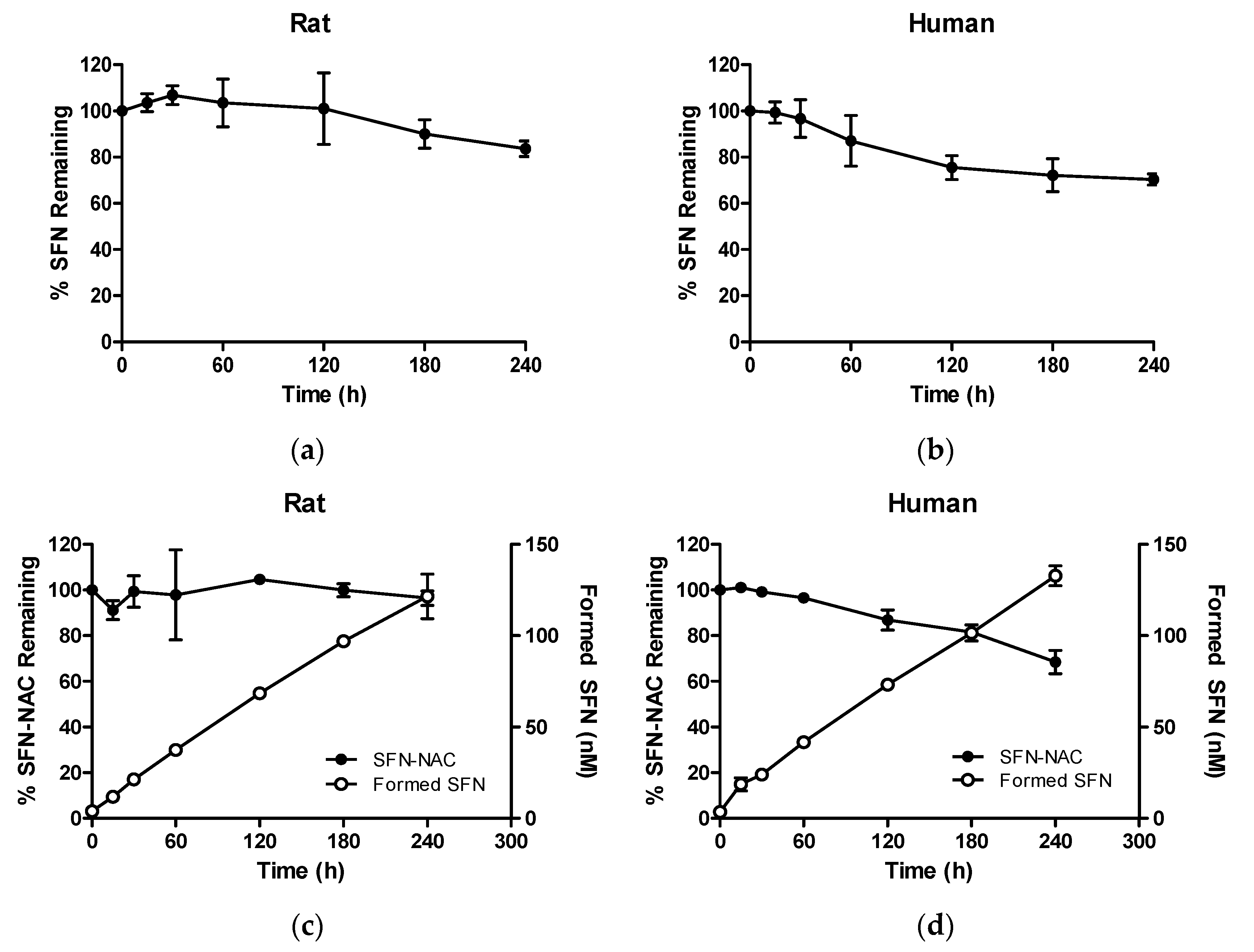
3.2.1. Metabolic Stability of SFN and SFN-NAC
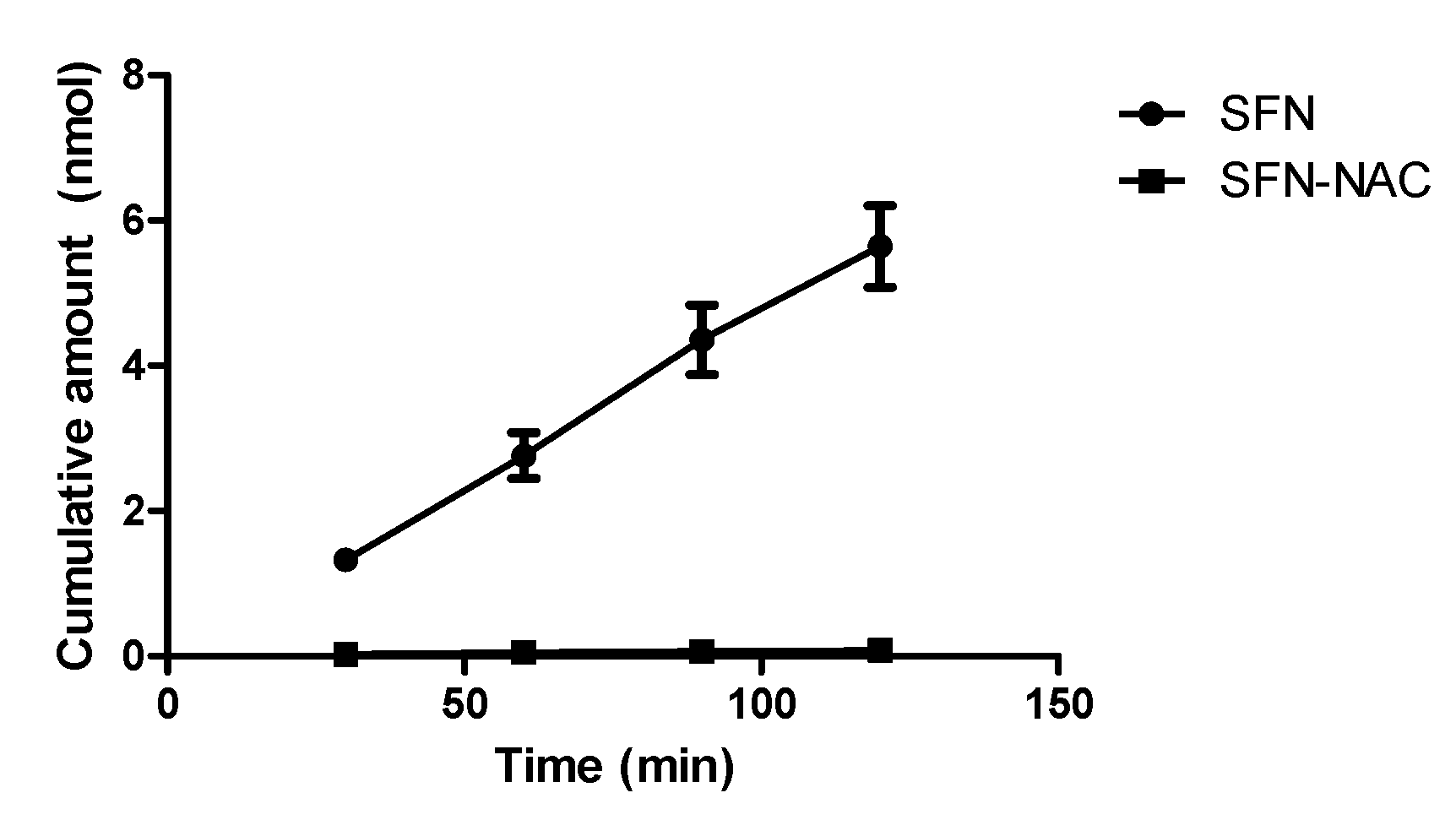
3.2.2. Apparent Permeability for Absorption Direction in Caco-2 Cells
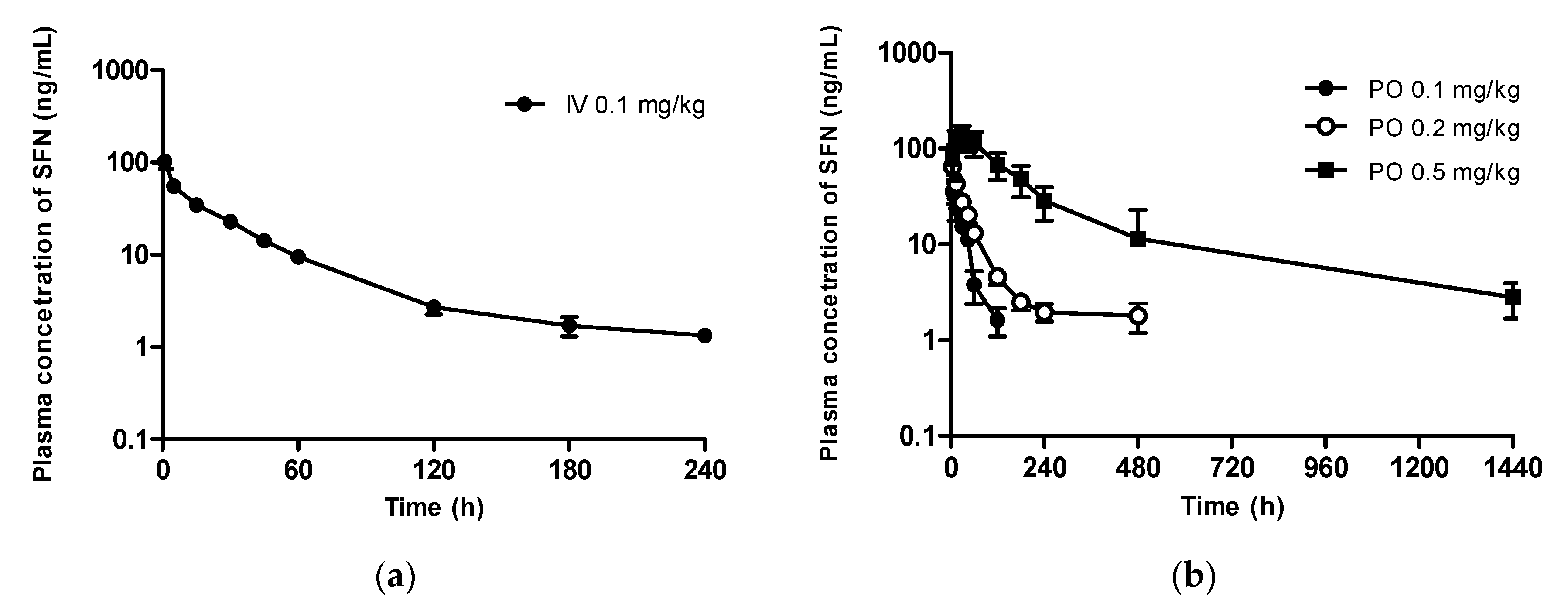
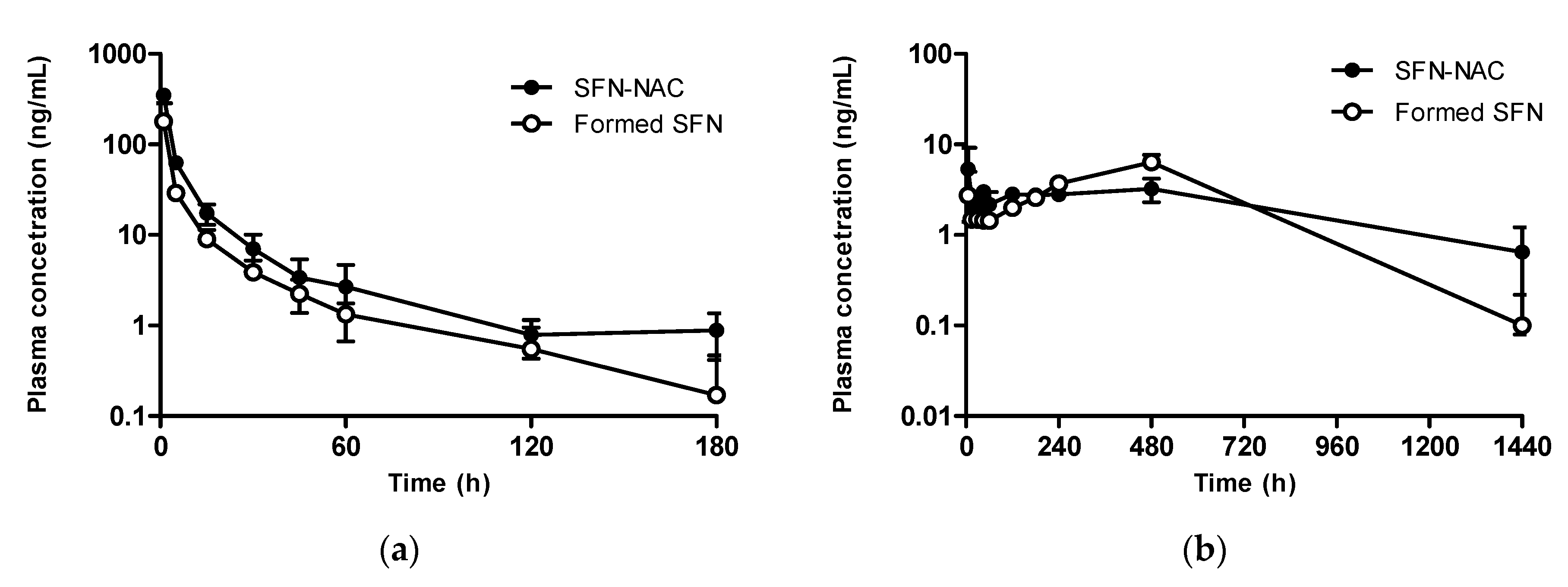
3.3. In Vivo Pharmacokinetic Study at a Micro-Dose Range
3.3.1. Sulforaphane
3.3.2. Sulforaphane N-acetylcysteine
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, H.; Zhao, C.; Tian, Y.; Lu, J.; Zhang, G.; Liang, S.; Chen, D.; Liu, X.; Kuang, W.; Zhu, M. Src family kinases and pulmonary fibrosis: A review. Biomed. Pharmacother. 2020, 127, 110183. [Google Scholar] [CrossRef]
- Liu, P.; Miao, K.; Zhang, L.; Mou, Y.; Xu, Y.; Xiong, W.; Yu, J.; Wang, Y. Curdione ameliorates bleomycin-induced pulmonary fibrosis by repressing TGF-β-induced fibroblast to myofibroblast differentiation. Respir. Res. 2020, 21, 58. [Google Scholar] [CrossRef] [Green Version]
- Phan, S.H. The myofibroblast in pulmonary fibrosis. Chest 2002, 122, 286S–289S. [Google Scholar] [CrossRef] [PubMed]
- Park, S.D.; Jung, J.H.; Lee, H.W.; Kwon, Y.M.; Chung, K.H.; Kim, M.G.; Kim, C.H. Zedoariae rhizoma and curcumin inhibits platelet-derived growth factor-induced proliferation of human hepatic myofibroblasts. Int. Immunopharmacol. 2005, 5, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Fenwick, G.R.; Heaney, R.K. Glucosinolates and their breakdown products in cruciferous crops, foods and feeding stuffs. Food Chem. 1983, 11, 249–271. [Google Scholar] [CrossRef]
- Wattenberg, L.W. Inhibition of carcinogenic effects of polycyclic hydrocarbons by benzyl isothiocyanate and related compounds. J. Natl. Cancer Inst. 1977, 58, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Hecht, S.S. Chemoprevention of isothiocyanates. J. Cell. Biochem. Suppl. 1995, 22, 195–209. [Google Scholar] [CrossRef]
- Chung, E.L. Chemoprevention of lung cancer by isothiocyanates and their conjugates in A/J mouse. Exp. Lung Res. 2001, 27, 319–330. [Google Scholar] [CrossRef]
- Conaway, C.C.; Wang, C.X.; Pittman, B.; Yang, Y.M.; Schwartz, J.E.; Tian, D.; McIntee, E.J.; Hecht, S.S.; Chung, F.L. Phenethyl isothiocyanate and sulforaphane and their N-acetylcysteine conjugates inhibit malignant progression of lung adenomas induced by tobacco carcinogens in A/J mice. Cancer Res. 2005, 65, 8548–8557. [Google Scholar] [CrossRef] [Green Version]
- Kan, S.F.; Wang, J.; Sun, G.X. Sulforaphane regulates apoptosis- and proliferation-related signaling pathways and synergizes with cisplatin to suppress human ovarian cancer. Int. J. Mol. Med. 2018, 42, 2447–2458. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.M.; Conaway, C.C.; Chiao, J.W.; Wang, C.X.; Amin, S.; Whysner, J.; Dai, W.; Reinhardt, J.; Chung, F.L. Inhibition of benzo(a)pyrene-induced lung tumorigenesis in A/J mice by dietary N-acetylcysteine conjugates of benzyl and phenethyl isothiocyanates during the post-initiation phase is associated with activation of MAP kinases and p53 activity and induction of apoptosis. Cancer Res. 2002, 62, 2–7. [Google Scholar]
- Kyung, S.Y.; Kim, D.Y.; Yoon, J.Y.; Son, E.S.; Kim, Y.J.; Park, J.W.; Jeong, S.H. Sulforaphane attenuates pulmonary fibrosis by inhibiting the epithelial-mesenchymal transition. BMC Pharmacol. Toxicol. 2018, 19, 13. [Google Scholar] [CrossRef] [Green Version]
- Conaway, C.C.; Getahun, S.M.; Liebes, L.L.; Pusateri, D.J.; Topham, D.K.; Botero-Omary, M.; Chung, F.L. Disposition of glucosinolates and sulforaphane in humans after ingestion of steamed and fresh broccoli. Nutr. Cancer 2000, 38, 168–178. [Google Scholar] [CrossRef]
- Gasper, A.V.; Al-Janobi, A.; Smith, J.A.; Bacon, J.R.; Fortun, P.; Atherton, C.; Taylor, M.A..; Hawkey, C.J.; Barrett, D.A.; Mithen, R.F. Glutathione S-transferase M1 polymorphism and metabolism of sulforaphane from standard and high-glucosinolate broccoli. Am. J. Clin. Nutr. 2005, 82, 1283–1291. [Google Scholar] [CrossRef]
- Mennicke, W.H.; Gorler, K.; Krumbiegel, G.; Lorenz, D.; Rittmann, N. Studies on the metabolism and excretion of benzyl isothiocyanate in man. Xenobiotica 1988, 82, 39–47. [Google Scholar] [CrossRef]
- Jiao, D.; Ho, C.T.; Fioles, P.; Chung, F.L. Identification and quantification of the N-acetylcysteine conjugate of allyl isothiocyanate in human urine after ingestion of mustard. Cancer Epidemiol. Biomarkers Prev. 1994, 3, 487–492. [Google Scholar]
- Tang, L.; Li, G.; Song, L.; Zhang, Y. The principal urinary metabolites of dietary isothiocyanates, N-acetylcysteine conjugates, elicit the same anti-proliferative response as their parent compounds in human bladder cancer cells. Anticancer Drugs 2006, 17, 297–305. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, G.; Tian, H.; Hu, Y.; Wu, S.; Geng, Y.; Lin, K.; Wu, W. Sulforaphane metabolites cause apoptosis via microtubule disruption in cancer. Endocr. Relat. Cancer 2018, 25, 255–268. [Google Scholar] [CrossRef]
- Mitsiogianni, M.; Koutsidis, G.; Mavroudis, N.; Trafalis, D.T.; Botaitis, S.; Franco, R.; Zoumpourlis, V.; Amery, T.; Galanis, A.; Pappa, A.; et al. The role of isothiocyanates as cancer chemo-preventive, chemo-therapeutic and anti-melanoma agents. Antioxidants 2019, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- Hu, R.; Hebbar, V.; Kim, B.R.; Chen, C.; Winnik, B.; Buckley, B.; Soteropoulos, P.; Tolias, P.; Hart, R.P.; Kong, A.N. In vivo pharmacokinetics and regulation of gene expression profiles by isothiocyanate sulforaphane in the rat. J. Pharmacol. Exp. Ther. 2004, 310, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Hanlon, N.; Coldham, N.; Gielbert, A.; Kuhnert, N.; Sauer, M.J.; King, L.J. Ioannides C Absolute bioavailability and dose-dependent pharmacokinetic behaviour of dietary doses of the chemopreventive isothiocyanate sulforaphane in rat. Br. J. Nutr. 2008, 99, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Khor, T.O.; Yang, Q.; Huang, Y.; Wu, T.Y.; Saw, C.L.; Lin, W.; Androulakis, I.P.; Kong, A.N. Pharmacokinetics and pharmacodynamics of phase II drug metabolizing/antioxidant enzymes gene response by anticancer agent sulforaphane in rat lymphocytes. Mol. Pharm. 2012, 9, 2819–2827. [Google Scholar] [CrossRef] [Green Version]
- Doan, T.N.K.; Vo, D.K.; Kim, H.; Balla, A.; Lee, Y.; Yoon, I.S.; Maeng, H.J. Differential effects of 1α,25-dihydroxyvitamin D3 on the expressions and functions of hepatic CYP and UGT enzymes and its pharmacokinetic consequences in vivo. Pharmaceutics 2020, 12, 1129. [Google Scholar] [CrossRef]
- Maeng, H.J.; Chapy, H.; Zaman, S.; Pang, K.S. Effects of 1α,25-dihydroxyvitamin D3 on transport and metabolism of adefovir dipivoxil and its metabolites in Caco-2 cells. Eur. J. Pharm. Sci. 2012, 46, 149–166. [Google Scholar] [CrossRef]
- Selman, M.; King, T.E.; Pardo, A. Idiopathic pulmonary fibrosis: Prevailing and evolving hypotheses about its pathogenesis and implications for therapy. Ann. Intern. Med. 2001, 134, 136–151. [Google Scholar] [CrossRef]
- Willis, B.C.; Liebler, J.M.; Luby-Phelps, K.; Nicholson, A.G.; Crandall, E.D.; du Bois, R.M.; Borok, Z. Induction of epithelial-mesenchymal transition in alveolar epithelial cells by transforming growth factor-beta1: Potential role in idiopathic pulmonary fibrosis. Am. J. Pathol. 2005, 166, 1321–1332. [Google Scholar] [CrossRef]
- Richeldi, L.; Collard, H.R.; Jones, M.G. Idiopathic pulmonary fibrosis. Lancet 2017, 389, 1941–1952. [Google Scholar] [CrossRef]
- Nakamura, T.; Matsushima, M.; Hayashi, Y.; Shibasaki, M.; Imaizumi, K.; Hashimoto, N.; Shimokata, K.; Hasegawa, Y.; Kawabe, T. Attenuation of transforming growth factor-β-stimulated collagen production in fibroblasts by quercetin-induced heme oxygenase-1. Am. J. Respir. Cell Mol. Biol. 2011, 44, 614–620. [Google Scholar] [CrossRef]
- Shapiro, T.A.; Fahey, J.W.; Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Stephenson, K.K.; Wade, K.L.; Ye, L.; Talalay, P. Safety, tolerance, and metabolism of broccoli sprout glucosinolates and isothiocyanates: A clinical phase I study. Nutr. Cancer 2006, 55, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Alumkal, J.J.; Slottke, R.; Schwartzman, J.; Cherala, G.; Munar, M.; Graff, J.N.; Beer, T.M.; Ryan, C.W.; Koop, D.R.; Gibbs, A.; et al. A phase II study of sulforaphane-rich broccoli sprout extracts in men with recurrent prostate cancer. Investig. New Drugs 2015, 33, 480–489. [Google Scholar] [CrossRef]
- Lozanovski, V.J.; Polychronidis, G.; Gross, W.; Gharabaghi, N.; Mehrabi, A.; Hackert, T.; Schemmer, P.; Herr, I. Broccoli sprout supplementation in patients with advanced pancreatic cancer is difficult despite positive effects-results from the POUDER pilot study. Investig. New Drugs 2020, 38, 776–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | IV | PO | ||
|---|---|---|---|---|
| 0.1 mg/kg | 0.1 mg/kg | 0.2 mg/kg | 0.5 mg/kg | |
| AUClast (μg·min/mL) | 2.35 ± 0.154 | 1.85 ± 0.377 | 3.16 ± 0.073 | 29.8 ± 8.64 |
| AUCinf (μg·min/mL) | 2.47 ± 0.205 | 1.92 ± 0.411 | 3.57 ± 0.253 | 31.0 ± 8.89 |
| t1/2 (min) | 61.3 ± 36.2 | 41.9 ± 7.60 | 197.7 ± 169 | 320 ± 75.4 |
| MRT (min) | 56.2 ± 11.7 | |||
| Cmax (μg/mL) | - | 0.036 ± 0.010 | 0.065 ± 0.012 | 0.143 ± 0.035 |
| Tmax (min) | - | 15.0 ± 0.0 | 5.0 ± 0.0 | 30.0 ± 18.4 |
| Vss (mL/kg) | 2268 ± 337 | |||
| CL (mL/min/kg) | 40.7 ± 3.4 | |||
| F (%) | 77.8% | 72.4% | 251.4% |
| Parameters | IV (SFN-NAC 0.1 mg/kg) | PO (SFN-NAC 0.5 mg/kg) | ||
|---|---|---|---|---|
| SFN-NAC | SFN | SFN-NAC | SFN | |
| AUClast (μg·min/mL) | 2.11 ± 0.502 | 1.07 ± 0.215 | 2.67 ± 1.22 | 1.73 ± 0.186 |
| AUCinf (μg·min/mL) | 2.16 ± 0.515 | 1.11 ± 0.233 | NC | NC |
| t1/2 (min) | 33.2 ± 14.2 | 48.6 ± 11.0 | NC | NC |
| MRT (min) | 15.8 ± 7.7 | |||
| Cmax (μg/mL) | - | 0.006 ± 0.003 | 0.006 ± 0.001 | |
| Tmax (min) | - | |||
| Vss (mL/kg) | 711 ± 262 | - | - | - |
| CL (mL/min/kg) | 48.3 ± 10.5 | - | - | - |
| F(%) | - | - | >24.8% | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, E.S.; Fei, X.; Yoon, J.-H.; Seo, S.-Y.; Maeng, H.-J.; Jeong, S.H.; Kim, Y.C. Comparison of Pharmacokinetics and Anti-Pulmonary Fibrosis-Related Effects of Sulforaphane and Sulforaphane N-acetylcysteine. Pharmaceutics 2021, 13, 958. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13070958
Son ES, Fei X, Yoon J-H, Seo S-Y, Maeng H-J, Jeong SH, Kim YC. Comparison of Pharmacokinetics and Anti-Pulmonary Fibrosis-Related Effects of Sulforaphane and Sulforaphane N-acetylcysteine. Pharmaceutics. 2021; 13(7):958. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13070958
Chicago/Turabian StyleSon, Eun Suk, Xiang Fei, Jin-Ha Yoon, Seung-Yong Seo, Han-Joo Maeng, Sung Hwan Jeong, and Yu Chul Kim. 2021. "Comparison of Pharmacokinetics and Anti-Pulmonary Fibrosis-Related Effects of Sulforaphane and Sulforaphane N-acetylcysteine" Pharmaceutics 13, no. 7: 958. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13070958