The Responses of Plant Leaf CO2/H2O Exchange and Water Use Efficiency to Drought: A Meta-Analysis

1
International Institutes for Earth System Science, Nanjing University, Nanjing 210023, China
2
Jiangsu Provincial Key Laboratory of Geographic Information Science and Technology, Nanjing University, Nanjing 210023, China
3
Zhejiang A&F University, Lin’an, Zhejiang 311300, China
4
School of Earth Sciences and Engineering, Hohai University, No. 8 Fo cheng xi Road, Nanjing 211100, China
*
Author to whom correspondence should be addressed.
Sustainability 2018, 10(2), 551; https://0-doi-org.brum.beds.ac.uk/10.3390/su10020551
Submission received: 27 December 2017
/
Revised: 9 February 2018
/
Accepted: 19 February 2018
/
Published: 21 February 2018
(This article belongs to the Special Issue Impacts of Climate Change on Hydrology, Water Quality and Ecology)
Abstract
:Persistent drought severely inhibits plant growth and productivity, which negatively affects terrestrial primary productivity worldwide. Therefore, it is important to investigate the impacts of drought on plant leaf CO2/H2O exchange and water use efficiency. This study assessed the responses of net photosynthesis (Pn), stomatal conductance (Gs), transpiration (Tr), and instantaneous water use efficiency (WUE) to drought based on a worldwide meta-analysis of 112 published studies. The results demonstrated that drought decreased Pn, Tr, and Gs significantly and differently among different moderators. C4 plants had smaller Pn reduction than C3 plants, which gives C4 plants an advantage in Pn. But their WUE decreased under drought conditions, indicating a great flexibility in C4 WUE. Annual herbs sacrificed WUE (−6.2%) to maintain efficient Pn. Perennial herbs took a different strategy in response to drought with an increased WUE (25.1%). Deciduous tree species displayed a greater increase in WUE than conifers and evergreen species. Additionally, Gs had a significant correlation with Pn and Tr, but an insignificant correlation with WUE, which could be because WUE is affected by other factors (e.g., air flow, CO2 concentration, and relative humidity). These findings have significant implications for understanding the worldwide effects of drought on plant leaf CO2/H2O exchange and water use efficiency.
1. Introduction
With a changing climate, drought has been predicted to be more intense and frequent in many regions of the world [1]. Water scarcity has become one of the most adverse environmental factors for plant growth and productivity, limiting terrestrial primary productivity worldwide, especially in the semi-arid regions of the northern hemisphere [2,3]. As a primary resource for plant growth and productivity, water is directly or indirectly involved in all physiological processes. Plant leaf CO2/H2O exchange and water use efficiency (WUE, the ratio of net photosynthesis (Pn) to transpiration (Tr)) are important parameters for estimating vegetation productivity and for planning water use in arid and semi-arid areas. An increased WUE can improve plant productivity, and it is necessary for securing environmental sustainable food production in drier environments [4]. Therefore, the effect of drought on leaf CO2/H2O exchange and WUE is a major issue, especially with a further changing climate [5,6,7].
The effect of drought on plant leaf CO2/H2O exchange and WUE has been intensively studied. According to previous studies, stomata not only control CO2 flux, but also regulate water flux in plants to reduce water loss and, therefore, affect plant WUE [8,9,10,11,12]. The earliest leaf defense against desiccation is stomatal closure to avoid excessive water loss [13] since it is much quicker than changes in the root system, leaf area, and chloroplast ultrastructure. Decreasing the supply of CO2 to mesophyll tissue under drought conditions is widely recognized as the primary effect of drought on carbon assimilation [4,14,15,16]. During this process, abscisic acid (ABA) reaches the leaves through the transpiration stream for drought-induced root-to-leaf signaling, which induces the stomata closure. Stomata also control Tr, which can cool leaf temperatures during drought through water loss [17].
WUE is a key physiological parameter for plant selection. It can be measured at different scales, ranging from instantaneous measurements on the leaf to more integrative measurements at the plant and crop level [18]. Most studies of WUE have used instantaneous measurements of leaf photosynthesis and transpiration to characterize environmental effects [6,19,20], although one study evaluated WUE at the whole-plant level and compared the instantaneous and whole-plant values [18]. In general, WUE has been reported to increase [5,12,21,22] and improve productivity and reduce water stress under drought conditions [7]. As portable equipment for measuring leaf gas exchange rates facilitates the simultaneous measurement of photosynthesis and transpiration, WUE is an early response indicator and an important index for studying the response of plant physiology and ecology to global environmental change.
The responses of plant leaf CO2/H2O exchange and WUE to drought differ with the characteristics of the drought event (e.g., the speed, severity, and duration) [23] and with plant endogenous factors (e.g., individual species, growth stage, and plant form) [4,24,25,26]. Moreover, the results may be different within one species in one experiment [27]. Therefore, there is an urgent need to integrate results across studies to estimate the effects of drought on plant leaf CO2/H2O exchange and WUE. In recent years, a number of publications have focused on meta-analysis of plants under stress conditions; this study method is favored because it summarizes and integrates the results from numerous independent experiments while accounting for variability across experiments [28,29]. Meta-analyses have been used to study the responses of plant physiological traits [30] and grain yield [31,32] to drought conditions; and the method has been used to study the responses of plant leaf gas exchange and growth under elevated CO2 and temperature [33,34]. Researchers have used meta-analysis to explore the central tendency and identify different patterns of leaf gas exchange and water status responses to drought under different moderators (e.g., drought intensity, metabolic CO2 assimilation, and growth forms) [30]. However, few studies have applied meta-analysis to the worldwide WUE response of plants to an environmental factor.
In this study, we conducted a meta-analysis of plant leaf CO2/H2O exchange (Pn, stomatal conductance (Gs), and Tr) and WUE of plants under drought conditions. Our aim was to determine the responses of plant leaf CO2/H2O exchange and WUE to drought. Furthermore, we explored different responses between herb life forms and tree foliage types because foliage types can summarize physiological strategies for water use efficiency and growth patterns in some plant species [35,36,37]. The objectives of this study were to investigate (1) how drought intensities, photosynthetic pathways (C3 and C4), and plant forms (herbs, shrubs, trees, and lianas) affect the responses of WUE, Pn, Tr, and Gs to drought; (2) the differences between different moderators (e.g., annual herbs (A-herbs) and perennial herbs (P-herbs) and tree foliage types (conifer, deciduous, and evergreen)) under drought; and (3) the relationships between Gs, Pn, Tr, and WUE for the two photosynthetic pathways and four plant forms.
2. Methods
2.1. Data Collection
The meta-analysis data were collected from peer reviewed journal articles, which were reported or online in the Web of Science and the China Knowledge Resource Integrated Databases. The articles, covering the last 30 years, were selected by the subject term combinations: drought/water stress and water use efficiency/photosynthesis/gas exchange. Finally, there were 112 published papers selected from more than 600 published related papers. The selected criteria are summarized as follows: (1) the data reported in the papers must be from experiments, including two datasets: control and treatment; (2) all the experiments should report at least one of the observations of plant species (e.g., water use efficiency (WUE), net photosynthesis rate (Pn), transpiration (Tr) and stomatal conductance (Gs). (3) All the observations in the control and treatment groups should have the mean, standard deviation/error and sample size directly extracted from the context, tables and digitized graphs.
Furthermore, datasets were grouped according to (i.) drought intensity or relative soil water content (RSWC), (ii.) photosynthesis pathways, (iii.) plant forms, (iv.) herb life forms and (v.) foliage types. These groups are used as moderators in the meta-analysis to distinguish effective values across and at different moderators, in addition to the overall effects. If the plant forms, foliage type and herb life-form had not been elaborated in the research papers; we would utilize the information from the website of Flora of China: http://frps.eflora.cn/. Relevant environment data were also collected: air temperature (Ta), air flow (Af), CO2 concentration (CO2C, air relative humidity (RH), leaf temperature (Tl), photosynthetically active radiation (PAR), and leaf to air vapor pressure deficit (VPD).
- (i.)
- Drought intensity is classified into three levels (mild, moderate and severe). Where the drought intensity was not provided in the studies, it would be classified according to the RSWC: mild stress (55% ≤ RSWC < 75%), moderate stress (40% ≤ RSWC < 55%) and severe stress (RSWC < 40%).
- (ii.)
- Photosynthesis pathways include the C3 and C4 photosynthetic pathway (C3 and C4 plants), but we did not take crassulacean acid metabolism (CAM) photosynthesis into consideration.
- (iii.)
- Plant forms (e.g., trees, shrubs, lianas and herbs) have widely different mechanical architecture, which can also vary phenotypically with the environment.
- (iv.)
- Herb life forms: annual and perennial herbs. A-herbs and P-herbs are short for annual herbs and perennial herbs, respectively.
- (v.)
- Foliage types: there are three different foliage types, including needle-leaved evergreen conifers (conifer), evergreen broadleaves (evergreen) and deciduous broadleaves (deciduous).
It should be noted that a total of 124 plant species (92 genera in 44 families) are recorded, and 1690 observations are selected from 112 published papers, including 521 observations of Pn, 404 observations of Tr, 356 observations of Gs and 409 observations of WUE.
2.2. Meta-Analysis
The effect size for each observation is calculated as the response ratio (LnR) to represent the magnitude of the responses of plant leaf CO2/H2O exchange and WUE to drought:
where Xe and Xc are the mean response values of each individual observation in the treatment and control groups. The variance of the response ratio (LnR) was calculated:
where Se and Sc are the standard deviations for the treatment and control groups; ne and nc are the sample sizes for the treatment and control groups; Xe and Xc are the same indicators as in Equation (1). In order to improve the accuracy of LnR and reduce its variability, the mean weighted response ratio (LnR+) is calculated from LnR:
where i is the categorical subdivision for group analysis; the reciprocal of the variance () is the weight of each LnR and calculated as follows:
The meta-analyses are performed using METAWIN 2.0 (Sinauer Associates, Sunderland, UK) [38]. The effect is analyzed by the random effect model. The mean effect size of each categorical subdivision is calculated, and if the 95% confidence intervals (CI) of LnR+ do not overlap with zero, this means that the drought has a significant promoting or inhibiting effect on plant leaf CO2/H2O exchange and WUE. The bigger the value is, the greater the influence of drought on the plants.
Then, the LnR+ is reported as the percent change (D, %), which is more visible than LnR+ and calculated as follows:
Regression analyses are performed to establish the relationships between Gs, Pn, Tr and WUE with environmental factors under drought in all plant species, two photosynthesis pathways and four plant forms. The Gs, Pn, Tr and WUE for this regression analysis are dependent and measured from the same leaf at the same time. The LnR is effect size for Gs, Pn, Tr and WUE and calculated as Equation (1). Figures are performed with Origin Pro 9.0 software (OriginLab Corporation, Northampton, MA, USA) with Microsoft Windows.
3. Results
3.1. Effects of Drought on Plant Leaf CO2/H2O Exchange and WUE
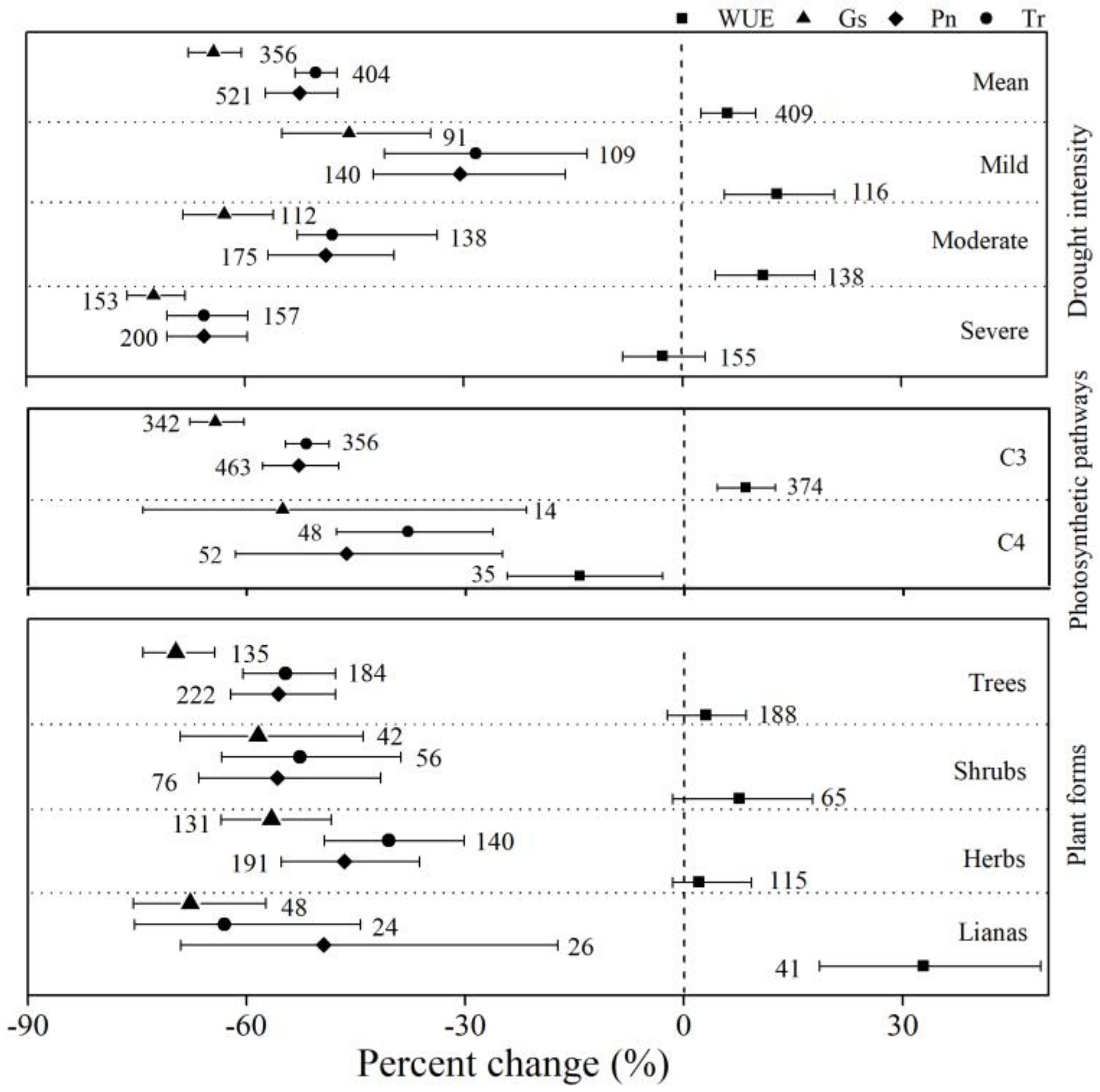
As shown in Figure 1, drought showed a significant, adverse influence on plant leaf CO2/H2O exchange. Across all studies, drought decreased Pn, Tr, and Gs by 52.5%, 50.3%, and 64.3%, respectively, but increased WUE by 6.2% (Figure 1). Gs decreased more than Pn and Tr in all studies, regardless of the different groups. Drought decreased the Pn, Tr, and Gs of C3 plants by 52.9%, 51.8%, and 64.3%, respectively, which were greater than the decreases in Pn, Tr, and Gs for C4 plants (−46.3%, −37.9%, and −55.1%, respectively). However, drought increased the WUE in C3 plants by 8.4% and decreased the C4 plants’ WUE by 14.3%. Drought intensity showed an adverse influence on plant leaf CO2/H2O exchange, with the most adverse effects appearing under severe stress. The WUE increased by 13.0% and 11.1% under mild and moderate stress, indicating a significant promoting effect. Severe stress decreased WUE by 2.7%, although the 95% CI overlapped with zero. Drought decreased the Pn, Tr, and Gs of herbs by 46.6%, 40.5%, and 56.6%, respectively (Figure 1), and this was the lowest percent change among the four plant forms. The percent change of WUE for trees, shrubs, and herbs overlapped with zero, showing an insignificant effect of drought. The percent change of WUE was the highest for lianas (32.9%), indicating that drought had the greatest promoting effect on lianas among the four plant forms.
3.2. Effects of Drought on Plant Leaf CO2/H2O Exchange and WUE in C3 and C4 Plants
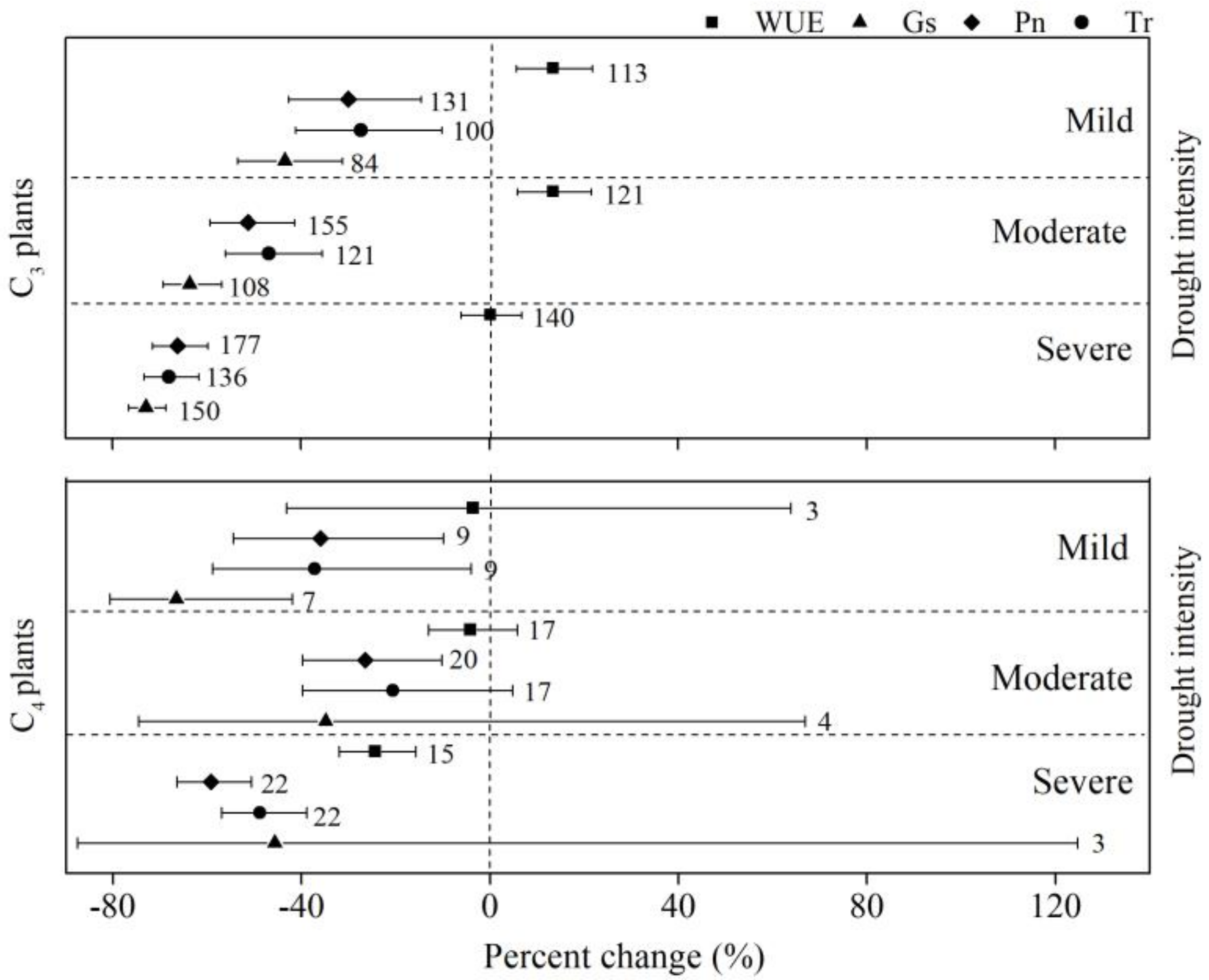
The results showed that the changes in Pn, Tr, and Gs in C3 plants were smaller than for C4 plants under mild stress, but larger than C4 plants under moderate and severe stress (Figure 2). The percent change of WUE for C3 plants was the same (13.5%) under mild and moderate stress, but the percent change overlapped with zero under severe stress (Figure 2). The WUE of C4 plants decreased by 3.5%, 4.1%, and 24.3% under mild, moderate, and severe stress, respectively, with the 95% CI overlapping with zero under mild and moderate stress.
3.3. Effects of Drought on Plant Leaf CO2/H2O Exchange and WUE of Herb Life Forms and Tree Foliage Types
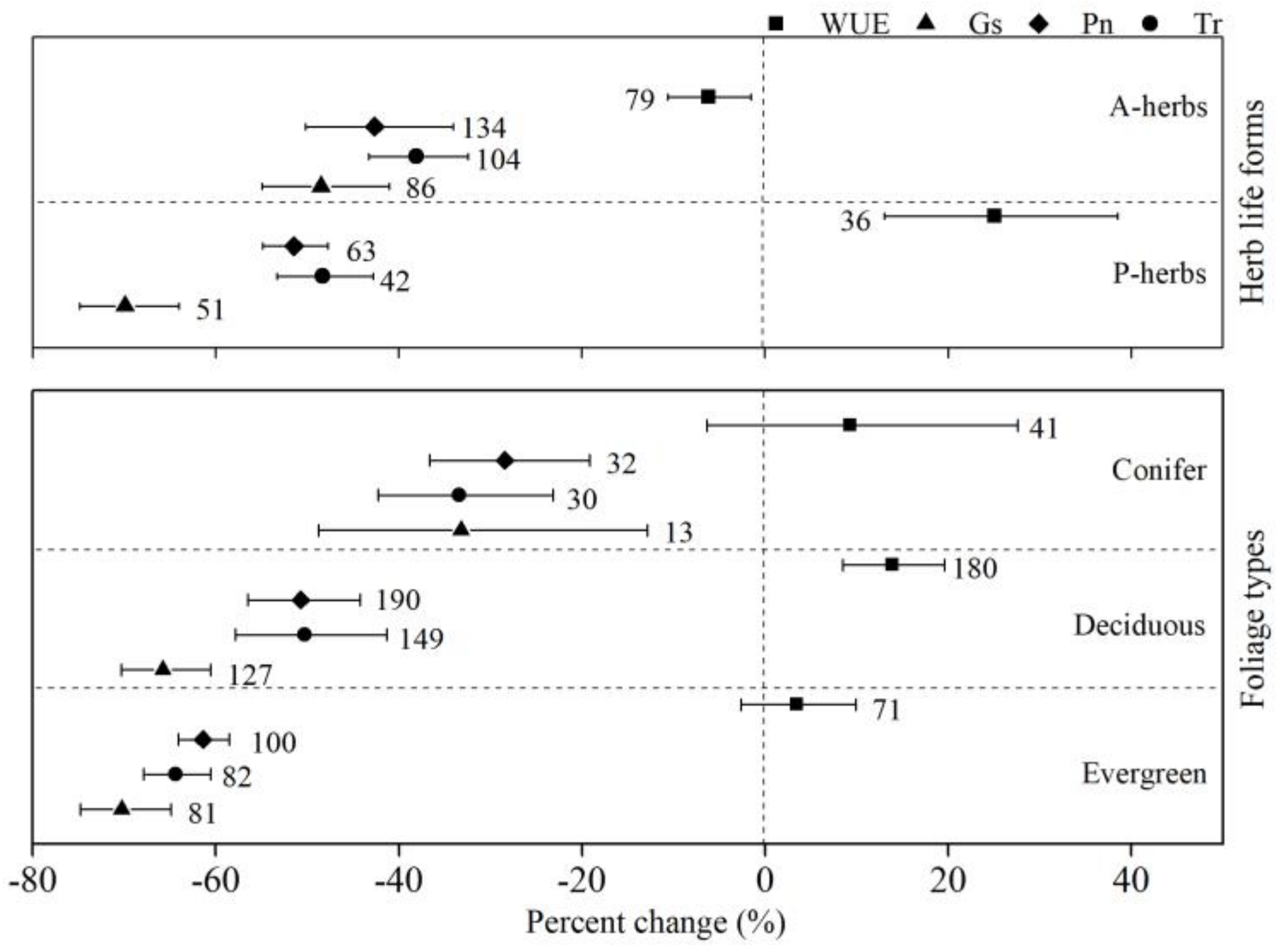
Herb life forms showed different responses to drought. P-herbs showed larger changes in Pn, Tr, and Gs (−51.4%, −48.3%, and −69.9%, respectively) than A-herbs (−42.6%, −38.1%, and −48.5%, respectively). Drought increased the P-herbs’ WUE by 25.1%, but decreased the A-herbs’ WUE by 6.2%. Drought decreased Pn, Tr, and Gs of conifer plants by −28.4%, −33.4%, and −33.2%, respectively, the smallest reduction of the foliage types. Evergreen plants had the greatest decrease in Pn, Tr, and Gs (−61.4%, −64.4%, and −70.3%, respectively). Drought had the greatest promoting effect on the WUE of deciduous plants (13.9%). The 95% CIs overlapped for the percent changes of WUE for conifers and evergreen plants and the intervals overlapped with zero (Figure 3).
3.4. Relationship of Gs, Pn, Tr and WUE With Environmental Factors
The results showed that there was a significant positive correlation between Gs and Pn (R2 = 0.5087, P < 0.01) across all plant types (Table 1). There was a positive correlation between Gs and Pn in both C3 plants (R2 = 0.4968, P < 0.01) and C4 plants (R2 = 0.7545, P < 0.01). Gs was correlated with Pn in trees, shrubs, herbs, and lianas (49.21%, 53.3%, 50.46%, and 72.1%, respectively (P < 0.01)) (Table 1). Our results showed a positive correlation between Gs and Tr at different moderators (P < 0.01). Gs was correlated with Tr for all species, C3 plants, and C4 plants (72.06%, 77.94%, and 76.57%, respectively). Gs was also correlated with Tr for trees, shrubs, herbs, and lianas (90.48%, 53.09%, 48.29%, and 87.37%, respectively). Gs and WUE had an insignificant negative correlation in different groups, although shrubs had a positive correlation. Gs was correlated with WUE for all plants (16.15% (P < 0.01)), but only 1.04% (P = 0.12) and 15.47% (P = 0.16) correlations for C3 and C4 plants, respectively. The correlation between Gs and WUE was 3.49% (P = 0.076), 13.67% (P = 0.783), 23.79%, and 52.7% (P < 0.01) for trees, shrubs, herbs, and lianas, respectively.
Pearson correlation analysis showed that the response ratios of the Pn to drought did not exhibit any significant correlation with air flow (Af), CO2 concentration (CO2C), air relative humidity (RH), leaf temperature (Tl), photosynthetically active radiation (PAR) or leaf to air vapor pressure deficit (VPD) (Table 2), but a significant positive with air temperature (Ta) (P < 0.05). The response ratios of Tr to drought was observed a significant negative relationship with Af and a positive relationship with RH (P < 0.05). Significant positive relationships were observed between the response ratios of Gs to drought with CO2C, RH (P < 0.01) and Tl (P < 0.05). WUE had a significant positive relationship with Af (P < 0.01), but significant negative relationships with CO2C (P < 0.05) and RH (P < 0.01).
4. Discussion
Our meta-analysis was the first comprehensive overview of the effects of drought on plant leaf CO2/H2O exchange and WUE. We found that drought strongly inhibited plant leaf CO2/H2O exchange (Figure 1), and the overall inhibitory effect increased with increasing drought stress. We found that Gs decreased more than Pn and Tr across all studies and in different groups, which was consistent with previous meta-analysis results [30]. The governing of stomata opening and closure is crucial for terrestrial plants (particularly C3 plants) to balance carbon acquisition and water loss by transpiration [39]. Decreased transpiration caused by the regulating mechanism to reduce Gs during drought conditions may be an adaptive behavior to protect the conducting system against hydraulic failure while restricting the entry of CO2 [40,41].
The changes in Pn and Tr controlled by Gs caused changes in WUE under drought condition. The unexpected finding was that drought increased WUE, to some extent, across all studies, and the promoting effect was found under mild and moderate stress. The decrease in Pn, Tr, and Gs and concurrent increase in WUE in response to decreased soil moisture in species is common [5,22,42]. Theoretically, WUE may be improved by partial closure of stomata so that the intercellular CO2 concentration is sufficient for saturation of Pn while the rate of water loss (Tr) can be significantly lowered under mild and moderate stress [21].
Herbs had a smaller decrease in Gs than trees under drought conditions, which supports the hypothesis that herbs adapt poorly to drought conditions [30] due to their high Tr and significant water loss in the absence of a large root system. Trees have higher drought resistance than other plants due to their lower Tr and large root systems, and this study confirmed that the deep rooting habit of trees enables them to extract water from depths of up to 8m, reducing sensitivity to surface soil water deficiency [22].
The change in WUE in trees, shrubs, herbs, and lianas were significantly different. Lianas had the largest change in WUE followed by shrubs, trees, and herbs. Drought had a marked improvement on lianas’ WUE, but the effect was not significant for trees, shrubs, and herbs (Figure 1). The Gs of lianas and trees decreased more than shrubs and herbs, indicating their superior adaptation to drought, as they benefit from strong stomatal control to avoid excessive water loss during water deficits [43]. Lianas had smaller reductions in Pn and Gs (Figure 1) than trees, which is consistent with previous research that they have better access to deep soil water than co-occurring trees [43]. Lianas had the largest increase in WUE, suggesting this plant type has an advantage over trees due to a smaller proportional decline in Pn and a larger decline in Tr during drought.
The responses of plant leaf CO2/H2O exchange and WUE to drought can vary due to other factors (i.e., species and stress intensity) [14]. Therefore, we identified the responses of plant leaf CO2/H2O exchange and WUE to drought as affected by the following combination moderators: drought intensity and photosynthetic pathways, herbs life forms, and foliage types.
4.1. Photosynthetic Pathways
The leaf CO2/H2O exchange in C3 plants was more influenced by drought than C4 plants, showing significant and larger reductions in Pn, Tr, and Gs. This suggested that C4 plants have higher carbon sequestration under drought conditions. These results are consistent with previous research that C4 plants have an advantages in Pn in drought conditions compared with C3 plants [30,44,45] because the C4 CO2-concentrating mechanism offers C4 photosynthesis a greater buffering capacity against CO2 shortages due to partial stomatal closure under water stress [46]. However, drought increased C3 WUE overall, but tended to decrease C4 WUE (Figure 1). The result seems to contradict the advantage C4 plants have over C3 plants in WUE under drought conditions [5,47]. We observed reductions in C4 WUE, especially for severely stressed plants (e.g., maize (Zea mays) [27] and some C4 grass species). Severe stress causes notable reductions in the WUE of maize in the vegetative and reproductive stages, whereas moderate stress results in significantly decreased WUE in the later growth stages [27]. Pearcy et al. [48] compared the photosynthetic type of plants limited by water deficiency, and the advantage of the C4 plant (Amaranthus retroflexus) over the C3 plant (Chenopodium album) displayed in good water conditions disappeared in drought conditions. These results indicated that drought is not the precondition for the dominance of C4 plants in many plant communities [49]. The C4 WUE is complex; the reduction in leaf Pn and Tr under water stress varies with the intensity and time of the drought, the plant species, and a variety of environmental factors. Some C4 plant species (e.g., Spartina alterniflora) have increased WUE to adapt to severe drought conditions [50]. This indicates that C4 plants have flexible WUE. Drought does, to a certain extent, increase the C3 plants’ WUE, but the WUE of C3 plants is not necessarily higher than that of C4 plants under drought conditions. Research has shown that C4 plants have a greater WUE than C3 plants in desert areas [51]. For example, a previous study found that the WUE of C4 plants was significantly higher than that of C3 species under drought stress in Flaveria species [5]. Atriples confertifolia (C4) has been found to have significantly higher WUE in summer drought conditions than Ceratoides lanata (C3), although they had similar ecotypes in similar environmental conditions [52].
4.2. Herbs Life Forms
Our study found that the lifespan of herbs (A-herbs and P-herbs) has an apparent impact on the responses of leaf CO2/H2O exchange and WUE to drought. P-herbs’ percent changes of Pn, Tr, and Gs (mainly alfalfa and grazing grass) were larger than those of A-herbs (mainly crops such as rice, maize, barley, and wheat). Also, drought decreased the WUE in A-herbs, but increased the WUE in P-herbs, showing an opposite effect (Figure 3). These findings suggest that perennial herb species have a different drought response than annual species. A large decline in the Gs of P-herbs reduced an already low Tr to avoid dehydration. Therefore, leaves and stems of P-herbs are more vulnerable to dieback from high temperatures destroying their photosynthetic mechanisms. Other plant parts (e.g., roots, caudex, and various types of underground stems) survive under or close to the ground in drought conditions. Studies have suggested that dehydration avoidance and dormancy are perennial herbs’ main strategies for survival in summer droughts in Mediterranean areas [53]. Furthermore, drought survival can impact WUE through plant mortality and stand recovery after drought. In drought conditions, annual herbs try to maintain their Pn to finish their reproductive growth through higher Gs of CO2 and higher water flux exchange than perennial herbs. This causes a reduction in annual herb WUE. Research has confirmed that A-herbs have a better WUE than perennial herbs in desert areas [51].
4.3. Foliage Types
We found that the leaf CO2/H2O exchange in conifers, evergreens, and deciduous plants decreased markedly under drought stress, but the percent changes were different. Evergreen trees had the largest changes in Pn, Tr, and Gs under drought stress (Figure 3), and conifers had the smallest change. We expected that the different responses between the three foliage types may be caused by different leaf functional anatomy (e.g., leaf area, thickness, specific leaf area, density, and stomatal morphology) [54,55], which is supported by a previous study that showed a tight coordination between leaf morphology and photosynthetic properties in water deficit [56]. These results have been found in several tree species that down-regulate photosynthesis during drought, suggesting that species with different leaf habits differ with respect to morphological, chemical, and physiological traits [17]. We found that all percent changes in WUE for coniferous, deciduous, and evergreen plants were greater than zero, especially in the deciduous species. This suggests that stomata closure in response to H2O flux is more sensitive than the response to carbon flux [22]. In addition, evergreen species are thought to have leaf traits that cause lower photosynthesis and transpiration rates, which reduces WUE [57]. Evergreen species may extend their leaf lifespans by protecting their photosynthetic machinery from overheating through evaporative cooling [17,57]. However, we found that deciduous species increased WUE under drought conditions with a smaller reduction in Tr than Pn. In addition to losing a portion of their leaves, deciduous species avoid high temperature damage by minimizing water loss from their remaining leaves [58].
4.4. Relationship of Gs, Pn, Tr and WUE with Environmental Factors
Based on the fitted results of Gs with Pn and Tr in this study and previous research [30], Gs showed significant relationship with Pn and Tr in C3 and C4 plants and the four plant forms (trees, shrubs, herbs, and lianas) on a global scale. The results indicated that stomata play a more important role in controlling transpiration rates than photosynthesis rates [30]. Many studies have determined that stomatal limitation is the dominant control of Pn and Tr reduction under drought stress [4,13,16,59,60]. Unexpectedly, Gs had a low explanation rate (16.15%) across all studies, suggesting it did not explain WUE results (Table 1). It also had low explanation rates for photosynthetic pathways (C3 and C4 plants) and plant forms (trees, shrubs, herbs, and lianas). However, our study found a negative correlation between WUE and Gs (Table 1), which is supported by previous reports that stress-induced reduction in Gs has a greater effect on Tr than Pn [61]. Therefore, WUE is a complex index that is affected by Gs and other factors. The Pearson correlation analysis showed that the response ratios of WUE to drought had a significant correlation with the air flow rate (Af), and a significant, negative relationship with CO2 concentration and relative humidity (RH) among all plants (Table 2). Previous research has suggested that WUE is in part driven by changes in environmental conditions, such as relative humidity in the air and temperature [62], and the leaf position in the canopy can also have a marked effect on instantaneous leaf WUE [18].
5. Conclusions
In this meta-analysis study, we found that drought decreased Pn, Tr, and Gs significantly and differently among different moderators. The WUE increase of lianas was greater than trees, shrubs, and herbs. C4 plants had a smaller Pn reduction than C3 plants, which gives C4 plants an advantage in Pn. But their WUE decreased under drought conditions, indicating a great flexibility in C4 WUE. We found that annual herbs sacrificed high WUE to maintain efficient Pn to finish their reproductive growth in their short life cycle. Perennial herb species have a different strategy in response to drought with an increased WUE (25.1%). Deciduous tree species displayed a greater increase in WUE than conifer and evergreen species. Additionally, we found that Gs was significantly positively correlated with Pn and Tr, but was insignificantly negatively correlated with WUE, which could explain the stress-induced reduction in Gs having a greater effect on Tr than Pn. WUE is a complex index that not only affected by Gs, but also by other factors (e.g., air flow, CO2 concentration, and relative humidity). These findings have significant implications for understanding the worldwide effects of drought on plant leaf CO2/H2O exchange and water use efficiency.
Acknowledgments
The authors thank Arthur P. Cracknell and Miss Pauline Lovell for improving the English in this paper. The study was supported by the funding partially from the NNSF of China Major Program (41601442, 41501212 & 41171324), the Fundamental Research Funds for the Central Universities (No. 2017B06814) and the fundamental research project of MOST (2005DKA32306).
Author Contributions
Jinmeng Zhang and Hong Jiang designed the study; Jinmeng Zhang performed the data collection and analysis; Jinmeng Zhang, Hong Jiang, Xinzhang Song, Jiaxin Jin and Xiuying Zhang interpreted the results and wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Trenberth, K.E.; Dai, A.; Schrier, G.V.D.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Parry, M.L.; Canziani, O.F.; Palutikof, J.P.; Linden, P.J.V.D.; Hanson, C.E. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Encycl. Lang. Linguist. 2007, 12, 171–175. [Google Scholar]
- Vargas, R.; Sonnentag, O.; Abramowitz, G.; Carrara, A.; Chen, J.M.; Ciais, P.; Correia, A.; Keenan, T.F.; Kobayashi, H.; Ourcival, J.M. Drought Influences the Accuracy of Simulated Ecosystem Fluxes: A Model-Data Meta-analysis for Mediterranean Oak Woodlands. Ecosystems 2013, 16, 749–764. [Google Scholar] [CrossRef]
- Sikder, S.; Qiao, Y.; Dong, B.; Shi, C.; Liu, M. Effect of water stress on leaf level gas exchange capacity and water-use efficiency of wheat cultivars. Indian J. Plant Physiol. 2016, 21, 300–305. [Google Scholar] [CrossRef]
- Dias, M.C.; Brüggemann, W. Water-use efficiency in Flaveria species under drought-stress conditions. Photosynthetica 2010, 48, 469–473. [Google Scholar] [CrossRef]
- Gago, J.; Douthe, C.; Florezsarasa, I.; Escalona, J.M.; Galmes, J.; Fernie, A.R.; Flexas, J.; Medrano, H. Opportunities for improving leaf water use efficiency under climate change conditions. Plant Sci. 2014, 226, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Franks, P.J.; Doheny-Adams, T.W.; Brittonharper, Z.J.; Gray, J.E. Increasing water-use efficiency directly through genetic manipulation of stomatal density. New Phytol. 2015, 207, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Jones, H. Stomatal control of photosynthesis and transpiration. J. Exp. Bot. 1998, 49, 387–398. [Google Scholar] [CrossRef]
- Galmés, J.; Conesa, M.À.; Ochogavía, J.M.; Perdomo, J.A.; Francis, D.M.; Ribascarbó, M.; Savé, R.; Flexas, J.; Medrano, H.; Cifre, J. Physiological and morphological adaptations in relation to water use efficiency in Mediterranean accessions of Solanum lycopersicum. Plant Cell Environ. 2011, 34, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Zhou, G.; Wang, Y.; Liang, Q.; Chen, J.; Cheng, Y.; Shen, R. Plant species diversity and dynamics in forests invaded by Moso bamboo (Phyllostachys edulis) in Tianmu Mountian Nature Reserve. Biodiv. Sci. 2013, 21, 288–295. [Google Scholar]
- Hentschel, R.; Hommel, R.; Poschenrieder, W.; Grote, R.; Holst, J.; Biernath, C.; Gessler, A.; Priesack, E. Stomatal conductance and intrinsic water use efficiency in the drought year 2003: A case study of European beech. Trees 2016, 30, 153–174. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Li, Y. Improving water-use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought resistant wheat. Crop J. 2017, 5, 231–239. [Google Scholar] [CrossRef]
- Lehmann, P.; Or, D. Effects of stomata clustering on leaf gas exchange. New Phytol. 2015, 207, 1015–1025. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.P.; Osório, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How Plants Cope with Water Stress in the Field? Photosynthesis and Growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Rouhi, V.; Samson, R.; Lemeur, R.; Damme, P.V. Photosynthetic gas exchange characteristics in three different almond species during drought stress and subsequent recovery. Environ. Exp. Bot. 2007, 59, 117–129. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis Under Drought and Salt Stress: Regulation Mechanisms from Whole Plant to Cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Ishida, A.; Yamazaki, J.Y.; Harayama, H.; Yazaki, K.; Ladpala, P.; Nakano, T.; Adachi, M.; Yoshimura, K.; Panuthai, S.; Staporn, D. Photoprotection of evergreen and drought-deciduous tree leaves to overcome the dry season in monsoonal tropical dry forests in Thailand. Tree Physiol. 2014, 34, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Medrano, H.; Tomás, M.; Martorell, S.; Flexas, A.; Hernández, E.; Rosselló, J.; Pou, A.; Escalona, J.M.; Bota, J. From leaf to whole-plant water use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target. Crop J. 2015, 3, 220–228. [Google Scholar] [CrossRef]
- Morison, J.I.L.; Baker, N.R.; Mullineaux, P.M.; Davies, W.J. Improving water use in crop production. Philos. Trans. R. Soc. London 2008, 363, 639–658. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C (3) plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Andersen, M.N.; Jacobsen, S.E.; Jensen, C.R. Stomatal control and water use efficiency of soybean (Glycine max L. Merr.) during progressive soil drying. Environ. Exp. Bot. 2005, 54, 33–40. [Google Scholar] [CrossRef]
- Muthuri, C.W.; Ong, C.K.; Craigon, J.; Mati, B.M.; Ngumi, V.W.; Black, C.R. Gas exchange and water use efficiency of trees and maize in agroforestry systems in semi-arid Kenya. Agric. Ecosyst. Environ. 2009, 129, 497–507. [Google Scholar] [CrossRef]
- Jaiphong, T.; Tominaga, J.; Watanabe, K.; Mai, N.; Takaragawa, H.; Suwa, R.; Ueno, M.; Kawamitsu, Y. Effects of duration and combination of drought and flood conditions on leaf photosynthesis, growth and sugar content in sugarcane. Plant Prod. Sci. 2016, 19, 1–11. [Google Scholar] [CrossRef]
- Anyia, A.O.; Herzog, H. Water-use efficiency, leaf area and leaf gas exchange of cowpeas under mid-season drought. Eur. J. Agron. 2004, 20, 327–339. [Google Scholar] [CrossRef]
- Bloemen, J.; Vergeynst, L.L.; Overlaet-Michiels, L.; Steppe, K. How important is woody tissue photosynthesis in poplar during drought stress? Trees 2016, 30, 63–72. [Google Scholar] [CrossRef]
- Reed, C.C.; Loik, M.E. Water relations and photosynthesis along an elevation gradient for Artemisia tridentata during an historic drought. Oecologia 2016, 181, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Ge, T.; Sui, F.; Bai, L.; Tong, C.; Sun, N. Effects of water stress on growth, biomass partitioning, and water-use efficiency in summer maize (Zea mays L.) throughout the growth cycle. Acta Physiolog. Plant. 2012, 34, 1043–1053. [Google Scholar] [CrossRef]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The Meta-Analysis of Response Ratios in Experimental Ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Lajeunesse, M.J. On the meta-analysis of response ratios for studies with correlated and multi-group designs. Ecology 2011, 92, 2049–2055. [Google Scholar]
- Yan, W.; Zhong, Y.; Shangguan, Z. A meta-analysis of leaf gas exchange and water status responses to drought. Sci. Rep. 2016, 6, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Mallikarjuna, S.B.; Prashant, V.; Shalabh, D.; Ahmed, H.U.; Arvind, K. Meta-analysis of grain yield QTL identified during agricultural drought in grasses showed consensus. Bmc Genom. 2011, 12, 319–337. [Google Scholar]
- Kooi, C.J.R.M.; Löw, M. Growth and yield stimulation under elevated CO2 and drought: A meta-analysis on crops. Environ. Exp. Bot. 2016, 122, 150–157. [Google Scholar]
- Curtis, P.S. A meta-analysis of leaf gas exchange and nitrogen in trees grown under elevated carbon dioxide. Plant Cell Environ. 1996, 19, 127–137. [Google Scholar] [CrossRef]
- Wang, D.; Philpott, S.M. A meta-analysis of plant physiological and growth responses to temperature and elevated CO2. Oecologia 2012, 169, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Meinzer, F.C.; Qi, J.H.; Goldstein, G.; Cao, K.F. Midday stomatal conductance is more related to stem rather than leaf water status in subtropical deciduous and evergreen broadleaf trees. Plant Cell Environ. 2013, 36, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; Holbrook, N.M. Leaf physiology does not predict leaf habit; examples from tropical dry forest. Trees 2004, 19, 290–295. [Google Scholar] [CrossRef]
- Choat, B.; Ball, M.C.; Luly, J.G.; Donnelly, C.F.; Holtum, J.A. Seasonal patterns of leaf gas exchange and water relations in dry rain forest trees of contrasting leaf phenology. Tree Physiol. 2006, 26, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.S.; Adams, D.C.; Gurevitch, J. Meta Win: Statistical Software for Meta-Analysis. Version 2.0; Sinauer Associates: Sunderland, UK, 2000. [Google Scholar]
- Brodribb, T.J.; Mcadam, S.A.M. Passive Origins of Stomatal Control in Vascular Plants. Science 2011, 331, 582–585. [Google Scholar] [CrossRef] [PubMed]
- Mcdowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Mcdowell, N.G. Mechanisms Linking Drought, Hydraulics Carbon Metabolism, and Vegetation Mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Liang, N.; Maruyama, K. Interactive effects of CO2 enrichment and drought stress on gas exchange and water-use efficiency in Alnus firma. Environ. Exp. Bot. 1995, 35, 353–361. [Google Scholar] [CrossRef]
- Chen, Y.J.; Cao, K.F.; Schnitzer, S.A.; Fan, Z.X.; Zhang, J.L.; Bongers, F. Water-use advantage for lianas over trees in tropical seasonal forests. New Phytol. 2015, 205, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Ripley, B.S.; Gilbert, M.E.; Ibrahim, D.G.; Osborne, C.P. Drought constraints on C4 photosynthesis: Stomatal and metabolic limitations in C3 and C4 subspecies of Alloteropsis semialata. J. Exp. Bot. 2007, 58, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.H.; Ripley, B.S.; Martin, T.; De-Wet, L.A.; Woodward, F.I.; Osborne, C.P. Physiological advantages of C4 grasses in the field: A comparative experiment demonstrating the importance of drought. Glob. Chang. Biol. 2014, 20, 1992–2003. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, O. C4 photosynthesis and water stress. Ann. Bot. 2009, 103, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Way, D.A.; Katul, G.G.; Manzoni, S.; Vico, G. Increasing water use efficiency along the C3 to C4 evolutionary pathway: A stomatal optimization perspective. J. Exp. Bot. 2014, 65, 3683–3693. [Google Scholar] [CrossRef] [PubMed]
- Pearcy, R.W.; Tumosa, N.; Williams, K. Relationships between growth, photosynthesis and competitive interactions for a C3 and C4 plant. Oecologia 1981, 48, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Sage, R.F.; Wedin, D.A. The Biogeography of C4 Photosynthesis: Patterns and Controlling Factors. In C4 Plant Biology; Academic Press: Cambridge, MA, USA, 1999. [Google Scholar]
- Hessini, K.; Martínez, J.P.; Gandour, M.; Albouchi, A.; Soltani, A.; Abdelly, C. Effect of water stress on growth, osmotic adjustment, cell wall elasticity and water-use efficiency in Spartina alterniflora. Environ. Exp. Bot. 2009, 67, 312–319. [Google Scholar] [CrossRef]
- Sun, H.L.; Jian-Ying, M.A.; Wang, S.M.; Zhang, X. The Study of Stable Carbon Isotope Composition in Desert Plants of Junggar Basin. J. Desert Res. 2007, 27, 972–976. [Google Scholar]
- Caldwell, M.M.; White, R.S.; Moore, R.T.; Camp, L.B. Carbon balance, productivity, and water use of cold-winter desert shrub communities dominated by C3 and C4 species. Oecologia 1977, 29, 275. [Google Scholar] [CrossRef] [PubMed]
- Lelièvre, F.; Seddaiu, G.; Ledda, L.; Porqueddu, C.; Volaire, F. Water use efficiency and drought survival in Mediterranean perennial forage grasses. Field Crops Res. 2011, 121, 333–342. [Google Scholar] [CrossRef]
- Terashima, I.; Hanba, Y.T.; Tholen, D.; Niinemets, Ü. Leaf functional anatomy in relation to photosynthesis. Plant Physiol. 2011, 155, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Tholen, D.; Boom, C.; Zhu, X.G. Opinion: Prospects for improving photosynthesis by altering leaf anatomy. Plant Sci. 2012, 197, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Binks, O.; Meir, P.; Rowland, L.; Costa, A.C.L.D.; Vasconcelos, S.S.; Oliveira, A.A.R.D.; Ferreira, L.; Mencuccini, M. Limited acclimation in leaf anatomy to experimental drought in tropical rainforest trees. Tree Physiol. 2016, 36, 1550–1561. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, K.W.; Poorter, L.; Sterck, F.J.; Borghetti, F.; Ward, D.; Bie, S.D.; Langevelde, F.V. Leaf adaptations of evergreen and deciduous trees of semi-arid and humid savannas on three continents. J. Ecol. 2013, 101, 430–440. [Google Scholar] [CrossRef]
- Hasselquist, N.J.; Allen, M.F.; Santiago, L.S. Water relations of evergreen and drought-deciduous trees along a seasonally dry tropical forest chronosequence. Oecologia 2010, 164, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Cornic, G.; Massacci, A. Leaf Photosynthesis Under Drought Stress. 1996, 5, 347–366. [Google Scholar]
- Cornic, G. Drought stress inhibits photosynthesis by decreasing stomatal aperture—Not by affecting ATP synthesis. Trends Plant Sci. 2000, 5, 187–188. [Google Scholar] [CrossRef]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.M.; Barbaroux, C.; Le, T.D.; Bréchet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides x Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, P.Z.; Cousins, A.B. Carbon isotopes and water use efficiency in C4 plants. Current Opin. Plant Biol. 2016, 31, 155–161. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Percent change of net photosynthesis (Pn), stomatal conductance (Gs), transpiration (Tr), and water-use efficiency (WUE) under three different moderators: photosynthetic pathway (C3 and C4 plants), drought stress (mild, moderate, and severe), and plant forms (trees, shrubs, herbs, and lianas). Numbers near the symbols specify the number of data points, and the error bars show the 95% confidence interval (CI).
Figure 1.
Percent change of net photosynthesis (Pn), stomatal conductance (Gs), transpiration (Tr), and water-use efficiency (WUE) under three different moderators: photosynthetic pathway (C3 and C4 plants), drought stress (mild, moderate, and severe), and plant forms (trees, shrubs, herbs, and lianas). Numbers near the symbols specify the number of data points, and the error bars show the 95% confidence interval (CI).

Figure 2.
Comparison of the percent change of net photosynthesis (Pn), stomatal conductance (Gs), transpiration (Tr), and water-use efficiency (WUE) for photosynthesis pathways (C3 and C4 plants) under different drought intensities (mild, moderate, and severe). Numbers near the symbols specify the number of data points, and the error bars show the 95% CI.
Figure 2.
Comparison of the percent change of net photosynthesis (Pn), stomatal conductance (Gs), transpiration (Tr), and water-use efficiency (WUE) for photosynthesis pathways (C3 and C4 plants) under different drought intensities (mild, moderate, and severe). Numbers near the symbols specify the number of data points, and the error bars show the 95% CI.

Figure 3.
Percent changes of net photosynthesis (Pn), stomatal conductance (Gs), transpiration (Tr), and water-use efficiency (WUE) for herb life forms (A-herbs and P-herbs) and tree foliage types (conifer, deciduous, and evergreen). Numbers near the symbols specify the number of data points, and the error bars show the 95% CI.
Figure 3.
Percent changes of net photosynthesis (Pn), stomatal conductance (Gs), transpiration (Tr), and water-use efficiency (WUE) for herb life forms (A-herbs and P-herbs) and tree foliage types (conifer, deciduous, and evergreen). Numbers near the symbols specify the number of data points, and the error bars show the 95% CI.


{kind=link}
{kind=link}
{kind=link}
Table 1.
The relationships between the Gs response ratio and other factors (Pn, Tr, WUE).
| Moderators | Fitting | R2 | P | N |
|---|---|---|---|---|
| All species | Pn = 0.6846Gs − 0.1801 | 0.5087 | <0.01 | 333 |
| Tr = 0.7937Gs − 0.0862 | 0.7206 | <0.01 | 361 | |
| WUE = − 0.2354Gs − 0.1115 | 0.1615 | <0.01 | 350 | |
| C3 plants | Pn = 0.7378Gs − 0.1482 | 0.4968 | <0.01 | 319 |
| Tr = 0.8249Gs − 0.0650 | 0.7794 | <0.01 | 347 | |
| WUE = − 0.0858Gs − 0.0788 | 0.0104 | 0.12 | 347 | |
| C4 plants | Pn = 0.7744Gs − 0.0326 | 0.7545 | <0.01 | 25 |
| Tr = 0.6699Gs − 0.1391 | 0.7657 | <0.01 | 25 | |
| WUE = − 0.1723Gs − 0.0016 | 0.1547 | 0.16 | 14 | |
| Trees | Pn = 0.8067Gs − 0.0904 | 0.4921 | <0.01 | 132 |
| Tr = 0.9004Gs + 0.0258 | 0.9048 | <0.01 | 132 | |
| WUE = − 0.1819Gs − 0.2457 | 0.0349 | 0.08 | 130 | |
| Shrubs | Pn = 0.6377Gs − 0.4584 | 0.5330 | <0.01 | 43 |
| Tr = 0.6527Gs − 0.4181 | 0.5309 | <0.01 | 43 | |
| WUE = 0.1487Gs − 0.0104 | 0.1367 | 0.78 | 40 | |
| Herbs | Pn = 0.6819Gs − 0.1525 | 0.5046 | <0.01 | 132 |
| Tr = 0.6297Gs − 0.1453 | 0.4829 | <0.01 | 132 | |
| WUE = − 0.3579Gs − 0.1480 | 0.2379 | <0.01 | 116 | |
| Lianas | Pn = 0.4848Gs − 0.0795 | 0.7207 | <0.01 | 24 |
| Tr = 0.8601Gs + 0.0006 | 0.8737 | <0.01 | 24 | |
| WUE = − 0.4282Gs − 0.1900 | 0.527 | <0.01 | 41 |
Gs: Stomatal conductance; Pn: Net photosynthesis; Tr: Transpiration rate; WUE: Water-use efficiency; N: Number of samples.
Table 2.
Pearson correlation coefficients between the Pn, Tr, WUE response ratios and other factors.
Table 2.
Pearson correlation coefficients between the Pn, Tr, WUE response ratios and other factors.
| Categorical Type | Ta | Af | CO2C | RH | Tl | PAR | VPD | |
|---|---|---|---|---|---|---|---|---|
| All plants | Pn | 0.19* | 0.11 | 0.07 | 0.07 | 0.09 | 0.11 | 0.05 |
| Tr | 0.21 | −0.46** | 0.08 | 0.23* | 0.14 | 0.10 | 0.01 | |
| Gs | 0.08 | −0.22 | 0.26** | 0.38** | 0.24* | 0.09 | 0.10 | |
| WUE | 0.09 | 0.38 ** | −0.14* | −0.31** | −0.19 | 0.09 | 0.09 | |
| N | 219 | 70 | 309 | 125 | 140 | 394 | 83 |
Ta: air temperature; Af: air flow; CO2C: CO2 concentration; RH: air relative humidity; Tl: leaf temperature; PAR: photosynthetically active radiation; VPD: leaf to air vapor pressure deficit; N: number of samples; * < 0.05; ** < 0.01.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, J.; Jiang, H.; Song, X.; Jin, J.; Zhang, X. The Responses of Plant Leaf CO2/H2O Exchange and Water Use Efficiency to Drought: A Meta-Analysis. Sustainability 2018, 10, 551. https://0-doi-org.brum.beds.ac.uk/10.3390/su10020551
AMA Style
Zhang J, Jiang H, Song X, Jin J, Zhang X. The Responses of Plant Leaf CO2/H2O Exchange and Water Use Efficiency to Drought: A Meta-Analysis. Sustainability. 2018; 10(2):551. https://0-doi-org.brum.beds.ac.uk/10.3390/su10020551
Chicago/Turabian StyleZhang, Jinmeng, Hong Jiang, Xinzhang Song, Jiaxin Jin, and Xiuying Zhang. 2018. "The Responses of Plant Leaf CO2/H2O Exchange and Water Use Efficiency to Drought: A Meta-Analysis" Sustainability 10, no. 2: 551. https://0-doi-org.brum.beds.ac.uk/10.3390/su10020551
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.