Organic Farming Improves Soil Microbial Abundance and Diversity under Greenhouse Condition: A Case Study in Shanghai (Eastern China)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sites Description
2.2. Soil Collection
2.3. Soil Chemical Analysis
2.4. Soil Microbial Analysis
2.5. Statistical Analysis
3. Results
3.1. Soil Chemical Properties
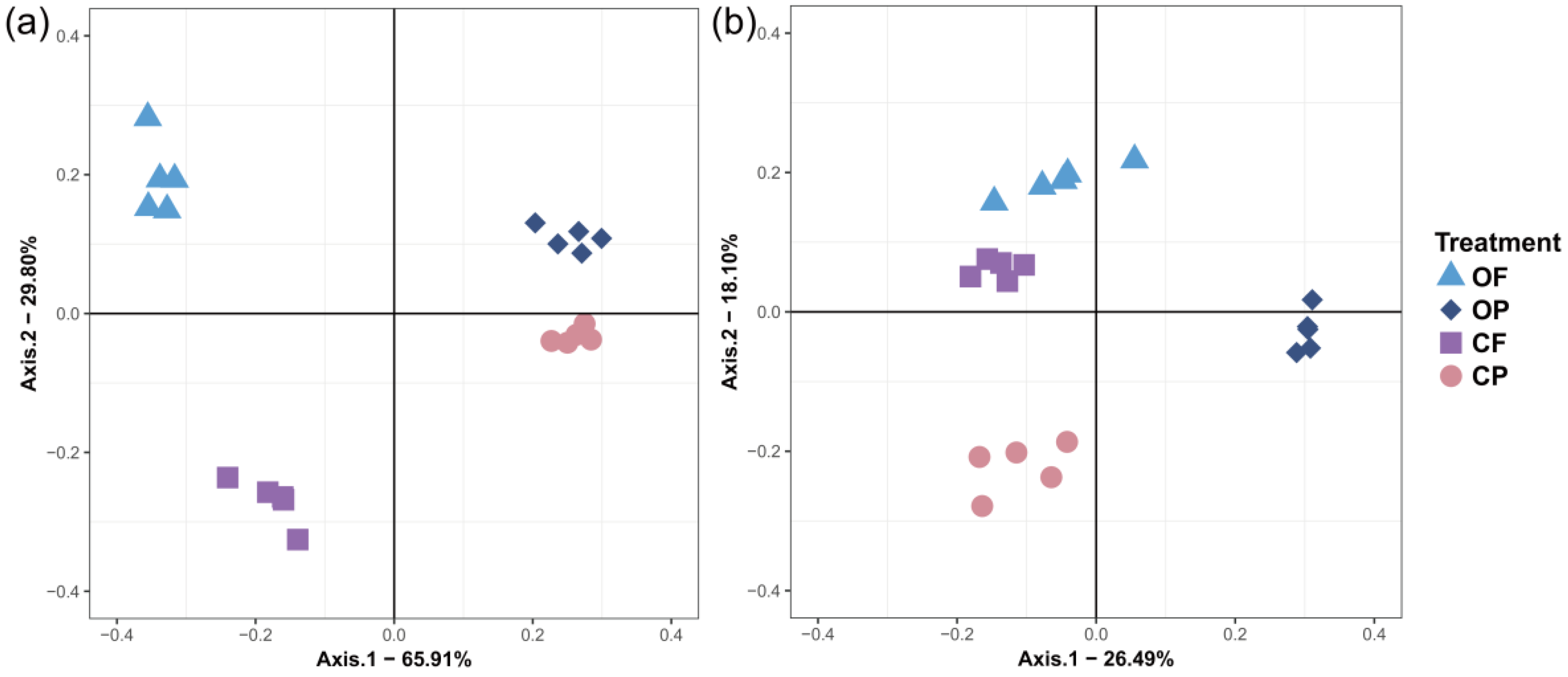
3.2. Microbial α- and β-Diversity
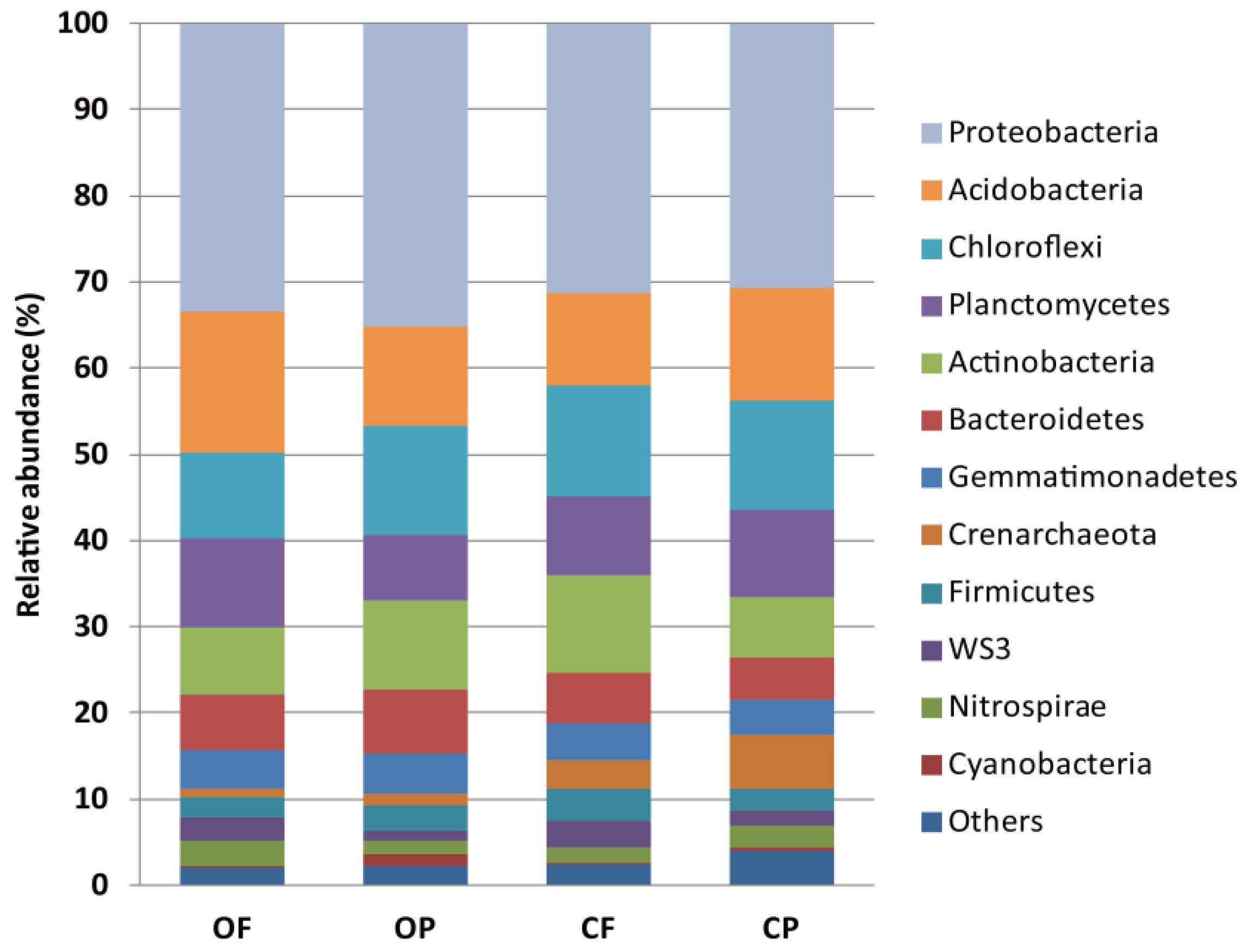
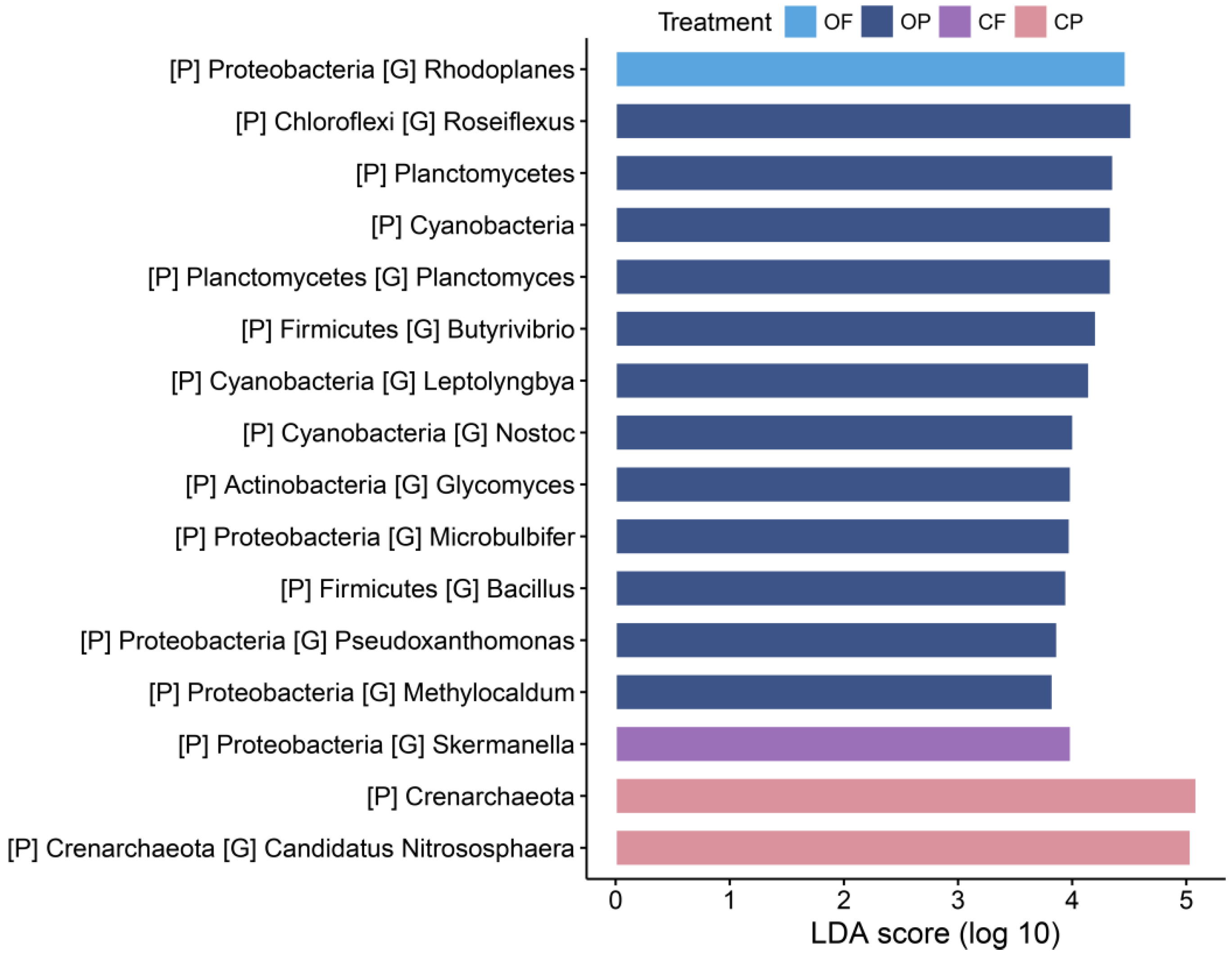
3.3. Cultivation System-Associated Microbial Taxa
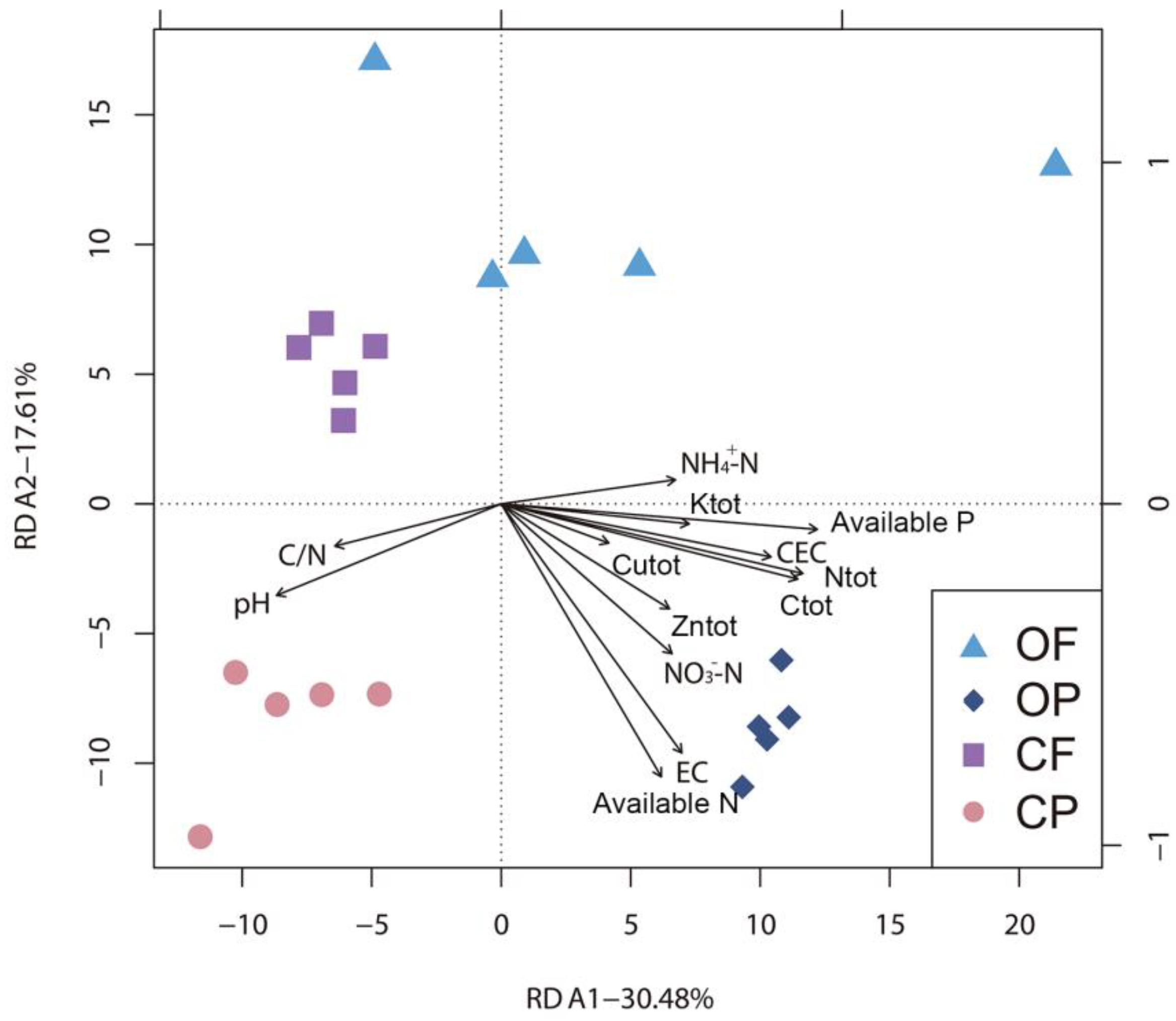
3.4. Relationships between the Microbial Community and Edaphic Properties
4. Discussion
4.1. Agricultural Management Affects Soil Properties
4.2. Soil Microbial Community Shifted by Cultivation Systems
4.3. Differential Taxa for Cultivation Systems
4.4. Relationships between Soil Properties and Microbial Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Doran, J.W.; Zeiss, M.R. Soil health and sustainability: Managing the biotic component of soil quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef]
- Bacon, C.W.; Palencia, E.R.; Hinton, D.M. Abiotic and Biotic Plant Stress-Tolerant and Beneficial Secondary Metabolites Produced by Endophytic Bacillus Species. In Plant Microbes Symbiosis: Applied Facets; Arora, N.K., Ed.; Springer India: Uttar Pradesh, India, 2015; pp. 163–177. ISBN 978-813222068-8. [Google Scholar]
- Ding, L.; Su, J.; Sun, G.; Wu, J.; Wei, W. Increased microbial functional diversity under long-term organic and integrated fertilization in a paddy soil. Appl. Microbiol. Biotechnol. 2018, 102, 1969–1982. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [PubMed]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Bruggen, A.H.C.; Semenov, A.M.; Van Diepeningen, A.D.; De Vos, O.J.; Blok, W.J. Relation between soil health, wave-like fluctuations in microbial populations, and soil-borne plant disease management. Eur. J. Plant. Pathol. 2006, 115, 105–122. [Google Scholar] [CrossRef]
- Degrune, F.; Dufrêne, M.; Colinet, G.; Massart, S.; Taminiau, B.; Bodson, B.; Hiel, M.P.; Daube, G.; Nezer, C.; Vandenbol, M. A novel sub-phylum method discriminates better the impact of crop management on soil microbial community. Agron. Sustain. Dev. 2015, 35, 1157–1166. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Perez, P.G.; Zhang, R.; Nielsen, S.; Huang, D.; Thomas, T. Effects of different C/N ratios on bacterial compositions and processes in an organically managed soil. Biol. Fertil. Soils 2018, 54, 137–147. [Google Scholar] [CrossRef]
- Xiong, W.; Li, Z.; Liu, H.; Xue, C.; Zhang, R.; Wu, H.; Li, R.; Shen, Q. The Effect of Long-Term Continuous Cropping of Black Pepper on Soil Bacterial Communities as Determined by 454 Pyrosequencing. PLoS ONE 2015, 10, e0136946. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Bueno, N.; Valenzuela-Encinas, C.; Marsch, R.; Ortiz-Gutiérrez, D.; Verhulst, N.; Govaerts, B.; Dendooven, L.; Navarro-Noya, Y. Bacterial indicator taxa in soils under different long-term agricultural management. J. Appl. Microbiol. 2016, 120, 921–933. [Google Scholar] [CrossRef] [PubMed]
- Bünemann, E.K.; Schwenke, G.D.; Van Zwieten, L. Impact of agricultural inputs on soil organisms—A review. Aust. J. Soil Res. 2006, 44, 379–406. [Google Scholar] [CrossRef]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Xi, X.; Tang, X.; Luo, D.; Gu, B.; Lam, S.K.; Vitousek, P.M.; Chen, D. Policy distortions, farm size, and the overuse of agricultural chemicals in China. Proc. Natl. Acad. Sci. USA 2018, 115, 7010–7015. [Google Scholar] [CrossRef] [PubMed]
- Gomiero, T.; Pimentel, D.; Paoletti, M.G. Environmental impact of different agricultural management practices: Conventional vs. Organic agriculture. Crit. Rev. Plant. Sci. 2011, 30, 95–124. [Google Scholar] [CrossRef]
- Organic Agriculture Worldwide 2016: Current Statistics. Available online: http://orgprints.org/32677/19/Willer-2018-global-data-biofach.pdf (accessed on 14 February 2018).
- Wang, W.; Wang, H.; Feng, Y.; Wang, L.; Xiao, X.; Xi, Y.; Luo, X.; Sun, R.; Ye, X.; Huang, Y.; et al. Consistent responses of the microbial community structure to organic farming along the middle and lower reaches of the Yangtze River. Sci. Rep. 2016, 6, 35046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, A.; Dick, W.A. Bacterial Community Diversity in Soil Under two Tillage Practices as Determined by Pyrosequencing. Microb. Ecol. 2015, 70, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, A.L.; Sheaffer, C.C.; Wyse, D.L.; Staley, C.; Gould, T.J.; Sadowsky, M.J. Structure of bacterial communities in soil following cover crop and organic fertilizer incorporation. Appl. Microbiol. Biot. 2016, 100, 9331–9341. [Google Scholar] [CrossRef] [PubMed]
- Gomez, E.; Ferreras, L.; Toresani, S. Soil bacterial functional diversity as influenced by organic amendment application. Bioresour. Technol. 2006, 97, 1484–1489. [Google Scholar] [CrossRef] [PubMed]
- Lupwayi, N.Z.; Larney, F.J.; Blackshaw, R.E.; Kanashiro, D.A.; Pearson, D.C.; Petri, R.M. Pyrosequencing reveals profiles of soil bacterial communities after 12 years of conservation management on irrigated crop rotations. Appl. Soil Ecol. 2017, 121, 65–73. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, Y.; Lu, S.; Xiang, Q.; Yu, X.; Zhao, K.; Zou, L.; Chen, Q.; Tu, S.; Zhang, X. Long-term Fertilization Structures Bacterial and Archaeal Communities along Soil Depth Gradient in a Paddy Soil. Front. Microbiol. 2017, 8, 1516. [Google Scholar] [CrossRef] [PubMed]
- Bonanomi, G.; De Filippis, F.; Cesarano, G.; La Storia, A.; Ercolini, D.; Scala, F. Organic farming induces changes in soil microbiota that affect agro-ecosystem functions. Soil Biol. Biochem. 2016, 103, 327–336. [Google Scholar] [CrossRef]
- Kang, Y.; Chang, Y.C.A.; Choi, H.S.; Gu, M. Current and future status of protected cultivation techniques in Asia. Acta Hortic. 2013, 987, 33–40. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2006; World Soil Resources Report; FAO: Rome, Italy, 2010. [Google Scholar]
- Ma, B.; Li, X.; Chang, S.X. Capping material type affects rhizosphere bacteria community structure in the cover soil in oil sands reclamation. J. Soil Sediments 2017, 17, 2516–2523. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoč, T.; Salzberg, S.L. Flash: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microb. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Whalen, J.K.; Chang, C.; Clayton, G.W.; Carefoot, J.P. Cattle Manure Amendments Can Increase the pH of Acid Soils. Soil Sci. Soc. Am. J. 2000, 64, 962–966. [Google Scholar] [CrossRef]
- Ge, T.; Nie, S.A.; Wu, J.; Shen, J.; Xiao, H.A.; Tong, C.; Huang, D.; Hong, Y.; Iwasaki, K. Chemical properties, microbial biomass, and activity differ between soils of organic and conventional horticultural systems under greenhouse and open field management: A case study. J. Soil Sediments 2010, 11, 25–36. [Google Scholar] [CrossRef]
- Teng, Y.; Wu, J.; Lu, S.; Wang, Y.; Jiao, X.; Song, L. Soil and soil environmental quality monitoring in China: A review. Environ. Int. 2014, 69, 177–199. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.; Si, Y.; Xing, Y.; Li, Y. Illumina MiSeq sequencing investigation on the contrasting soil bacterial community structures in different iron mining areas. Environ. Sci. Pollut. Res. 2015, 22, 10788–10799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, D.; Li, F. Complexities surrounding China’s soil action plan. Land Degrad. Dev. 2017, 28, 2315–2320. [Google Scholar] [CrossRef]
- Bainard, L.D.; Hamel, C.; Gan, Y. Edaphic properties override the influence of crops on the composition of the soil bacterial community in a semiarid agroecosystem. Appl. Soil Ecol. 2016, 105, 160–168. [Google Scholar] [CrossRef]
- Zhang, X.; Johnston, E.R.; Li, L.; Konstantinidis, K.T.; Han, X. Experimental warming reveals positive feedbacks to climate change in the Eurasian Steppe. ISME J. 2017, 11, 885–895. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sharp, C.E.; Jones, G.M.; Grasby, S.E.; Brady, A.L.; Dunfield, P.F. Stable-Isotope probing identifies uncultured planctomycetes as primary degraders of a complex heteropolysaccharide in soil. Appl. Environ. Microb. 2015, 81, 4607–4615. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, A.; Hetz, S.; Kolb, S. Microbial responses to chitin and chitosan in oxic and anoxic agricultural soil slurries. Biogeosciences 2014, 11, 3339–3352. [Google Scholar] [CrossRef]
- Chua, J.P.S.; Orlovich, D.A.; Summerfield, T.C. Cyanobacteria in New Zealand indigenous grasslands. N. Z. J. Bot. 2014, 52, 100–115. [Google Scholar] [CrossRef] [Green Version]
- Castellano-Hinojosa, A.; Correa-Galeote, D.; Palau, J.; Bedmar, E.J. Isolation of N2-fixing rhizobacteria from Lolium perenne and evaluating their plant growth promoting traits. J. Basic Microbiol. 2016, 56, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, Q.; Chen, X.; Jiang, C. Isolation and Cultivation Methods of Actinobacteria. In Actinobacteria—Basics and Biotechnological Applications; Dhanasekaran, D., Ed.; InTech: Rijeka, Croatia, 2016; pp. 39–57. [Google Scholar]
- Abazari, M.; Zarrini, G.; Rasooli, I. Antimicrobial potentials of Leptolyngbya sp. and its synergistic effects with antibiotics. Jundishapur J. Microb. 2013, 6, 1–6. [Google Scholar]
- Ye, J.; Zhang, R.; Nielsen, S.; Joseph, S.D.; Huang, D.; Thomas, T. A combination of biochar-mineral complexes and compost improves soil bacterial processes, soil quality, and plant properties. Front. Microbiol. 2016, 7, 372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, Z.; Liu, S.; Yang, Y.; He, Z.; Ren, Z.; Zhou, J.; Li, D. Geochip-based analysis of microbial communities in alpine meadow soils in the Qinghai-Tibetan plateau. BMC Microbiol. 2013, 13, 72. [Google Scholar] [CrossRef] [PubMed]
- Schellenberger, S.; Kolb, S.; Drake, H. Metabolic responses of novel cellulolytic and saccharolytic agricultural soil Bacteria to oxygen. Environ. Microbiol. 2010, 12, 845–861. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, W. The cellulosome and cellulose degradation by anaerobic bacteria. Appl. Microbiol. Biot. 2001, 56, 634–649. [Google Scholar] [CrossRef]
- Horel, A.; Mortazavi, B.; Sobecky, P.A. Input of organic matter enhances degradation of weathered diesel fuel in sub-tropical sediments. Sci. Total Environ. 2015, 533, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Cappelletti, M.; Ghezzi, D.; Zannoni, D.; Capaccioni, B.; Fedi, S. Diversity of Methane-Oxidizing Bacteria in Soils from “Hot Lands of Medolla“ (Italy) Featured by Anomalous High-Temperatures and Biogenic CO2 Emission. Microbes Environ. 2016, 31, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, N.; Bradeen, J.; Tu, Z.; McKay, S.; Kinkel, L. Rhizosphere bacterial communities associated with long-lived perennial prairie plants vary in diversity, composition, and structure. Can. J. Microbiol. 2013, 59, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Kerou, M.; Offre, P.; Valledor, L.; Abby, S.; Melcher, M.; Nagler, M.; Weckwerth, W.; Schleper, C. Proteomics and comparative genomics of Nitrososphaera viennensis reveal the core genome and adaptations of archaeal ammonia oxidizers. Proc. Natl. Acad. Sci. USA 2016, 113, E7937–E7946. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Han, C.; Zhang, J.; Huang, Q.; Deng, H.; Deng, Y.; Zhong, W. Long-term fertilization effects on active ammonia oxidizers in an acidic upland soil in China. Soil Biol. Biochem. 2015, 84, 28–37. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Gao, X.H.; Zhang, Y.J.; Jia, M.; Lu, X.J.; Ma, Y.C.; Tian, F.; Xie, Q.; Tang, S.K. Skermanella rubra sp. nov. a bacterium isolated from the desert of Xinjiang, China. Antonie Van Leeuwenhoek 2015, 108, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Shi, Z.; Wang, H.; Wang, G. Skermanella stibiiresistens sp. nov. a highly antimony-resistant bacterium isolated from coal-mining soil, and emended description of the genus Skermanella. Int. J. Syst. Evol. Microbiol. 2012, 62, 1271–1276. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Khafipour, E.; Krause, D.O.; Entz, M.H.; de Kievit, T.R.; Fernando, W.G.D. Pyrosequencing Reveals the Influence of Organic and Conventional Farming Systems on Bacterial Communities. PLoS ONE 2012, 7, e51897. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Wu, J.; Liu, X.; Chen, X.; Wu, Y.; Yu, S. Inorganic phosphorus fertilizer ameliorates maize growth by reducing metal uptake, improving soil enzyme activity and microbial community structure. Ecotoxicol. Environ. Saf. 2017, 143, 322–329. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | OF | OP | CF | CP | ||||
|---|---|---|---|---|---|---|---|---|
| pH | 7.32 ± 0.03 | c | 7.28 ± 0.10 | c | 7.59 ± 0.26 | b | 7.83 ± 0.13 | a |
| EC (dS m−1) | 0.14 ± 0.01 | d | 0.34 ± 0.01 | a | 0.17 ± 0.01 | c | 0.19 ± 0.02 | b |
| Available N (mg kg−1) | 114.6 ± 14.3 | c | 2099 ± 410 | a | 106.1 ± 8.92 | c | 883.8 ± 31.2 | b |
| Available P (mg kg−1) | 181.7 ± 58.1 | b | 247.6 ± 32.0 | a | 9.40 ± 4.63 | c | 30.5 ± 4.63 | c |
| Ctot (%) | 1.93 ± 0.40 | b | 2.83 ± 0.45 | a | 1.33 ± 0.16 | c | 1.24 ± 0.04 | c |
| Ntot (%) | 0.19 ± 0.05 | b | 0.29 ± 0.04 | a | 0.13 ± 0.01 | c | 0.12 ± 0.01 | c |
| Ktot (%) | 2.06 ± 0.30 | ab | 2.28 ± 0.16 | a | 1.89 ± 0.16 | b | 1.21 ± 0.12 | c |
| C/N | 10.0 ± 0.50 | b | 9.74 ± 0.24 | b | 10.1 ± 0.57 | ab | 10.5 ± 0.47 | a |
| CEC (cmol(+) kg−1) | 13.2 ± 0.91 | b | 17.9 ± 1.41 | a | 12.2 ± 0.99 | b | 9.58 ± 0.50 | c |
| NO3−-N (mg kg−1) | 2.67 ± 2.07 | c | 46.4 ± 4.42 | a | 18.2 ± 13.1 | b | 3.36 ± 2.20 | c |
| NH4+-N (mg kg−1) | 7.86 ± 0.97 | a | 7.42 ± 1.08 | ab | 5.11 ± 0.47 | c | 6.40 ± 1.08 | b |
| Cutot (mg kg−1) | 17.5 ± 1.74 | ab | 23.2 ± 1.31 | a | 20.0 ± 7.94 | ab | 16.1 ± 1.15 | b |
| Zntot (mg kg−1) | 129.5 ± 13.3 | ab | 156.7 ± 3.08 | a | 120.1 ± 31.1 | b | 123.0 ± 22.1 | b |
| Cultivation System | Reads | OTUs | Coverage | ACE | Chao 1 | Shannon |
|---|---|---|---|---|---|---|
| OF | 41,616 | 5380 | 0.93 | 2227 ± 604.3 ab | 1439 ± 204.0 ab | 9.10 ± 0.19 b |
| OP | 48,983 | 5657 | 0.90 | 2738 ± 563.4 a | 1640 ± 169.9 a | 9.43 ± 0.13 a |
| CF | 36,064 | 5161 | 0.89 | 1866 ± 92.50 b | 1343 ± 85.83 b | 9.09 ± 0.08 b |
| CP | 35,842 | 5047 | 0.89 | 1810 ± 196.9 b | 1332 ± 144.0 b | 9.18 ± 0.17 b |
| Treatment | Genus | pH | EC | Available N | Available P | Ctot | Ntot | Ktot | CEC | NO3−-N | Cutot |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CF | Skermanella | 0.12 | −0.68 ** | −0.90 ** | −0.69 ** | −0.43 | −0.41 | −0.27 | −0.38 | −0.34 | −0.31 |
| CP | Candidatus Nitrososphaera | 0.66 ** | 0.02 | −0.28 | −0.74 ** | −0.81 ** | −0.86 ** | −0.84 ** | −0.85 ** | −0.33 | −0.60 |
| OF | Rhodoplanes | −0.64 ** | −0.34 | −0.10 | 0.57 | 0.71 ** | 0.72** | 0.62** | 0.66 ** | 0.04 | 0.30 |
| OP | Roseiflexus | −0.49 | 0.76 ** | 0.75 ** | 0.65 ** | 0.69 ** | 0.70 ** | 0.48 | 0.73 ** | 0.75 ** | 0.64 ** |
| Pseudoxanthomonas | −0.06 | 0.62 ** | 0.71 ** | 0.54 | 0.57 | 0.49 | 0.54 | 0.46 | 0.45 | 0.45 | |
| Butyrivibrio | −0.57 | 0.59 | 0.60 | 0.67 ** | 0.72 ** | 0.74 ** | 0.48 | 0.74 ** | 0.61 ** | 0.62 ** | |
| Leptolyngbya | −0.44 | 0.67 ** | 0.71 ** | 0.64 ** | 0.47 | 0.53 | 0.37 | 0.55 | 0.36 | 0.46 | |
| Nostoc | −0.23 | 0.74 ** | 0.64 ** | 0.43 | 0.51 | 0.50 | 0.69 ** | 0.58 | 0.82 ** | 0.54 | |
| Glycomyces | −0.35 | 0.56 | 0.54 | 0.43 | 0.59 | 0.56 | 0.33 | 0.58 | 0.53 | 0.61 ** | |
| Microbulbifer | −0.12 | 0.69 ** | 0.88 ** | 0.60 ** | 0.50 | 0.42 | 0.51 | 0.46 | 0.47 | 0.49 | |
| Bacillus | −0.16 | 0.58 | 0.53 | 0.44 | 0.62 ** | 0.53 | 0.32 | 0.49 | 0.58 | 0.49 | |
| Planctomyces | −0.05 | 0.67 ** | 0.86 ** | 0.57 | 0.45 | 0.36 | 0.41 | 0.30 | 0.35 | 0.33 | |
| Methylocaldum | −0.58 | 0.49 | 0.54 | 0.68 ** | 0.64 ** | 0.70 ** | 0.64 ** | 0.72 ** | 0.45 | 0.64 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, J.; Liang, Y.; Huang, D. Organic Farming Improves Soil Microbial Abundance and Diversity under Greenhouse Condition: A Case Study in Shanghai (Eastern China). Sustainability 2018, 10, 3825. https://0-doi-org.brum.beds.ac.uk/10.3390/su10103825
Liao J, Liang Y, Huang D. Organic Farming Improves Soil Microbial Abundance and Diversity under Greenhouse Condition: A Case Study in Shanghai (Eastern China). Sustainability. 2018; 10(10):3825. https://0-doi-org.brum.beds.ac.uk/10.3390/su10103825
Chicago/Turabian StyleLiao, Jianli, Yun Liang, and Danfeng Huang. 2018. "Organic Farming Improves Soil Microbial Abundance and Diversity under Greenhouse Condition: A Case Study in Shanghai (Eastern China)" Sustainability 10, no. 10: 3825. https://0-doi-org.brum.beds.ac.uk/10.3390/su10103825