
A Review on the Prevalence of Poxvirus Disease in Free-Living and Captive Wild Birds


1
Departamento de Genética, Fisiología y Microbiología, Facultad de Biología, Universidad Complutense de Madrid C, José Antonio Nováis, 28040 Madrid, Spain
2
Centro de Biotecnología y Genómica de Plantas, Universidad Politécnica de Madrid-Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (CBGP, UPM-INIA), 28223 Madrid, Spain
*
Author to whom correspondence should be addressed.
Microbiol. Res. 2021, 12(2), 403-418; https://0-doi-org.brum.beds.ac.uk/10.3390/microbiolres12020028
Submission received: 2 April 2021
/
Revised: 24 April 2021
/
Accepted: 27 April 2021
/
Published: 30 April 2021
Abstract
:Avian pox is a widespread infection in birds caused by genus Avipoxvirus pathogens. It is a noteworthy, potentially lethal disease to wild and domestic hosts. It can produce two different conditions: cutaneous pox, and diphtheritic pox. Here, we carry out an exhaustive review of all cases of avian pox reported from wild birds to analyze the effect and distribution in different avian species. Avian poxvirus strains have been detected in at least 374 wild bird species, a 60% increase on a 1999 review on avian pox hosts. We also analyze epizootic cases and if this disease contributes to wild bird population declines. We frequently observe very high prevalence in wild birds in remote island groups, e.g., Hawaii, Galapagos, etc., representing a major risk for the conservation of their unique endemic avifauna. However, the difference in prevalence between islands and continents is not significant given the few available studies. Morbidity and mortality can also be very high in captive birds, due to high population densities. However, despite the importance of the disease, the current detection rate of new Avipoxvirus strains suggests that diversity is incomplete for this group, and more research is needed to clarify its real extent, particularly in wild birds.
1. Introduction
Avian pox is an infectious disease of domestic and wild birds, caused by the transmission of viruses in the genus Avipoxvirus, family Poxviridae [1]. Poxviruses are large, linear, double-stranded DNA viruses with a genome of 250–400 kb encoding 250–300 proteins. They are amongst the largest animal viruses (~250 by 350 nm) and are oval or brick-shaped [2]. Avian poxvirus (APV) has been detected in birds from all continents (including Antarctica [3]), as well as a number of remote islands. The virus can be transmitted directly between host individuals, environmentally, or mechanically by biting arthropods [4,5,6].
The ICTV (International Committee on Taxonomy of Viruses) recognizes 12 species (and has proposed two more) in the genera Avipovirus [7], based on several criteria, although the phylogenetic distance and natural host are the primary criteria used for the creation of new species and genera [8]. A molecular survey has identified 152 unique sequences [9], suggesting that APV species limits are far from resolved. Genetic analyses are dominated by studies of the 578 bp 4 b gene fragment (fpv167 locus), chosen because it is relatively conservative between Avipoxvirus strains [10]. Most genetic lineages infect multiple, taxonomically varied host species, suggesting that adaptation to specific host taxa may be limited, at least for some strains [11,12]. It remains controversial whether strains are typically taxa specific or broad spectrum, and to what extent broad spectrum infection is influenced by anthropogenic factors.
2. Host Range
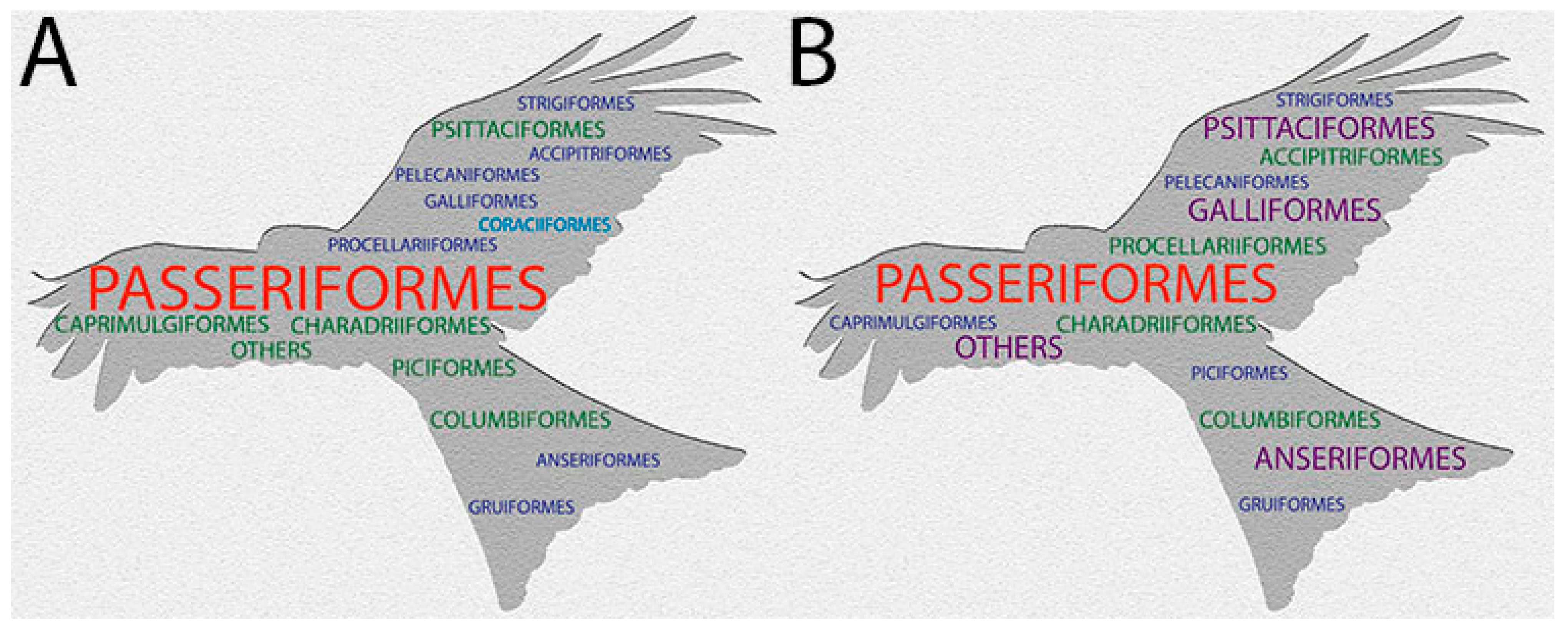
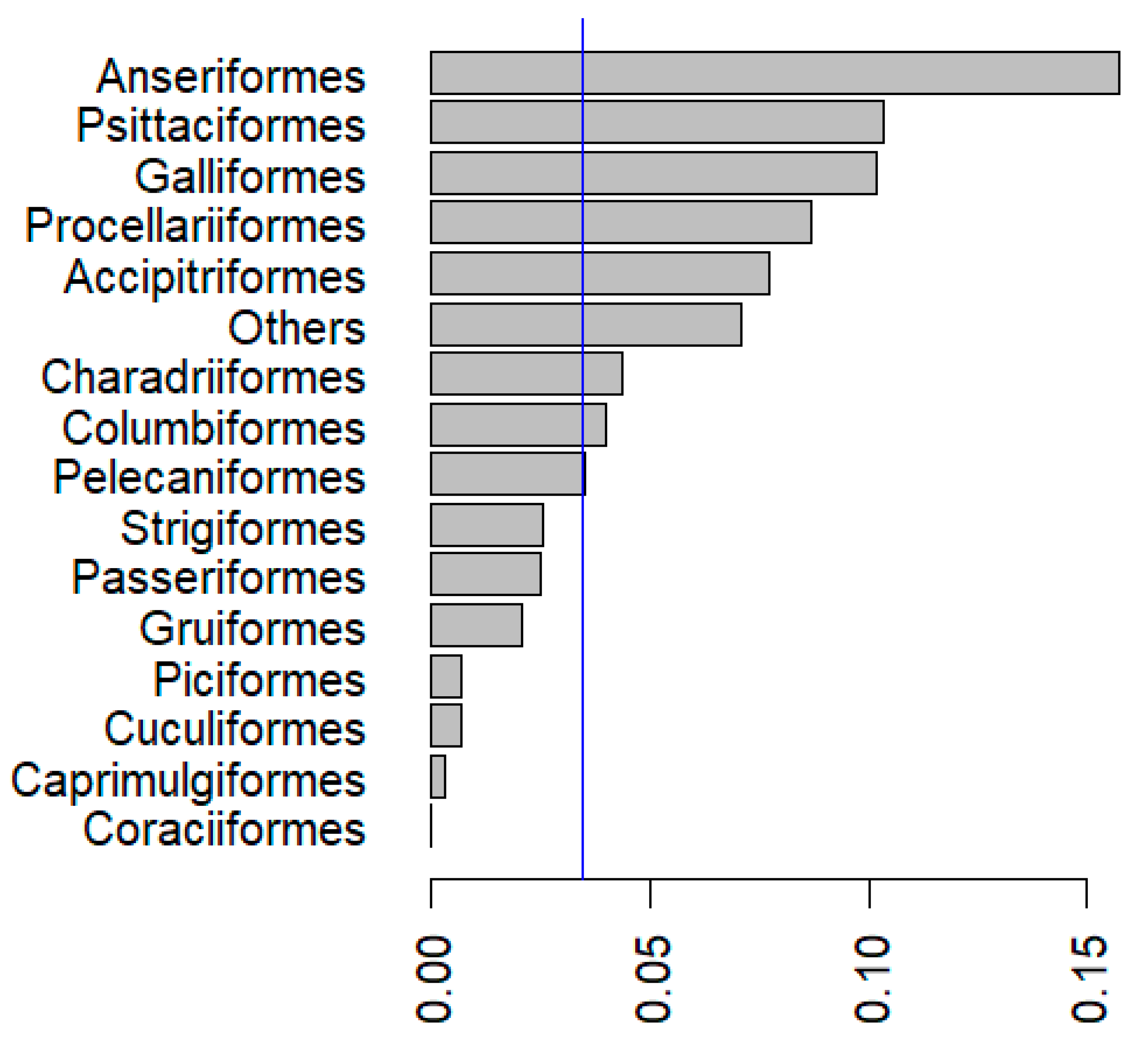
APV was reported to have been detected in 278 species of 23 orders in 1999 [13]. We report in this review, that APV has now been detected in at least 374 avian species from 23 orders (Table S1). APV infects a broad range of avian hosts: from ostriches to hummingbirds and including domestic birds, raptors, parrots, waterbirds, seabirds, etc. It has been detected in all major avian orders, except for Coraciiformes (kingfishers and allies), probably because this group contains relatively few species (1.7% of avian species), which are infrequently in contact with humans. Most APV detections (42%) have been from Passeriformes, the most speciose bird order (61% of species; Figure 1). We carried out an exact binomial test in R version 4.04 to determine whether the proportion of species infected with APV in each order (following Clements et al. [14]) were higher or lower than the expected level of 0.035% (Figure 2; Appendix A; Table S2). In total, APV was detected from more species than expected (p < 0.005) in six bird orders (including “others”, an amalgamation of several minority orders; explanation Table S2) and from less species than expected (p < 0.005) in four bird orders. Bird orders with more APV-positive species than expected include domestic species and larger species (Galliformes, Anseriformes, Psittaciformes, Accipitriformes, Procellariformes and most of “others”—Struthioniformes, Otidiformes, Ciconiiformes, etc.). Bird orders with less APV-positive species included small and cryptic species which are potentially easier to overlook (Caprimulgiformes, Passeriformes, Piciformes and Coraciiformes) and tend to be speciose, potentially biasing the results.
3. Pathology
Infection with APV can cause two common conditions. Cutaneous pox, or dry pox, which is probably the most common form, is characterized by wart-like growths on feather-free areas (legs, feet, beak or eyes) of the bird. Most mild cutaneous pox infections are small (1–5 mm in diameter) and are not debilitating [15]. However, extensive lesions can impair vision, feeding and breathing [15,16]. In recent outbreaks of pox lesions in great tits (Parus major) in the UK, many infected individuals had large growths in feathered areas [17], and it is possible that smaller pox lesions in feathered areas may be overlooked.
Poxvirus infection is not generally considered to be the direct cause of mortality of all affected birds, but the lesions that this disease produces may increase the risk of accident, predation, acquiring secondary infections, and reduce breeding and foraging success [2,18,19]. Due to these factors, some authors argue most wild birds die before reaching advanced stages of the disease [2]. However, the mortality rate associated to APV infection is similar to other infectious diseases [20].
Diphtheritic, or wet pox, causes growths on the mucous membranes of the mouth, throat, trachea and lungs. Occasionally, these lesions can be debilitating, leading to emaciation and death [15]. The diphtheritic form of the disease is observed most frequently in Psittaciformes, Galliformes, and Columbiformes [13] although it has also been reported in Struthioniformes [21] or Passeriformes [22]. Cutaneous and diphtheritic forms can also occur simultaneously in mixed pox infections [23,24]. In general, diphtheritic and mixed pox infections present higher mortality rates than the cutaneous form [13,22,25], and it seems to cause higher mortality in wild birds than in poultry [26]. Septicemic infection is a rare expression of the disease [27], reported especially in canaries [2,28].
The duration of infection is not well understood. It may vary according to APV strain, host species and individual immunology. It seems likely that mild APV infections usually last for 3–4 weeks, but that they can be considerably longer if complications arise [25]. In a deliberate infection study [29], the pox lesions in the majority of bird species disappeared in this time frame; however, lesions persisted for more than 10 weeks in 5-day old pigeon squabs deliberately infected with pigeon poxvirus.
4. Epidemiology
Infected birds are the main reservoir for APV. The most frequent transmission route is via biting insects, mostly midges (Ceratopogonidae) and mosquitoes (Culicidae) [23,30]. Other arthropods vectors have also been reported, sometimes as mechanical vectors such as fleas (Parapsyllus longicornis [31]) and blowflies (Phaenicia sp.) [16]. However, environmental infection represents an unquantified, and probably small, component of the transmission cycle. Objects contaminated with viable poxviruses, such as feeders or perches, are recorded to spread poxviruses and it appears to be spread also by the inhalation or ingestion of dust or aerosols [2]. Supplemental feeding using bird feeders might be a positive conservation measure, but also helps to spread the virus. Bird populations in forests with bird feeders show a higher prevalence of pox-like lesions than those without anthropogenic food supply [32]. Moreover, avian pox was the predominant disease among birds using feeders (81% of the diseased birds). It is likely that once contaminated objects have been removed, transmission dynamics return to their previous state: in the Wilcoxen study, APV prevalence 10 months after bird feeders had been removed were not significantly different from forests which never had anthropogenic food supplements. Another study [33] found that APV infection prevalence increased in house finches (Haemorhous mexicanus) in areas of high human population density and was inversely related to the proportion of undisturbed habitats.
The most important epizootiological factors of APV distribution include vector abundance, weather conditions, host density and host susceptibility [2,30,34].
It is commonly reported that poxvirus infection prevalence in wild birds increases after vector population peaks [34,35,36]. Poxvirus infection prevalence is lower at high altitudes, where vectors are less abundant [30,37,38].
Weather conditions may also explain higher prevalence of APV lesions. Lesions become more frequent with the seasonal increase in temperature and precipitation. In most cases, a peak in APV lesions is reported in summer and/or fall, when vectors are more abundant [35,39,40,41]. One UK study found that APV incidence throughout the calendar year in Paridae and non-Paridae peaked in the late summer/autumn (August–September) of each year [39]. Furthermore, APV prevalence is reportedly related to annual rainfall [35,37]. For example, the annual monitoring of pox virus prevalence (2003–2007) in Laysan Albatross chicks in Hawaii has shown dramatic changes between years (3% to 88%), with low prevalence in years with low annual rainfall, and few mosquitoes, and the highest known prevalence in years with high rainfall and abundant mosquitoes [42]. Rainfall amounts at least as high as pox epizootics in 1996 and 2004 have occurred 13 times since 1947, or once every 4.5 years [43].
Age and immune function are reported to be important factors explaining within-species differences in APV prevalence. In nearly all studies, avipox lesions are significantly more frequent in juveniles than in adults [40,44,45,46,47]—this is anticipated, as first-year birds will not have acquired specific immunity to APV infection. Le Loc’h et al. [41] showed that this pattern was true only for houbara bustards in Morocco but not in the United Arab Emirates, where pox infection affected juveniles and adults alike. To our knowledge, few studies [48,49] show that the prevalence of APV lesions in short-toed larks and Berthelot’s pipits in the Canary Islands and in mockingbirds in Galápagos were higher in adults than in juveniles. Similarly, the interannual variation in innate immune function correlates to temporal changes in the susceptibility to the disease [50].
5. Prevalence of Pox Lesions and APV Infections in Wild Birds
The literature on APV infection is dominated by case-reports or epizootics in specific species or groups during a set time-period or the study of defined outbreaks. Prevalence studies have only been carried out at a handful of sites around the world, and most studies focus on pathogen characterization in birds with proliferative skin lesions (pox-suspected), mainly of the legs, feet or head. The prevalence of avipoxvirus infection is often evaluated through the inspection of pox-like lesions, although in these cases the diagnosis is presumptive. Furthermore, this risks lumping active APV infection and recovered APV infections in counts of “positive” APV infections.
Studies on the frequency of pox-like lesions involving one or several species have been mainly focused on finches, sparrows and tits in the order Passeriformes. The largest survey undertaken to date (Table 1), was of approximately 54,000 Magellanic penguins (Spheniscus magellanicus) in Punta Tombo and Cabo Dos Bahías (Argentina) [31]. A 7-year study in Trinidad tested more than 9000 individuals of tropical birds [52]. There are also a few studies with sample sizes in the range of 2000–6000 individuals, which have been used to explore APV infection in falcons, quails, great tits, house sparrows, finches or mockingbirds in the UAE/Kuwait, USA, the UK, Spain and Galapagos [15,30,39,46,51,53,54].
The occurrence of avipox lesions and APV infection is highly variable (Table 1). An incidence around 0.5–1.5% has been reported as the modal avian pox prevalence of lesions on wild birds in continental regions, where avian pox and its hosts have had a long co-evolutionary history [2]. Nevertheless, some studies in continental areas reveal a wide range in the prevalence of infection, ranging from 0.9 to 12% [15,33,51,55,56]. In contrast, megadiverse tropical regions in Ecuador, French Guiana or Bermuda [54,57,58] show very low prevalences. Traditionally, pox-like lesions and avipox infection prevalences are considered to be higher in islands than in continents. We tested whether this difference was significant by analysing the prevalence of APV infection in continents and islands using generalized linear mixed models, where location was the fixed factor. Although the mean prevalence of this infection was higher in islands (11.2% vs. 2.3% in continents) this difference was not significant (t = 1.27; p = 0.21). However, as the number of studies available may not be high enough to analyse this difference with sufficient statistical power (n = 11 for continents and n = 18 for islands), future research on APV prevalence in wild bird populations will help clarify it.

{kind=link}
{kind=link}
Table 1.
Prevalence of avian poxvirus in surveys that are not identified by authors as epizootics or outbreaks.
Table 1.
Prevalence of avian poxvirus in surveys that are not identified by authors as epizootics or outbreaks.
| Order | Genera | Species | Common Name | Sample Size | Prevalence (%) | Location (Country) | Period | Reference | Range |
|---|---|---|---|---|---|---|---|---|---|
| Several | Several | 94 | Several | 889 | 0.0 | French Guiana | 2015–2016 | [59] | Continent |
| Several | Several | 3 | Doves and Gambel´s quail | 1792 | 0.0 | Arizona, USA | 1962 | [60] | Continent |
| Passeriformes | Centronyx | 1 | Henslow´s sparrow | 165 | 9.0 | Wisconsin, USA | 2005–2008 | [61] | Continent |
| Spheniscidae | Spheniscus | 1 | Magellanic Penguin | 54,000 | 0.2 | Punta Tombo and Cabo Dos Bahıas, Argentina | 1982–2010 | [31] | Continent |
| Several | Several | 132 | Several | 941 | 0.3 | Ecuador | 2015 | [57] | Continent |
| Passeriformes | Parus | 1 | Great tit | 1819 | 0.82 | Pilis Mountains, Hungary | 2006–2007 | [56] | Continent |
| Several | Several | 37 | Several | 1075 | 1.3 | Czech Republic | 2005 | [55] | Continent |
| Passeriformes | Passer | 1 | House sparrow | 2729 | 3.1 | Central/South, Spain | 2012–2014 | [46] | Continent |
| Passeriformes | Sylvia | 2 | Blackcap | 244 | 3.7 | Czech Republic | 2005 | [55] | Continent |
| Columbiformes | Streptopelia | 1 | Oriental turtle dove | 600 | 4.8 | Republic of Korea | 2011–2013 | [23] | Continent |
| Passeriformes | Bucanetes | 1 | Trumpeter finch | 228 | 0.0 | Canary Islands, Spain | 2002–2003 | [48] | Islands |
| Passeriformes | Passer | 1 | Spanish sparrow | 128 | 0.0 | Canary Islands, Spain | 2002–2003 | [48] | Islands |
| Passeriformes | Sitta | 1 | Nuthatch | 107 | 0.0 | Oxford, UK | 2009–2011 | [39] | Islands |
| Passeriformes | Cyanistes | 1 | Blue tit | 6700 | 0.06 | Oxford, UK | 2009–2011 | [39] | Islands |
| Passeriformes | Periparus | 1 | Coal tit | 1013 | 0.58 | Oxford, UK | 2009–2011 | [39] | Islands |
| Passeriformes | Poecile | 1 | Marsh tit | 579 | 0.85 | Oxford, UK | 2009–2011 | [39] | Islands |
| Passeriformes | Parus | 1 | Great tit | 3661 | 5.01 | Oxford, UK | 2009–2011 | [39] | Islands |
| Passeriformes | Aegithalos | 1 | Long-tailed tit | 42 | 0.0 | Oxford, UK | 2009–2011 | [39] | Islands |
| Phaethontiformes | Phaethon | 1 | White-tailed tropicbird | 81 | 0.3 | Castle Harbour Island Natural Reserve, Bermuda | 1958–1978 | [58] | Islands |
| Several | Several | 4 | Several | 9514 | 1,82 | Trinidad, Trinidad and Tobago | 1964–1971 | [52] | Islands |
| Passeriformes | Several | 11 | Passerines | 3122 | 13.5 | Hawaii, USA | 1977–1980 | [30] | Islands |
| Passeriformes | Hemignathus | 1 | Hawaii Amakihi | 94 | 2 | Hawaii, USA | 2002 | [62] | Islands |
| Procellariiformes | Phoebastria | 1 | Laysan albatross | 70 | 3 | Hawaii, USA | 2003/2007 | [42] | Islands |
| Passeriformes | Several | 9 | Passerines | 484 | 5 | Hawaii, USA | 1995 | [63] | Islands |
| Procellariiformes | Phoebastria | 1 | Waved albatross | 314 | 6 | Galapagos Islands, Ecuador | 2013–2014 | [24] | Islands |
| Passeriformes | Several | 17 (at least) | Darwin’s finch and Galapagos mockingbird | 3607 | 6.3 | Galapagos Islands, Ecuador | 1898–1906 | [54] | Islands |
| Passeriformes | Several | 10 | Darwin´s finch | 898 | 7 | Galapagos Islands, Ecuador | 2000–2002/2004 | [38] | Islands |
| Passeriformes | Anthus | 1 | Berthelot´s pipit | 447 | 8 | Canary Islands, Spain. Madeira and Ilhas Selvagens, Portugal | ND | [64] | Islands |
Several surveys have been carried out in house sparrows (Passer domesticus), permitting the comparison of prevalences of APV in the same species in different ecosystems and regions. Van Riper et al. [30] found prevalences of 2.5% and 4.9% of active and inactive avian pox lesions in 81 house sparrows captured in Hawaii (albeit a limited sample size) and similar prevalences of 3.1% were reported in an extensive sample size (2729) in Spain [46]. In contrast, Smits et al. [48] did not report any clinical signs of infection in Spanish sparrows (P. hispanolensis) in the Canary Islands (n = 128).
In some regions, the prevalence of APV seems to be increasing over time. Since 2005, the number of APV cases in great tit has increased markedly in Central Europe [56] and in Britain [17]. In Britain the three-fold increase in APV cases in Paridae from 2006-2010, and the expansion both north and west from the first detection in south-east England, was indicative of an emerging infection. One rigorous study [54] found 6.3% of individuals symptomatic for APV—not counting recovered infections. However, the authors caution that visual inspection for cutaneous lesions is not APV diagnosis as they were only able to lab confirm APV infection in 21/59 symptomatic samples, suggesting the true prevalence may have been closer to 2%. They found alternative aetiology for lesions in almost all negative cases using histopathology. This shows that determining APV prevalences relying only on observational data can be misleading, and confirmation in the lab should be carried out afterwards in order to obtain accurate prevalence values.
The literature also reports several epizootic events (Table 2), showing exceptionally high prevalences (up to 88%) as in the case of House and Galapagos finches, mockingbirds, Short-toed larks, Berthelot´s pipit, Hawai';i 'elepaio, crows or Layssan Albatross. The infection is highly invasive and pathogenic to naïve bird populations in island groups such as Hawaii [30], Galapagos [50,54,65] and Canary Islands [48], where they pose a threat to the unique and vulnerable endemic avifauna [24,66]. The sampling of native Hawaiian forest birds in Hawaiian Islands showed a prevalence ranging from 5% to 13.5% [30,63]. In contrast, no pox-like lesions were detected among non-native bird species in Hawaii [63]. Some examples include very high prevalences (26–29%) in Berthelot’s pipits within and between three archipelagos of Macaronesia, although the overall prevalence was 8% [48]. Similarly, in the Canary Islands, pox-lesion prevalences were 50% in lesser short-toed larks (Calandrella rufescens) and 28% of Berthelot’s pipits (Anthus berthelotii) trapped around farmyards [64]. Additionally, epizootics of poxvirus infection have been described in several game species (Order Galliformes). An outbreak in bobwhite quails (Colinus virginianus) in southwest Georgia and northcentral Florida showed an approximate 12-fold increase (1 to 39%) in the incidence of avian pox. These authors estimated a mortality rate of approximately 0.6 to 1.2% [15]. APV prevalence in Gambel’s quail (Lophortyx gambelii) in Southern Arizona was between 11–16% [60] whereas in California quails (Callipepla californica) in Oregon it reached 21–26% [67,68]. Similarly, the population of red-legged partridges (Alectoris rufa) in Spain shows a very high prevalence (around 60%) [40].
6. Contribution to Wild Population Declines
Several studies suggest that APV contributed to atypical population declines of some wild bird species. For example, since the disease was identified in Carrion and Large-billed crows (Corvus corone, C. macrorhynchos) in Japan in 2006, the mortality in these species has increased dramatically [47]. Infection is also cited as one factor in the extirpation from the wild of another Corvid, the Hawaiian Crow (C. hawaiiensis) [71]. Evidence for the role of APV in this extirpation is presumptive; nonetheless, two APV strains were characterized from captive individuals of this species in a later study (72), so this virus should be considered a threat to this endangered species. Factors that may have heightened APV incidence include the increase of arthropod vector populations in areas where they were not previously common, possibly due to the climate change, and modification of migration habits [31,72,73]. Another important factor to consider is human impact on wild bird populations associated with urban habitats, such as the previously mentioned crows. In such cases, bird density may increase the transmission of infectious diseases like avian pox by increasing the pool of susceptible hosts [74].
Novel poxvirus isolates discovered to infect wild bird populations usually produce outbreaks with high rates of mortality, decreasing the number of individuals which were naïve to these strains, especially when infection is caused by species translocation. A number of studies find that APV has been introduced to remote islands in introduced domestic or wild released birds (for example, Hawaii, Galapagos, Canary Islands), with the result that the disease spread rapidly, with prevalences much higher in the endemic avifauna than in introduced avian species, which have evolved in the presence of the pathogen [30,37,48,63,69,75]. It is difficult to confirm this claim hundreds of years after the putative introduction. However, it is noteworthy, that in some island groups, very high mortality rates have been documented in the endemic avifauna, and sometimes overlaps with other putatively introduced diseases transmitted by arthropod vectors, such as avian malaria [30,48,65,66,76]. APV prevalence has been shown to be significantly higher in native than introduced species [62,63]. It is important to understand the incidence and dynamics of APV infection, to avoid population declines in these communities and to establish effective control measures [17,39,73].
The rapid spread of the infection in islands is well documented. An APV outbreak was reported in two colonies of gentoo penguins in the Falkland Islands in early February, and just one month after the first report, 27% of one colony were reported dead from the disease [77].
In a necropsy of 137 Laysan albatross chicks (summer 1987), five carcasses (3.6%) had lesions consistent with avipox infection, though pox was a secondary finding considered unrelated to the cause of death [78]. This contribution mentions unpublished data recording epizootic pox with extensive, severe morbidity has occurred irregularly in Laysan albatross chicks at Sand Island. Young et al. [34] showed extremely high avipox incidence (up to 88% in one year) but with virtually no mortality, and attributing the two mortalities to other causes. The fledging rate on Midway Atoll in pox epizootic years (82%) did not differ from low pox prevalence years (80%) or the average rate (86%).
7. Outbreaks in Captive Wild Birds
Studies of APV outbreaks in captive birds has largely focused on poultry and domestic birds, primarily chickens and turkeys [79]. However, similar conditions occur with free-living species in commercial farms, zoos, recovery centres, aviaries and captive-breeding programmes [22,41,44,80,81,82,83,84] (Table S3). Confining birds brings together species and populations, and any parasites they might host, to form unnatural communities, often at artificially high concentrations. It thus favours the rapid dispersion of APV infection if the virus arrives in the enclosure. It is, however, easier to monitor (and simpler to detect) the infection in captive than in wild populations. Mortality rates among captive birds may reach very high levels, and in many cases their death it is due to the systemic form of the disease, for example in canaries [28,85,86,87].
APV infections may be introduced to captive populations from wild birds, or newly introduced captive birds, fomites, or by arthropod vectors. The introduction of infectious diseases to aviaries or zoos by sick house sparrows, feral pigeons (or rock doves) or common starlings has been documented [88] and is frequently suspected, especially in open aviaries like rooftop aviaries [89]. Silent carriers, infected individuals that show no clinical signs, have been documented to spread APV infection inside aviaries after a ten-day quarantine [90], which may be an important route of transmission. Suboptimal conditions of the aviary together with the abundance of bloodsucking arthropods such as mites or mosquitoes also trigger APV infections in aviaries, and dust and dander become an important way of transmission in these facilities [91].
There are several documented cases of accidental or deliberate introduction of infected hosts to distant sites. For example, the introduction of 25 stone curlews (Burhinus oedicnemus) to a captive breeding facility that already contained 140 stone curlews caused disease symptoms, but no mortality, in 100% of the original captive population [92], initially. A second APV outbreak occurred in the same farm six months later, in 10 males segregated from the rest of the population led to morbidity in 7/10 and death in 5/10 died. The outbreak was limited to this species and none of the four gallinaceous species on the farm became sick. Another case of host translocation occurred in Germany, when 21 peregrine falcons (Falco peregrinus) were released as part of a reintroduction programme, and within five years six of them had died displaying APV signs [93]. Likewise, the introduction of 25 hatch-year lovebirds (Agapornis spp.) apparently triggered a poxvirus outbreak among captive exotic parrots 7 days following their arrival [83]. Lastly, two outbreaks of mortality in South Island saddlebacks (Philesturnus carunculatus) occurred following translocation to two of New Zealand’s offshore islands. Four of 12 individuals captured had apparent APV lesions, and this was confirmed by PCR and sequencing in two of two cases. The outbreak was associated to a mortality of 60% [94], though this was concurrent with avian malaria infections, intestinal gut parasites and heavy infestation with hippoboscid flies in some individuals, so the actual cause of death was not demonstrated conclusively.
Several APV outbreaks were described from captive-breeding programmes for houbara and MacQueen’s bustards (Chlamydotis undulata and Chlamydotis macqueenii) in Morocco and United Arab Emirates from 2006 to 2013, with prevalence rates of up to 2.3%, However, the impact of this disease on the ability of birds to reproduce, in terms of courtship display frequency, and egg production, seemed low and mainly non-significant [41,95].
8. Seroprevalence Surveillance
Prevalence studies give a useful snapshot of how the prevalence of incomplete syndromic or, perhaps individuals which have detectable APV infections, However, as lesions appear 2–3 weeks after virus transmission and lesions may remain only for a short period, and possibly some birds never display visible lesions: thus, the recorded prevalence might be lower than the actual prevalence. The true incidence of the viral infection, that is, the number of birds that both develop lesions and become infected, is unknown. Data on APV seroprevalence, together with data on the prevalence of syndromic or positive individuals, can help demonstrate if the disease is widely distributed among wild birds, even when prevalence is very low.
Nevertheless, there are many criticisms of estimating disease prevalence based on serology. The duration of the immune response is unknown, and if the length of the immune response is short, seronegative birds may be individuals that have seroconverted (even though they have been infected at some point of their life). Additionally, pathogens cause mortality in some individuals thus eliminating them from the population, and reducing the apparent prevalence of seropositive birds. Furthermore, it is most unlikely that the serological screening techniques that have been developed for detecting APV strains are capable of detecting the complete diversity of species, genotypes or lineages. Most serological testing for antibodies to APV use antigen specific for fowl poxvirus strains, most associated with infection in chickens. Several studies [48,96] used a mixed antigen (FPV/Pigeon poxvirus/Turkey Poxvirus) or (FPV/Pigeon poxvirus) but excluding antigen from the more genetically different canary poxvirus, that most frequently infects wild birds.
There are very few seroprevalence studies in wild bird populations. One limited size-sample study in New Zealand in introduced passerine birds challenged with fowlpox antigen found unexpectedly high seroprevalence (mean 69.2%) [97]. Surprisingly, no birds with lesions were observed during sampling, suggesting that an effective immune response leads to the low frequency of clinical lesions. These birds were also seropositive to Plasmodium spp., suggesting infection by a common vector, but again, there were no clinical signs of disease. Authors hypothesized that the passerines involved in the study acted as a reservoir for both pathogens [97].
On the other hand, other APV serosurveys have been less rewarding. A study of psittacine sera samples collected from birds showing clinical signs of APV infection detected antibodies in a small fraction of subjects (18.9%) [83]. A serosurvey study for antibodies, that cross react to FPV and pigeonpox, in short-toed larks and Berthelot’s pipits in the Canary Islands found a very low seroprevalence (2.3% seropositive compared to 40% of birds symptomatic for pox infection). However, the Canary Island strains were highly divergent from known APV strains, so this might show rather that there is little cross-reactivity between the strains infecting two passerine species from the Canary Islands and the FPV and pigeonpox virus-based agar gel immunodiffusion kits [48].
9. Conclusions
APV infections are increasingly identified by molecular sequencing. Several full APV genomes have now been published. This has revealed a great diversity of unique APV sequences, far higher than the number of APV species recognised by the ICTV. It has also shown that many, but not all, APV strains can infect multiple host species. A future challenge in the study of APV will be to obtain more sequence information, especially of full genomes, to shed light on strain diversity, taxonomic relations and strain-host limits.
In 1999, APV had been detected in 232 bird species. This has now increased to 374 species, still from 23 orders, an increase of about 60%. This list will probably increase due to the renewed focus on wild animal pathogens, combined with the increased use of more sensitive molecular surveillance methods. Detection of APV is not evenly spread through Class Aves: some bird orders contain significantly more APV positive species than expected, and others have significantly less. Future studies should explore whether this is a genuine pattern, or an artefact of sampling bias.
Wild bird surveys reveal highly variable prevalence levels. The background level appears to be fairly low, but is punctuated by epizootics with very high prevalence. Epizootics have been documented in remote island groups, in captive birds (in farms, aviaries, captive release programmes, etc), but also in wild birds in a continental setting. The epizootics raise questions about whether they are occurring more frequently, the implications of APV infection in endangered bird populations, and the risk of APV to the success of reintroduction programmes.
Serology has been little used for the study of APV in wild birds. Developing sensitive serological methods, capable of distinguishing between a broad range of APV strains could potentially expand our understanding of the frequency of infection, the proportion of individuals that suffer clinical infections, and reveal how APV maintains infection cycles between epizootics.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/microbiolres12020028/s1, Table S1: Prevalence of avian poxvirus in surveys that are not identified by authors as epizootics or outbreaks, Table S2: The spectrum of APV host diversity, Table S3: Prevalence of avian poxvirus outbreaks in captive birds.
Author Contributions
Conceptualization, R.A.J.W., D.A.T., and L.B.; methodology, R.A.J.W., D.A.T., and L.B.; validation, R.A.J.W., D.A.T., and L.B.; formal analysis, D.A.T., R.A.J.W.; investigation, R.A.J.W., D.A.T., and L.B.; resources, L.B.; data curation, R.A.J.W., D.A.T., and L.B.; writing—original draft preparation, R.A.J.W., D.A.T., and L.B.; writing—review and editing, R.A.J.W., D.A.T., and L.B.; visualization, R.A.J.W., D.A.T., and L.B.; supervision, L.B.; project administration, L.B.; funding acquisition, L.B. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Spanish National Research Agency (projects CGL2013-41642-P/BOS and CGL2017-82117-P, co-funded by the European Regional Development Fund). D.D.A.T was supported by a PhD fellowship funded by UCM (CT27/16-CT28/16). R.A.J.W. was supported by College REF Investment Fund, University of Derby, UK during part of the writing of this manuscript.
Data Availability Statement
Data used in this contribution are available in Supplementary Material.
Acknowledgments
Many thanks to Mark Bulling, University of Derby, for excellent statistical advice, assistance and R coding. Thanks also to Izaskun Fernández for assistance with bibliographic search.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
We compared whether the proportion of species infected with APV in each order (following Clements et al. 2019) were higher or lower than expected. We first calculated a single value of the number of species infected by APV across all orders by dividing the sum APV species by the sum of species in all orders = 374/10,721 = 0.035. This was compared to the number of APV positive species within each order (the actual proportions: AVP number (in that order)/no. species (in that order). We thus treated each species within each order as a Bernoulli experiment, i.e., asking if the actual proportion of APV to number of species in that order is different to the expected proportion. That is, use an exact binomial test. All analyses were carried out in R version 4.04 for windows. The code is available on request.
References
- Beukema, E.L.; Brown, M.P.; Hayball, J.D. The potential role of fowlpox virus in rational vaccine design. Expert Rev. Vaccines 2006, 5, 565–577. [Google Scholar] [CrossRef]
- van Riper, C.; Forrester, D. Avian Pox. In Infectious Diseases of Wild Birds; Thomas, N.J., Hunter, D.B., Eds.; Blackwell Publishing: Ames, IA, USA, 2007; pp. 131–176. [Google Scholar]
- Shearn-Bochsler, V.; Green, D.E.; Converse, K.A.; Docherty, D.E.; Thiel, T.; Geisz, H.N.; Fraser, W.R.; Patterson-Fraser, D.L. Cutaneous and diphtheritic avian poxvirus infection in a nestling Southern Giant Petrel (Macronectes giganteus) from Antarctica. Polar Biol. 2008, 31, 569–573. [Google Scholar] [CrossRef]
- Yeo, G.; Wang, Y.; Chong, S.M.; Humaidi, M.; Lim, X.F.; Mailepessov, D.; Chan, S.; How, C.B.; Lin, Y.N.; Huangfu, T.; et al. Characterization of Fowlpox virus in chickens and bird-biting mosquitoes: A molecular approach to investigating avipoxvirus transmission. J. Gen. Virol. 2019, 100, 838–850. [Google Scholar] [CrossRef] [PubMed]
- Kligler, I.J.; Aschner, M. Demonstration of presence of fowl pox virus in wild caught mosquitoes (Culex Pipiens). Proc. Soc. Exp. Biol. Med. 1931, 28, 463–465. [Google Scholar] [CrossRef]
- Thi, C.; Huong, T.; Murano, T.; Uno, Y.; Usui, T.; Yamaguchi, T. Avian pathology molecular detection of avian pathogens in poultry red mite (Dermanyssus gallinae) collected in chicken farms. J. Vet. Med. Sci. 2014, 14, 0253. [Google Scholar] [CrossRef]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; et al. Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses (2020). Arch. Virol. 2020, 165, 2737–2748. [Google Scholar] [CrossRef]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2016). Arch. Virol. 2016, 161, 2921–2949. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, A.M.; Gibson, D.J.; Barta, J.R.; Poulson, R.; Brown, J.D.; Allison, A.B.; Nemeth, N.M. Bayesian phylogenetic analysis of avipoxviruses from North American wild birds demonstrates new insights into host specificity and interspecies transmission. Avian Dis. 2019, 63, 427–432. [Google Scholar] [CrossRef]
- Lüschow, D.; Hoffmann, T.; Hafez, H.M. Differentiation of avian poxvirus strains on the basis of nucleotide sequences of 4b gene fragment. Avian Dis. 2004, 48, 453–462. [Google Scholar] [CrossRef]
- Gyuranecz, M.; Foster, J.T.; Dan, A.; Ip, H.S.; Egstad, K.F.; Parker, P.G.; Higashiguchi, J.M.; Skinner, M.A.; Hofle, U.; Kreizinger, Z.; et al. Worldwide phylogenetic relationship of avian poxviruses. J. Virol. 2013, 87, 4938–4951. [Google Scholar] [CrossRef] [Green Version]
- Jarmin, S.; Manvell, R.; Gough, R.E.; Laidlaw, S.M.; Skinner, M.A. Avipoxvirus phylogenetics: Identification of a PCR length polymorphism that discriminates between the two major clades. J. Gen. Virol. 2006. [Google Scholar] [CrossRef] [PubMed]
- Bolte, A.L.; Meurer, J.; Kaleta, E.F. Avian host spectrum of avipoxviruses. Avian Pathol. 1999, 28, 415–432. [Google Scholar] [CrossRef] [Green Version]
- Clements, J.F.; Schulenberg, T.S.; Iliff, M.J.; Billerman, S.M.; Fredericks, T.A.; Sullivan, B.L.; Wood, C.L. The eBird/Clements Checklist of Birds of the World: v2019. Available online: https://www.birds.cornell.edu/clementschecklist/download/ (accessed on 25 March 2021).
- Davidson, W.R.; Kellogg, F.E. An epornitic of avian pox in wild bobwhite quail. J. Wildl. Dis. 1980, 16, 293–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Docherty, D.E.; Long, R.I.; Flickinger, E.L.; Locke, L.N. Isolation of poxvirus from debilitating cutaneous lesions on four immature grackles (Quiscalus sp.). Avian Dis. 1991, 244–247. [Google Scholar] [CrossRef] [Green Version]
- Lachish, S.; Bonsall, M.B.; Lawson, B.; Cunningham, A.A.; Sheldon, B.C. Individual and population-level impacts of an emerging poxvirus disease in a wild population of great tits. PLoS ONE 2012, 7, e48545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, S.E.; Lowenstine, L.J.; Ardans, A.A. Avian pox in blue-fronted Amazon parrots. J. Am. Vet. Med. Assoc. 1981, 179, 1218–1222. [Google Scholar]
- Medina, F.M.; Ramírez, G.A.; Hernández, A. Avian pox in white-tailed laurel-pigeons from the Canary Islands. J. Wildl. Dis. 2004, 40, 351–355. [Google Scholar] [CrossRef] [Green Version]
- Fanke, J.; Wibbelt, G.; Krone, O. Mortality factors and diseases in free-ranging Eurasian cranes (Grus grus) in Germany. J. Wildl. Dis. 2011, 47, 627–637. [Google Scholar] [CrossRef]
- Perelman, B.; Gur-Lavie, A.; Samberg, Y. Pox in ostriches. Avian Pathol. 1988, 17, 735–739. [Google Scholar] [CrossRef]
- Donnelly, T.M.; Crane, L.A. An epornitic of avian pox in a research aviary. Avian Dis. 1984, 28, 517–525. [Google Scholar] [CrossRef]
- Lee, H.R.; Koo, B.-S.; Kim, J.-T.; Kim, H.-C.; Kim, M.-S.; Klein, T.A.; Shin, M.-S.; Lee, S.; Jeon, E.-O.; Min, K.-C.; et al. Molecular epidemiology of avian poxvirus in the oriental turtle dove (Streptopelia orientalis) and the biting midge (Culicoides arakawae) in the Republic of Korea. J. Wildl. Dis. 2017, 53, 749–760. [Google Scholar] [CrossRef]
- Tompkins, E.M.; Anderson, D.J.; Pabilonia, K.L.; Huyvaert, K.P. Avian pox discovered in the critically endangered waved albatross (Phoebastria irrorata) from the Galápagos Islands, Ecuador. J. Wildl. Dis. 2017, 53, 891–895. [Google Scholar] [CrossRef]
- Tripathy, D.N.; Reed, W.M. Pox. In Diseases of Poultry; John Wiley & Sons: Halbrook, NJ, USA, 2020; pp. 364–381. [Google Scholar] [CrossRef]
- Weli, S.C.; Tryland, M. Avipoxviruses: Infection biology and their use as vaccine vectors. Virol. J. 2011, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Paulo, S.; Catroxo, M.M. Identification of poxvirus under transmission electron microscopy during outbreak period in wild birds, in São Paulo, Brazil. Int. J. Morphol 2009, 27, 577–585. [Google Scholar]
- Reza, K.; Nasrin, A.; Mahmoud, S. Clinical and pathological findings of concurrent poxvirus lesions and aspergillosis infection in canaries. Asian Pac. J. Trop. Biomed. 2013, 3, 182–185. [Google Scholar] [CrossRef] [Green Version]
- Kirmse, P. Host specificity and pathogenicity of pox viruses from wild birds. Bull. Wildl. Dis. Assoc. 1969, 5, 376–386. [Google Scholar] [CrossRef]
- van Riper, C., III; van Riper, S.G.; Hansen, W.R. Epizootiology and effect of avian pox on Hawaiian forest birds. Auk 2002, 119, 929–942. [Google Scholar] [CrossRef]
- Kane, O.J.; Uhart, M.M.; Rago, V.; Pereda, A.J.; Smith, J.R.; Van Buren, A.; Clark, J.A.; Boersma, P.D. Avian pox in Magellanic Penguins (Spheniscus magellanicus). J. Wildl. Dis. 2012, 48, 790–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilcoxen, T.E.; Horn, D.J.; Hogan, B.M.; Hubble, C.N.; Huber, S.J.; Flamm, J.; Knott, M.; Lundstrom, L.; Salik, F.; Wassenhove, S.J.; et al. Effects of bird-feeding activities on the health of wild birds. Conserv. Physiol. 2015, 3, cov058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraudeau, M.; Mousel, M.; Earl, S.; McGraw, K. Parasites in the city: Degree of urbanization predicts poxvirus and coccidian infections in house finches (Haemorhous mexicanus). PLoS ONE 2014, 9, e86747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akey, B.L.; Nayar, J.K.; Forrester, D.J. Avian pox in Florida wild turkeys: Culex nigripalpus and Wyeomyia vanduzeei as experimental vectors. J. Wildl. Dis. 1981, 17, 597–599. [Google Scholar] [CrossRef]
- Forrester, D.J. The ecology and epizootiology of avian pox and malaria in wild turkeys. Bull. Soc. Vector Ecol. 1991, 16, 127–148. [Google Scholar]
- Samuel, M.D.; Woodworth, B.L.; Atkinson, C.T.; Hart, P.J.; LaPointe, D.A. The epidemiology of avian pox and interaction with avian malaria in Hawaiian forest birds. Ecol. Monogr. 2018, 88, 621–637. [Google Scholar] [CrossRef]
- VanderWerf, E.A. Distribution and potential impacts of avian poxlike lesions in ‘elepaio at Hakalau Forest National Wildlife Refuge. Stud. Avian Biol. 2001, 22, 247–253. [Google Scholar]
- Kleindorfer, S.; Dudaniec, R.Y. Increasing prevalence of avian poxvirus in Darwin’s finches and its effect on male pairing success. J. Avian Biol. 2006, 37, 69–76. [Google Scholar] [CrossRef]
- Lachish, S.; Lawson, B.; Cunningham, A.A.; Sheldon, B.C. Epidemiology of the emergent disease Paridae pox in an intensively studied wild bird population. PLoS ONE 2012, 7, e38316. [Google Scholar] [CrossRef] [Green Version]
- Buenestado, F.; Gortázar, C.; Millán, J.; Höfle, U.; Villafuerte, R. Descriptive study of an avian pox outbreak in wild red-legged partridges (Alectoris rufa) in Spain. Epidemiol. Infect. 2004, 132, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Le Loc’h, G.; Paul, M.C.; Camus-Bouclainville, C.; Bertagnoli, S. Outbreaks of pox disease due to canarypox-like and fowlpox-like viruses in large-scale houbara bustard captive-breeding programmes, in Morocco and the United Arab Emirates. Transbound. Emerg. Dis. 2016, 63, e187–e196. [Google Scholar] [CrossRef] [PubMed]
- Young, L.C.; VanderWerf, E.A. Prevalence of avian pox virus and effect on the fledging success of Laysan Albatross. J. Field Ornithol. 2008, 79, 93–98. [Google Scholar] [CrossRef]
- VanderWerf, E.A.; Burt, M.D.; Rohrer, J.L.; Mosher, S.M. Distribution and prevalence of mosquito-borne diseases in O’ahu’elepaio. Condor 2006, 108, 770–777. [Google Scholar] [CrossRef]
- Gortázar, C.; Millán, J.; Höfle, U.; Buenestado, F.J.; Villafuerte, R.; Kaleta, E.F. Pathology of avian pox in wild red-legged partridges (Alectoris rufa) in Spain. Ann. N. Y. Acad. Sci. 2002, 969, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.A. Differential prevalence of avian pox in adult and immature California quail. J. Wildl. Dis. 1986, 22, 564–566. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Martínez, J.; Ferraguti, M.; Figuerola, J.; Martínez-De La Puente, J.; Williams, R.A.J.; Herrera-Dueñas, A.; Aguirre, J.I.; Soriguer, R.; Escudero, C.; Moens, M.A.J.; et al. Prevalence and genetic diversity of avipoxvirus in house sparrows in Spain. PLoS ONE 2016, 11, e0168690. [Google Scholar] [CrossRef] [Green Version]
- Fukui, D.; Nakamura, M.; Yamaguchi, T.; Takenaka, M.; Murakami, M.; Yanai, T.; Fukushi, H.; Yanagida, K.; Bando, G.; Matsuno, K.; et al. An epizootic of emerging novel avian pox in carrion crows (Corvus corone) and large-billed crows (Corvus macrorhynchos) in Japan. J. Wildl. Dis. 2016, 52, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Smits, J.E.; Tella, J.L.; Carrete, M.; Serrano, D.; López, G. An epizootic of avian pox in endemic short-toed larks (Calandrella rufescens) and Berthelot’s pipits (Anthus berthelotti) in the Canary Islands, Spain. Vet. Pathol. 2005, 42, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curry, R.L.; Grant, P.R. Demography of the cooperatively breeding Galapagos mockingbird, Nesomimus parvulus, in a climatically variable environment. J. Anim. Ecol. 1989, 58, 441–463. [Google Scholar] [CrossRef]
- Zylberberg, M.; Lee, K.A.; Klasing, K.C.; Wikelski, M. Increasing avian pox prevalence varies by species, and with immune function, in Galápagos finches. Biol. Conserv. 2012, 153, 72–79. [Google Scholar] [CrossRef]
- McClure, H.E. Epizootic lesions of house finches in Ventura county, California. J. Field Ornithol. 1989, 60, 421–430. [Google Scholar]
- Tikasingh, E.S.; Worth, C.B.; Spence, L.; Aitken, T.H. Avian pox in birds from Trinidad. J. Wildl. Dis. 1982, 18, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Tarello, W. Prevalence and clinical signs of avipoxvirus infection in falcons from the Middle East. Vet. Dermatol. 2008, 19, 101–104. [Google Scholar] [CrossRef]
- Parker, P.G.; Buckles, E.L.; Farrington, H.; Petren, K.; Whiteman, N.K.; Ricklefs, R.E.; Bollmer, J.L.; Jiménez-Uzcá, G. 110 years of avipoxvirus in the Galapagos Islands. PLoS ONE 2011, 6, e15989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulich, P.; Roubalová, E.; Dubská, L.; Sychra, O.; Šmíd, B.; Literák, I. Avipoxvirus in blackcaps (Sylvia atricapilla). Avian Pathol. 2008, 37, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palade, E.; Biró, N.; Dobos-Kovács, M.; Demeter, Z.; Mándoki, M.; Rusvai, M. Poxvirus infection in Hungarian great tits (Parus major): Case report. Acta Vet. Hung. 2008, 56, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Moens, M.A.J.; Pérez-Tris, J.; Milá, B.; Benítez, L. The biological background of a recurrently emerging infectious disease: Prevalence, diversity and host specificity of Avipoxvirus in wild Neotropical birds. J. Avian Biol. 2017, 48, 1041–1046. [Google Scholar] [CrossRef] [Green Version]
- Wingate, D.B.; Barker, I.K. Poxvirus infection of the white-tailed tropicbird (Phaethon lepturus) in Bermuda. J. Wildl. Dis. 1980, 16, 619–622. [Google Scholar] [CrossRef] [Green Version]
- Moens, M.A.J.; Pérez-Tris, J.; Cortey, M.; Benítez, L. Identification of two novel CRESS DNA viruses associated with an Avipoxvirus lesion of a blue-and-gray Tanager (Thraupis episcopus). Infect. Genet. Evol. 2018, 60, 89–96. [Google Scholar] [CrossRef]
- Blankenship, L.H.; Reed, R.E.; Irby, H.D. Pox in mourning doves and Gambel’s quail in Southern Arizona. J. Wildl. Manag. 1966, 30, 253–257. [Google Scholar] [CrossRef]
- Ellison, K.S.; Hofmeister, E.K.; Ribic, C.A.; Sample, D.W. Relatively high prevalence of pox-like lesions in Henslow’s sparrow among nine species of migratory grassland passerines in Wisconsin, USA. J. Wildl. Dis. 2014, 50, 810–816. [Google Scholar] [CrossRef]
- Aruch, S.; Atkinson, C.T.; Savage, A.F.; LaPointe, D.A. Prevalence and distribution of pox-like lesions, avian malaria, and mosquito vectors in Kīpahulu valley, Haleakalā National Park, Hawai’i, USA. J. Wildl. Dis. 2007, 43, 567–575. [Google Scholar] [CrossRef]
- Atkinson, C.T.; Lease, J.K.; Dusek, R.J.; Samuel, M.D. Prevalence of pox-like lesions and malaria in forest bird communities on leeward Mauna Loa Volcano, Hawaii. Condor 2005, 107, 537–546. [Google Scholar] [CrossRef]
- Illera, J.C.; Emerson, B.C.; Richardson, D.S. Genetic characterization, distribution and prevalence of avian pox and avian malaria in the Berthelot’s pipit (Anthus berthelotii) in Macaronesia. Parasitol. Res. 2008, 103, 1435–1443. [Google Scholar] [CrossRef]
- Thiel, T.; Whiteman, N.K.; Tirapé, A.; Baquero, M.I.; Cedeño, V.; Walsh, T.; Uzcátegui, G.J.; Parker, P.G. Characterization of canarypox-like viruses infecting endemic birds in the Galápagos Islands. J. Wildl. Dis. 2005, 41, 342–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wikelski, M.; Foufopoulos, J.; Vargas, H.; Snell, H. Galápagos birds and diseases: Invasive pathogens as threats for island species. Ecol. Soc. 2004, 9, 5. [Google Scholar] [CrossRef]
- Crawford, J.A.; Oates, R.M.; Helfer, D.H. Avian pox in California quail from Oregon. J. Wildl. Dis. 1979, 15, 447–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, J.A.; Kilbride, K.M. Prevalence of Poxvirus in a Population of California Quail from Oregon, 1975-1987. J. Wildl. Dis. 1988, 24, 718–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, H. Frequency and effect of pox-like lesions in Galapagos mockingbirds. J. Field Ornithol. 1987, 58, 101–102. [Google Scholar]
- Zylberberg, M.; Lee, K.A.; Klasing, K.C.; Wikelski, M. Variation with land use of immune function and prevalence of avian pox in Galapagos finches. Conserv. Biol. 2013, 27, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, C.D.; Temple, S.A.; van Riper, C., III; Hansen, W.R. Disease-related aspects of conserving the endangered Hawaiian crow. In Disease and Management of Threatened Bird Populations; Cooper, J.E., Ed.; ICBP Technical Publication Series No. 10; International Council for Bird Preservation: Cambridge, UK, 1989; pp. 77–87. [Google Scholar]
- Fuller, T.; Bensch, S.; Müller, I.; Novembre, J.; Pérez-Tris, J.; Ricklefs, R.E.; Smith, T.B.; Waldenström, J. The ecology of emerging infectious diseases in migratory birds: An assessment of the role of climate change and priorities for future research. Ecohealth 2012, 9, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Lawson, B.; Lachish, S.; Colvile, K.M.; Durrant, C.; Peck, K.M.; Toms, M.P.; Sheldon, B.C.; Cunningham, A.A. Emergence of a novel avian pox disease in British tit species. PLoS ONE 2012, 7, e40176. [Google Scholar] [CrossRef] [Green Version]
- Becker, D.J.; Streicker, D.G.; Altizer, S. Linking anthropogenic resources to wildlife-pathogen dynamics: A review and meta-analysis. Ecol. Lett. 2015, 18, 483–495. [Google Scholar] [CrossRef]
- Warner, R.E. The role of introduced diseases in the extinction of the endemic Hawaiian avifauna. Condor 1968, 70, 101–120. [Google Scholar] [CrossRef]
- Tripathy, D.N.; Schnitzlein, W.M.; Morris, P.J.; Janssen, D.L.; Zuba, J.K.; Massey, G.; Atkinson, C.T. Characterization of poxviruses from forest birds in Hawaii. J. Wildl. Dis. 2000, 36, 225–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, G. Outbreak of Avian Pox Virus in Gentoo Penguins in the Falklands; CABI: Oxon, UK, 2006. [Google Scholar]
- Sileo, L.; Sievert, P.R.; Samuel, M.D. Causes of mortality of albatross chicks at Midway Atoll. J. Wildl. Dis. 1990, 26, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Giotis, E.S.; Skinner, M.A. Spotlight on avian pathology: Fowlpox virus. Avian Pathol. 2019, 48, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Al Falluji, M.M.; Tantawi, H.H.; Al-bana, A.; Sheikhly, S. Pox infection among captive peacocks. J. Wildl. Dis. 1979, 15, 597–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, C.; Maegdefrau-Pollan, B.; Bilic, I.; Liebhart, D.; Richter, S.; Mitsch, P.; Hess, M. Outbreak of cutaneous form of poxvirus on a commercial turkey farm caused by the species fowlpox. Avian Dis. 2011, 55, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Samour, J.H.; Kaaden, O.R.; Wernery, U.; Bailey, T.A. An epornitic of avian pox in Houbara Bustards (Chlamydotis undulata macqueenii). J. Vet. Med. Ser. B 1996, 43, 287–292. [Google Scholar] [CrossRef]
- González-Hein, G.; González, C.; Hidalgo, H. Case report: An avian pox outbreak in captive psittacine birds in Chile. J. Exot. Pet Med. 2008, 17, 210–215. [Google Scholar] [CrossRef]
- Esteves, F.C.; Marín, S.Y.; Resende, M.; Silva, A.S.; Coelho, H.L.; Barbosa, M.B.; D’Aparecida, N.S.; de Resende, J.S.; Torres, A.C.D.; Martins, N.R. Avian pox in native captive psittacines, Brazil, 2015. Emerg. Infect. Dis. 2017, 23, 154–156. [Google Scholar] [CrossRef] [Green Version]
- Johnson, B.J.; Castro, A.E. Canary pox causing high mortality in an aviary. J. Am. Vet. Med. Assoc. 1986, 189, 1345–1347. [Google Scholar]
- Shivaprasad, H.L.; Kim, T.; Tripathy, D.; Woolcock, P.R.; Uzal, F. Unusual pathology of canary poxvirus infection associated with high mortality in young and adult breeder canaries (Serinus canaria). Avian Pathol. 2009, 38, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catania, S.; Carnaccini, S.; Mainenti, M.; Moronato, M.L.; Gobbo, F.; Calogero, T. Isolation of avipoxvirus from tongue of canaries (Serinus canaria) show severe localized proliferative glossitis. Avian Dis. 2017, 61, 531–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landolt, M.; Kocan, R.M. Transmission of avian pox from starlings to Rothchild’s mynahs. J. Wildl. Dis. 1976, 12, 353–356. [Google Scholar] [CrossRef]
- Hukkanen, R.R.; Richardson, M.; Wingfield, J.C.; Treuting, P.; Brabb, T. Avipox sp. in a colony of gray-crowned rosy finches (Leucosticte tephrocotis). Comp. Med. 2003, 53, 548–552. [Google Scholar] [PubMed]
- Giddens, W.E.; Swango, L.J.; Henderson, J.D.; Lewis, R.A.; Farner, D.S.; Carlos, A.; Dolowy, W.C. Canary pox in sparrows and canaries (Fringillidae) and in weavers (Ploceidae); pathology and host specificity of the virus. Vet. Pathol. 1971, 8, 260–280. [Google Scholar] [CrossRef] [Green Version]
- Tsai, S.S.; Chang, T.C.; Yang, S.F.; Chi, Y.C.; Cher, R.S.; Chien, M.S.; Itakura, C. Unusual lesions associated with avian poxvirus infection in rosy-faced lovebirds (Agapornis roseicollis). Avian Pathol. 1997, 26, 75–82. [Google Scholar] [CrossRef]
- Lierz, M.; Bergmann, V.; Isa, G.; Czerny, C.P.; Lueschow, D.; Mwanzia, J.; Prusas, C.; Hafez, H.H. Avipoxvirus infection in a collection of captive stone curlews (Burhinus oedicnemus). J. Avian Med. Surg. 2007, 21, 50–56. [Google Scholar] [CrossRef]
- Krone, O.; Essbauer, S.; Wibbelt, G.; Isa, G.; Rudloph, M.; Gough, R.E. Avipoxvirus infection in peregrine falcons (Falco peregrinus) from a reintroduction programme in Germany. Vet. Rec. 2004, 154, 110–113. [Google Scholar] [CrossRef]
- Alley, M.R.; Ha, H.J.; Howe, L.; Hale, K.A.; Cash, W. Concurrent avian malaria and avipox virus infection in translocated South Island saddlebacks (Philesturnus carunculatus carunculatus). N. Z. Vet. J. 2010, 58, 218–223. [Google Scholar] [CrossRef]
- Le Loc’h, G.; Souley, M.-N.A.; Bertagnoli, S.; Paul, M.C. Low impact of avian pox on captive-bred houbara bustard breeding performance. Front. Vet. Sci. 2017, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Tadese, T.; Potter, E.A.; Reed, W.M. Development of a mixed antigen agar gel enzyme assay (AGEA) for the detection of antibodies to poxvirus in chicken and turkey sera. J. Vet. Med. Sci. 2003, 65, 255–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, H.J.; Banda, M.; Alley, M.R.; Howe, L.; Gartrell, B.D. The seroprevalence of avipoxvirus and its association with avian malaria (Plasmodium spp.) infection in introduced passerine birds in the southern regions of the North Island of New Zealand. Avian Dis. 2013, 57, 109–115. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
This word cloud (Wordart.com; accessed 25 March 2021) represents: (A) the proportion of the 10,721 avian species in 41 avian [14] orders. For brevity, 24 orders constituting less than 1% of avian diversity are combined into “others”; (B) the number of species detected with APV in each order. Orders are color-coded to represent the percentage of avian species found/cases of APV detected in each order, where blue and small represents <3%, green 3–7.5%, purple 7.5–11%, and red and large >11%. Although APV has been detected in more Passeriformes species than non-Passeriformes species, more APV are detected in non-Passeriformes than in Passeriformes. Coraciiformes (light blue in (A), missing from (B)) contains 1.7% of avian species, though APV has not yet been detected in this order.
Figure 1.
This word cloud (Wordart.com; accessed 25 March 2021) represents: (A) the proportion of the 10,721 avian species in 41 avian [14] orders. For brevity, 24 orders constituting less than 1% of avian diversity are combined into “others”; (B) the number of species detected with APV in each order. Orders are color-coded to represent the percentage of avian species found/cases of APV detected in each order, where blue and small represents <3%, green 3–7.5%, purple 7.5–11%, and red and large >11%. Although APV has been detected in more Passeriformes species than non-Passeriformes species, more APV are detected in non-Passeriformes than in Passeriformes. Coraciiformes (light blue in (A), missing from (B)) contains 1.7% of avian species, though APV has not yet been detected in this order.

Figure 2.
Proportions (number of APV species in particular order/number in order. The blue line shows the expected level of 0.035% derived from the number of species in which APV has been detected/avian species. We found that the number of species infected with APV was higher than expected in six avian orders (Anseriformes, Psittaciformes, Galliformes, Procellariformes, Accipitriformes and “others”), lower than expected in four avian orders (Passeriformes, Piciformes, Caprimulgiformes and Coraciiformes), and did not differ from expectation in the remaining five orders.
Figure 2.
Proportions (number of APV species in particular order/number in order. The blue line shows the expected level of 0.035% derived from the number of species in which APV has been detected/avian species. We found that the number of species infected with APV was higher than expected in six avian orders (Anseriformes, Psittaciformes, Galliformes, Procellariformes, Accipitriformes and “others”), lower than expected in four avian orders (Passeriformes, Piciformes, Caprimulgiformes and Coraciiformes), and did not differ from expectation in the remaining five orders.

Table 2.
Prevalence of avian poxvirus in surveys that are not identified by authors as epizootics or outbreaks.
Table 2.
Prevalence of avian poxvirus in surveys that are not identified by authors as epizootics or outbreaks.
| Order | Species | Common Name | Sample Size | Prevalence (%) | Location, Country | Period | Reference | Range |
|---|---|---|---|---|---|---|---|---|
| Galliformes | Alectoris rufa | Red-legged partridge | 70 | 41.4 | Cádiz, Spain | 2000–2001 | [40,44] | Continent |
| Galliformes | Callipepla californica | California quail | 89 | 21 | Oregon, USA | 1982–1987 | [68] | Continent |
| Galliformes | Callipepla californica | California quail | 256 | 26 | Oregon, USA | 1975–1978 | [67] | Continent |
| Galliformes | Colinus virginianus | Bobwhite quail | 2586 | 0–38.9 (mean 12.1) | Florida, Georgia, North Carolina, Tennessee, USA | 1978–1979 | [15] | Continent |
| Galliformes | Callipepla gambelii | Gambel’s Quail | 175 | 11–16 | Arizona, USA | 1963 | [60] | Continent |
| Columbiformes | Columba macroura | Morning Dove | 1795 | 2.1 | Arizona, USA | 1963 | [60] | Continent |
| Passeriformes | Haemorhous mexicanus | House Finch | 9195 | Up to 25 | California, USA | 1977–1987 | [51] | Continent |
| Passeriformes | Haemorhous mexicanus | House Finch | 174 | 5–30 | Arizona, USA | 2011 | [33] | Continent |
| Passeriformes | Corvus corone Corvus macrorhynchos | Carrion crow, Large-billed crow | 2036 | 17.6 | Hokkaido, Japan | 2007–2012 | [47] | Islands |
| Passeriformes | Nesomimus parvulus | Galapagos mockingbird | 417 | 18.9 | Galapagos, Ecuador | 1982–1983 | [49] | Islands |
| Passeriformes | Nesomimus parvulus | Galapagos mockingbird | 117 | 16 | Galapagos, Ecuador | 1979–1980 | [69] | Islands |
| Passeriformes | Anthus berthelotti | Berthelot´s pipit | 139 | 28 | Canary Islands, Spain | 2002–2003 | [48] | Islands |
| Passeriformes | Calandrella rufescens | Short-toed lark | 395 | 50 | Canary Islands, Spain | 2002–2003 | [48] | Islands |
| Passeriformes | Chasiempis sanswichensis | Hawaii ‘elepaio | 97 | Up to 40 | Hawaii, USA | 1994–1997 | [37] | Islands |
| Passeriformes | SEVERAL | Galapagos finches (7 spp.) | 545 | 4–9 | Galapagos, Ecuador | 2008 and 2009 | [70] | Islands |
| Procellariiformes | Phoebastria inmutabilis | Laysan albatross | 92 | Up to 88 | Hawaii, USA | 2004 and 2006 | [42] | Islands |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Williams, R.A.J.; Truchado, D.A.; Benitez, L. A Review on the Prevalence of Poxvirus Disease in Free-Living and Captive Wild Birds. Microbiol. Res. 2021, 12, 403-418. https://0-doi-org.brum.beds.ac.uk/10.3390/microbiolres12020028
AMA Style
Williams RAJ, Truchado DA, Benitez L. A Review on the Prevalence of Poxvirus Disease in Free-Living and Captive Wild Birds. Microbiology Research. 2021; 12(2):403-418. https://0-doi-org.brum.beds.ac.uk/10.3390/microbiolres12020028
Chicago/Turabian StyleWilliams, Richard A. J., Daniel A. Truchado, and Laura Benitez. 2021. "A Review on the Prevalence of Poxvirus Disease in Free-Living and Captive Wild Birds" Microbiology Research 12, no. 2: 403-418. https://0-doi-org.brum.beds.ac.uk/10.3390/microbiolres12020028