Impacts of Dryland Farm Management Systems on Weeds and Ground Beetles (Carabidae) in the Northern Great Plains



Department of Land Resources and Environmental Sciences, Montana State University, P.O. Box 173120, Bozeman, MT 59717-3120, USA
*
Author to whom correspondence should be addressed.
Sustainability 2018, 10(7), 2146; https://0-doi-org.brum.beds.ac.uk/10.3390/su10072146
Submission received: 22 May 2018
/
Revised: 16 June 2018
/
Accepted: 20 June 2018
/
Published: 23 June 2018
(This article belongs to the Special Issue Sustainable Crop Production Systems)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Farming systems act as ecological filters impacting cropland associated biodiversity; however, the extent of these effects is largely unknown in the drylands of the Northern Great Plains (NGP), a key conventional and organic crop production region. In this 3-year (2013–2015) on-farm study, we addressed this knowledge gap by assessing the effects of conventional and organic farming systems on weed and ground beetle (carabidae) communities. We observed 25 weed species in conventional and 44 in organic fields. Weed species richness and weed species evenness were 237% and 137% greater in organic fields than in conventional fields. We collected a total of 1520 beetle specimens, representing 28 species in conventional and 37 in organic fields. Beetle activity-density and species richness were 220% and 156% greater in organic fields than in conventional fields. Both weed and ground beetle communities differed between conventional and organic fields, indicating that farming systems act as distinct ecological filters. We conclude that, in the drylands of the NGP, adoption of organic farming enhances weed abundance which could have potential management challenges, but provide floral resources to the pollinators and parasitoids. Similarly, greater diversity of ground beetles could enhance conservation biological control of crop pests, ultimately supporting for sustainable agriculture.
1. Introduction
Through various ecological processes, associated biodiversity in croplands could reduce crop yield or facilitate the provision of ecosystem services such as food, fiber, energy, air and water purification, climate regulation, nutrient cycling, pollination, and biological control of pests [1,2]. Nevertheless, there is a global decline in cropland-associated biodiversity which has been attributed to the simplification of crop rotations and the heavy reliance of industrialized crop production systems on off-farm chemical and mechanical inputs to control pest populations, maintain soil fertility, and secure yields [3,4].
Two particularly important groups of associated biodiversity in croplands are weeds and ground beetles (Coleoptera: Carabidae; a.k.a. carabid beetles). Weeds are often considered impediments to agricultural production due to their impacts on crop yields and interference with farming practices [5]. However, many weed species can also provide valuable ecosystem services including increasing soil organic matter, facilitating nutrient cycling, serving as propagule pools of arbuscular mycorrhizal fungi and symbiotic bacteria such as Rhizobia [6], and providing food and habitat for pollinators [7] and natural enemies [8,9]. Ground beetles are one of the most widespread, abundant, and diverse groups of insects. Because most ground beetles are generalist predators of a variety of crop pests including aphids, fly maggots, grubs, and slugs, and most species in the Harpalini and Zabrini tribes are seed predators [10,11], they can help regulate weed and insect pest populations [12,13]. Hence, it is particularly important to study how the farm management systems affect the two contrasting groups of associated biodiversity: weeds and ground beetles.
The Northern Great Plains (NGP) is an important dryland region for the production of conventional and organic small grain, pulse, hay, and oilseed crops [14,15], yet there is limited information regarding the impacts of agricultural management systems on the associated biodiversity, particularly ground beetles and their association with weeds [16]. The majority of chemical fallowed conventional row crop production (hereafter, conventional system) in this region uses synthetic inputs including herbicides and chemical fertilizers, and usually follows a simple crop-fallow rotation to control weeds, enhance soil fertility, and retain water moisture, respectively. On the other hand, organic tilled row crop production (hereafter, organic system) uses cover cropping, crop rotation, and tillage to manage soil fertility and pests. By relying on different suites of management tactics, conventional and organic systems may act as distinct ecological filters of weeds, arthropods, and other members of associated cropland biodiversity, selectively favoring some taxa while excluding others [17,18]. Yet, the extent of these effects is largely unknown in the dryland sections of the NGP. Hence, to assess how the impact to which conventional and organic management systems act as differential ecological filters of weed and ground beetle communities, we conducted a 3-year on-farm study in the NGP. The systems approach employed in our on-farm study precludes us from testing the specific mechanisms responsible for the observed results. Nevertheless, our study highlights the resulting impacts on the associated biodiversity and allow us to speculate on possible causes for the observed results. Hence, we hypothesized that (i) because they use different farm inputs, a conventional system would have different weed and ground beetle communities compared to an organic system, and (ii) an organic system, which does not use herbicides but practices tillage, would have greater abundance and diversity of weeds and ground beetles.
2. Materials and Methods
2.1. Site Description and Cropping History
We conducted a 3-year on-farm study comparing weeds and ground beetle communities in conventional and organic small grain production systems in Big Sandy, Montana, USA (Figure S1; 48.036° N, 110.014° W; Altitude 960 m). Big Sandy is located in Montana’s Golden Triangle, an important dryland wheat producing region in the Northern Great Plains, and primarily composed of small grain producing agricultural lands [15]. The 94-year mean annual precipitation for Big Sandy is 325 mm, and the mean annual minimum and maximum temperatures are −1.2 °C and 14.8 °C, respectively (Table S1). Big Sandy is underlain with a Telstad-Joplin loam (fine loamy, mixed, Aridic Argiboroll) with a pH of 7.9–8.2 [15].
Each year between 2013 and 2015, we selected three conventional fields with spring wheat production and three adjacent organic fields with Kamut wheat [Triticum turanicum Jakubz. (var. Kamut), a close relative of durum wheat] production (see details in [19]). Field sizes ranged from 25 ha to 70 ha and individual fields were separated by a minimum of 500 m. Organic fields had been certified by the Organic Crop Improvement Association (OCIA) International since 1989 and by the United States Department of Agriculture (USDA) organic certificate standards since 2003 [19]). All conventional fields followed a 2-year winter wheat-chemical no-till fallow rotation and all organic fields followed a multi-year continuous rotation of crops or cover crops (Table S2). In the conventional fields, the no-till fallow phase was chemically treated with glyphosate (Roundup®) at rates ranging from 1121 to 1682 g of active ingredient ha−1, depending on the farmer (Table S3). In organic fields, crop residues and cover crops were incorporated by chisel and/or disk tilling, whereas a disk plow was used as needed to manage weeds. Specific agronomic management practices, such as cultivation, crop rotation, fertilizer and pesticide applications, and tillage were at the grower’s discretion (Table S3), and insecticide use was absent in all conventional fields.
2.2. Weed and Ground Beetle Community Sampling
To compare the impacts of agricultural management systems on weed and ground beetle communities, each year, we established a randomly oriented 55 m transect within each field, located at least 150 m from any field border. To evaluate weed communities, we constructed three orthogonal 25 m sub-transects at random locations of the main 55 m transect, with one transect facing the opposite direction of the other two (Figure S2). On 17–18 June, 15–16 July, and 6–7 August in 2013; 23–24 June, 21–22 July, and 6–7 August in 2014; 16–17 June, 13–14 July, and 3–4 August in 2015, we visually quantified weeds based on their estimated percent cover by species of all individuals rooted within a 0.5 m × 1 m (n = 450 frames/sampling) quadrat at every meter interval along each sub-transect.
We estimated ground beetle activity density and community structure using pitfall traps, the most common method of passive sampling for ground dwelling insects [20]. Three pitfall traps were established at three regularly spaced subsampling locations along the main transect of each sampled field (n = 9 traps/field/sampling). One subsampling location was located at each end of the main transect and a third one at its center (Figure S1). To construct the pitfall traps, we dug a 10 cm × 12 cm hole using a post-hole auger, placed two stacked 0.5 L plastic cups (Solo Cup Co., Lake Forest, IL, USA) flush with the soil at ground surface, and filled approximately 25% of the top cup with a propylene glycol-based antifreeze (Arctic Ban Antifreeze, Camco Manufacturing, Greensboro, NC, USA) used as a killing agent and preservative. The pitfall traps (n = 54 traps/sampling) were open for a 48-h interval and the beetle samples in traps were collected on 19 June, 17 July, and 8 August in 2013; 25 June, 23 July, and 8 August in 2014; 18 June, 15 July, and 5 August in 2015. We transferred all ground beetles caught in the pitfall traps to plastic bags (Whirl-Pak®, Nasco Inc., Fort Atkinson, WI, USA) and stored them in the freezer until further processing. Frozen beetle samples were then cleaned in the lab, preserved in 70% ethyl alcohol, and identified to the species following [21]. A specimen of Agonum could not be positively identified to a species and so was recorded as morpho-species. Nomenclature for all species follows Bousquet [22].
2.3. Data Analysis
Metrics of weed and ground beetle community structures included abundance (measured as percentage cover for weeds and activity density for beetles), species richness, α-diversity, and evenness. We calculated species richness (S) as the total number of species present in each field and α-diversity using Simpson’s Diversity Index (1–D), for both groups:
where pi is the proportion of individuals of ith species in a community and S is the total number of species in that community [23]. Similarly, for both weeds and ground beetles, species evenness (J′) was calculated using Pielou’s evenness [24]:
where H′ is Shannon-Wiener diversity index, Hmax = log(S), and S is species richness. Shannon-Wiener diversity index is calculated as where pi is the proportion of each species [25].
After determining whether response variables were strongly correlated (rp > 0.6), we compared the metrics of weed and ground beetle diversity between conventional and organic agroecosystems and through time using linear and generalized linear mixed effects models with a random intercept for each year and each field. Agricultural management system was treated as fixed effects and year, fields, and sampling dates were treated as random effects, with fields nested within years and dates nested within fields. We did not include the seasonal (June–August) peak activity for ground beetles in our paper, as the activity was not different between the two systems.
Temporal differences in community structure of weeds and ground beetles were assessed using non-metric multidimensional scaling (NMDS) ordination and Permutational Multivariate Analysis of Variance (PERMANOVA) on a Bray-Curtis dissimilarity matrix. Pairwise dissimilarity was computed using the Bray-Curtis index:
where BCjk is the dissimilarity between communities j and k, aij and aik are the abundances of species i in sites j and k, respectively, and S is the combined total number of species in either weeds or beetle communities [26,27].
3. Results
3.1. Weed Communities
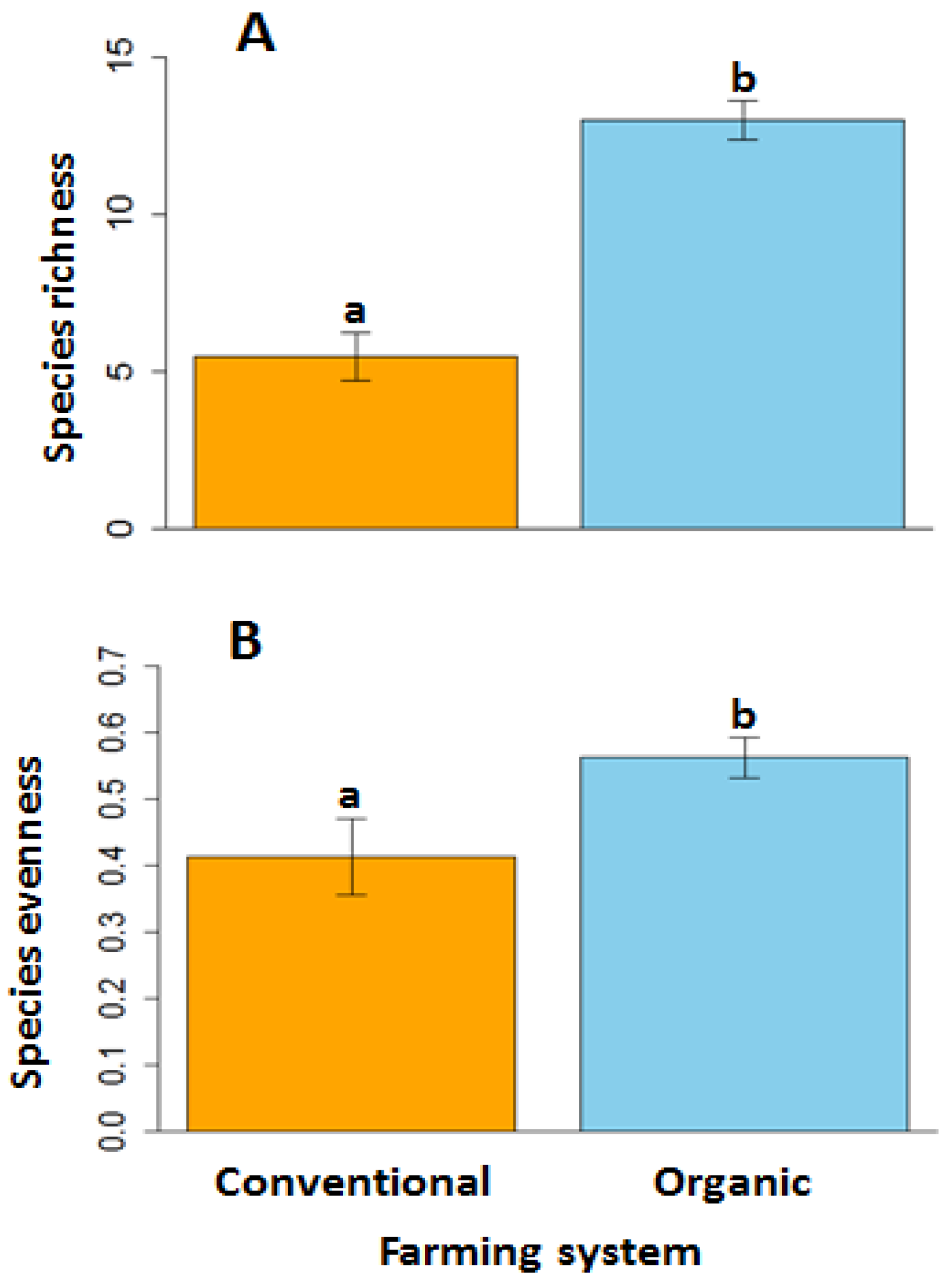
Over the 3-year period of our study, we observed a total of 48 weed species, 10 of which were grasses and 38 of which were forbs. Twenty-five of those species (nine grasses and 16 forbs) occurred in the conventional fields and 44 species (seven grasses and 37 forbs) occurred in the organic fields (Table S4). The five most abundant weed species in conventional systems were Bromus tectorum (8% cover of all species observed), Hordeum jubatum (5%), Avena fatua (5%), Salsola kali (3%), and Cirsium arvense (3%) and in organic systems were Salsola kali (36% cover of all species observed), Chenopodium album (7%), Polygonum convolvulus (7%), Avena fatua (5%), and Medicago sativa (4%). Mean weed species richness was 237% greater in organic fields than in conventional fields (Figure 1A; F = 22.1; df = 1, 14; p ≤ 0.001). Similarly, mean weed species evenness was 137% greater in organic fields than in conventional fields (Figure 1B; F = 6.6; df = 1, 14; p = 0.02).
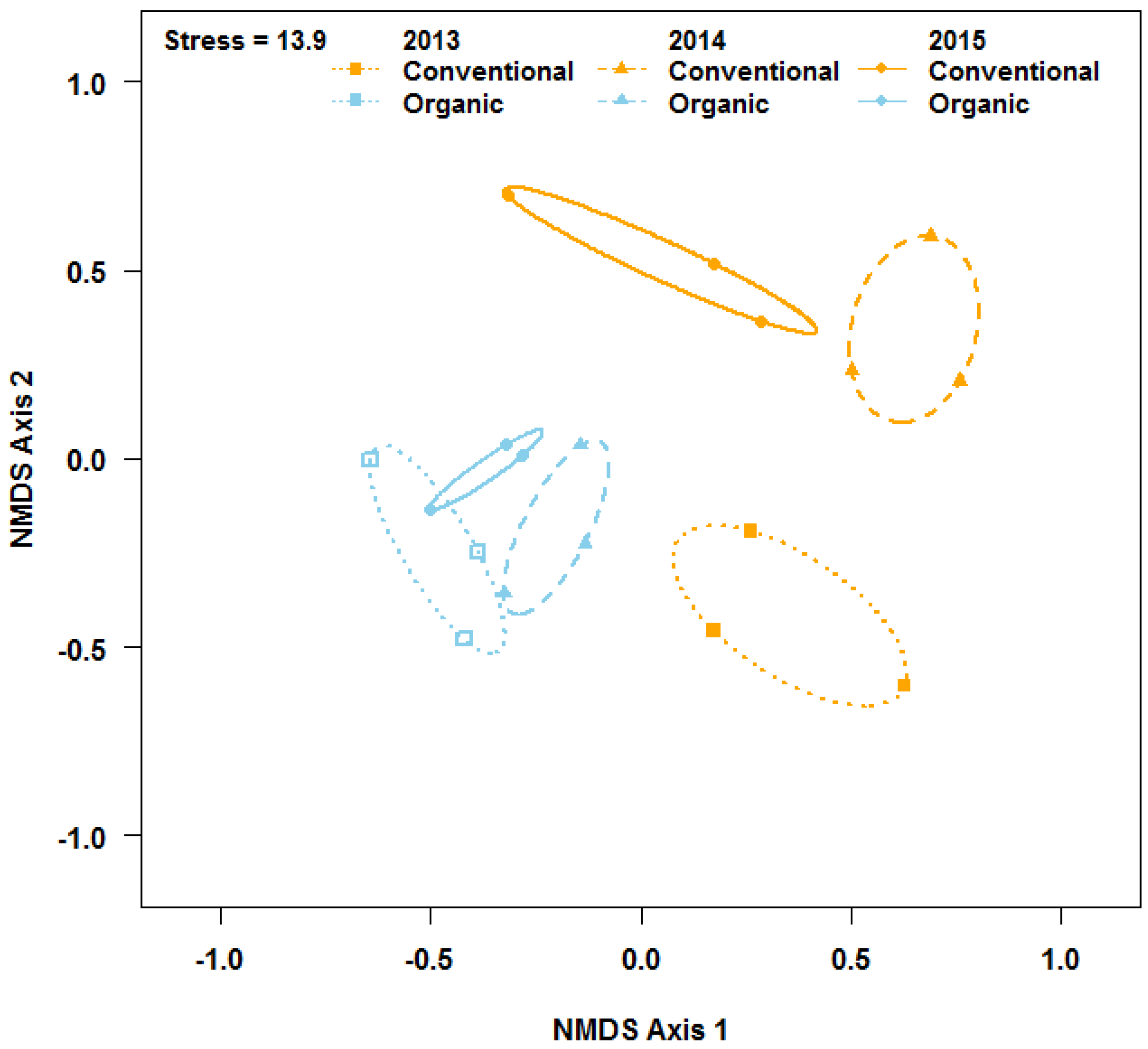
Weed community composition differed between conventional and organic fields (Figure 2; pseudo-F = 6.5; df = 1, 16, R2 = 0.29; p = 0.001), but the effects also depended on the year (pseudo-F = 2.75; df = 2, 12; R2 = 0.17; p = 0.02). Thlaspi arvense, Salsola kali, Carthamus tinctorius, Polygonum convolvulus, Avena fatua, Pisum sativum, Bromus tectorum, Chenopodium album, Medicago sativa, Amaranthus retroflexus, and Hordeum jubatum contributed 73% to the observed dissimilarity in weed community composition between the two farming systems (Figure S3). Common grasses such as B. tectorum and H. jubatum, were more associated with conventional fields, whereas forbs such as S. kali, T. arvense, P. convolvulus, and C. tinctorius were more associated with organic fields (Table S4; Figure S2). Despite the differences in weed community composition between the two farming systems, indicator species analysis did not reveal any key weed species associated either with conventional or organic fields. Also, weed community dispersion was greater across conventional fields than across organic fields (F = 16.7; df = 1, 16; p = 0.002).
3.2. Beetle Communities
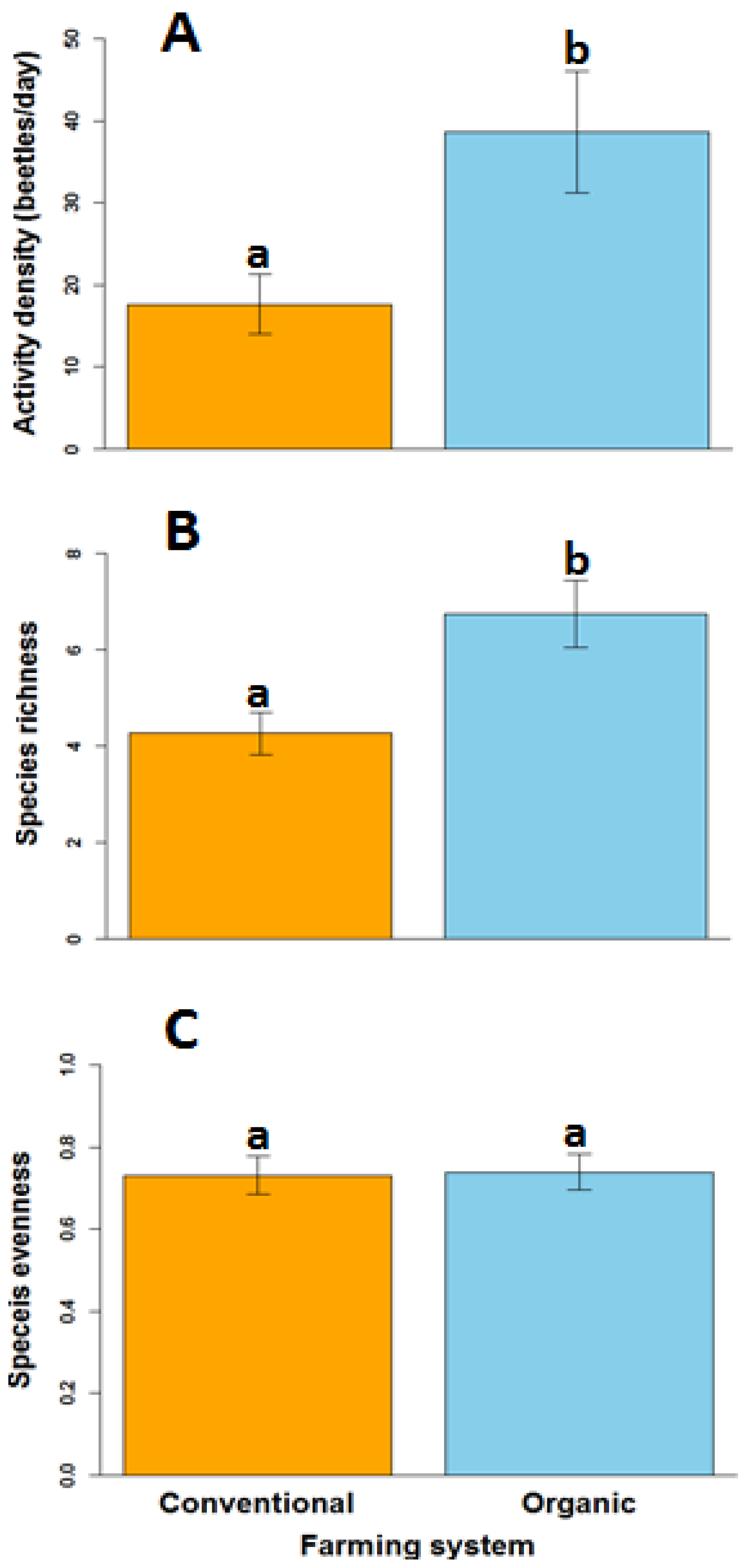
We collected a total of 1520 beetle specimens over the 3-year study period, representing 42 species, 28 of which occurred in conventional and 37 of which occurred in organic fields (Table S5). The five most abundant beetle species in conventional systems were Poecilus scitulus (n = 224; 47% of all beetles captured in conventional fields), Cratacanthus dubius (n = 77; 16%), Pasimachus elongatus (n = 30; 6%), Harpalus amputatus (n = 21; 4%), and Poecilus cursitor (n = 15; 3%), representing about 76% of the total capture. Similarly, the five most abundant species in organic systems were Poecilus scitulus (n = 518; 50% of all beetles captured in organic fields), Harpalus fraternus (n = 85; 8%), Cratacanthus dubius (n = 64; 6%), Poecilus corvus (n = 61; 6%), and Pasimachus elongatus (n = 56; 5%), representing about 75% of the total capture. We detected differences in ground beetle activity density and diversity between conventional and organic systems. Mean ground beetle activity density (beetles per sampling period) was 220% greater in organic systems than in conventional systems (Figure 3A; F = 12.38; df = 1, 14; p = 0.003). Similarly, mean ground beetle species richness was 156% greater in organic systems than in conventional systems (Figure 3B; F = 9.6; df = 1, 14; p ≤ 0.01). However, we did not detect any differences in evenness between organic and conventional systems (Figure 3C; F = 0.02; df = 1, 14; p = 0.9).
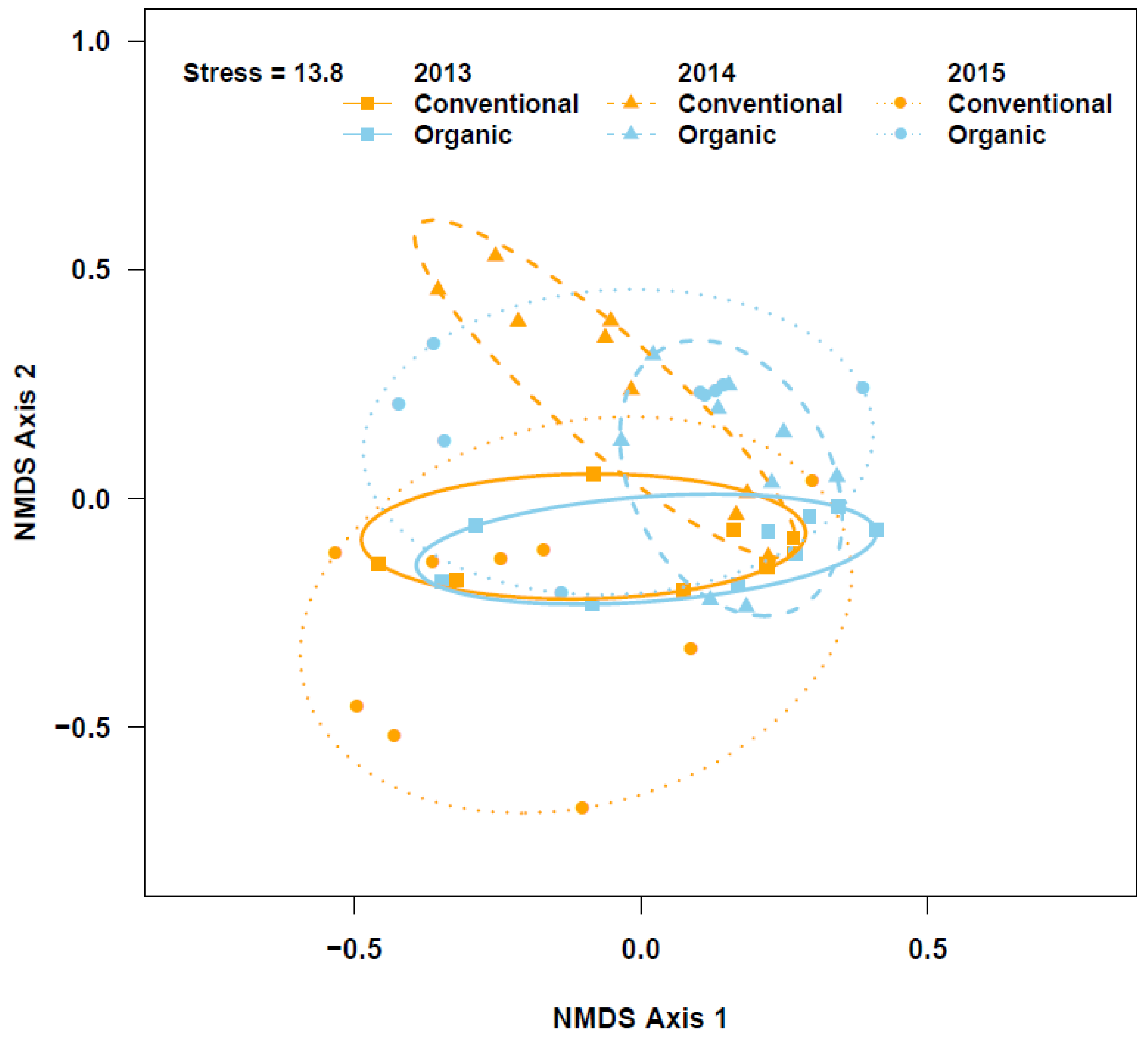
The PERMANOVA, based on activity density and species composition, indicated that the community composition of ground beetle differed between the farming systems (pseudo-F = 1.8; df = 1, 52, R2 = 0.03; p = 0.02), though the beetle communities seemingly overlapped in the two systems (Figure 4). Poecilus scitulus, C. dubius, H. amputatus, P. elongatus, P. corvus, Calosoma luxatum, Amara quenseli, Cicindela punctulata, and H. fraternus contributed 74% of the total observed dissimilarity in beetle community composition between the two farming systems. Like the weed community data, ground beetle community dispersion was also greater across conventional fields than across organic fields (Figure 4; F = 5.7; df = 1, 52; p = 0.02). Also, indicator species analysis revealed that H. fraternus preferentially occurred in organic systems (INDVAL = 0.54, p = 0.02), but we found no beetle indicative of conventional systems.
4. Discussion
This study indicated that organically- and conventionally-managed wheat fields in the drylands of the NGP act as different ecological filters of two contrasting components of the associated biodiversity: weeds and ground beetles. Interestingly, despite the biological and ecological differences between weed and ground beetles, these two groups of the associated biodiversity responded similarly to changes in farming systems. Specifically, we observed greater weed species richness, weed evenness, ground beetle activity density, and ground beetle species richness in organic fields compared with those in conventional fields. Also, both weed and ground beetle community composition differed between the two systems.
When evaluating weed communities, our results are consistent with those of Pollnac et al. [33], who found that organic small grain fields in the NGP had greater weed species richness and evenness than conventional small grain fields which, in turn, could potentially support beneficial insects such as pollinators, parasitoids, and generalists predators [6,19,34,35]. However, organic farms in the drylands of NGP may face exacerbated challenges in weed management [36,37], as the weeds are often considered impediments to agricultural production [5].
The differences in weed communities between the two studied farming systems could have resulted from the impact of different ecological filters imposed by the farm managers, such as herbicide use and simplified rotations in conventional fields, and tillage and increased crop rotation in organic fields [38]. Specifically, the frequent use of selective herbicides, such as auxin regulating in conventional small grain fields in the NGP would exclude dicotyledonous species [39], while selecting for monocotyledonous species. For example, we observed greater association of grasses such as B. tectorum and H. jubatum in conventional fields than in organic fields. Because tillage is frequently used in organic fields, the weed communities in those systems could shift to exclude perennial grasses and favor annual forbs, as frequent soil disturbance kills perennation structures and moves seeds to more surficial layers of the soil [40,41]. However, we also observed few of the perennial weeds such as C. arvense in our organic fields, possibly suggesting an ineffectiveness of tillage to control this species, but to break its roots and further increase its abundance and spread through vegetative propagation [36].
Consistent with Menalled et al. [42], we found greater temporal variability in weed community composition in conventional fields than in organic fields. This higher year-to-year variation in weed community composition in conventional fields could be due to the inconsistencies among different conventional growers in weed control success.
To our knowledge, this is the first study evaluating the impacts of conventional and organic row crop production systems on ground beetle community structure from the dryland section of the NGP. In agreement with previous studies conducted in temperate agricultural systems [12,43,44], we observed that ground beetle communities in the NGP were dominated by a small subset of species from the commonly occurring genera in the Northern Hemisphere including Amara, Agonum, Harpalus, Poecilus, and Pterostichus. In contradiction to previous studies [45,46,47], we found a greater activity density and diversity of ground beetles in tilled organic fields than in no-till conventional fields. The greater activity density and diversity of ground beetles in organic fields possibly enhances the biological control of weed seeds [48,49] and insect pests [50,51]. Compared to no-till conventional fields where crop residues may impede carabid movement; tillage in organic fields may affect the carabid activity positively by reducing crop residues and hence their capture rates could be higher [52]. Bengtsson et al. [9] and Ryan et al. [53] also found a greater abundance of weed seeds, insect pests and, concomitantly, more carabid beetles in organic systems compared with conventional systems. By contrast, previous studies have suggested that increased soil disturbance due to the fact that tillage in organic fields may reduce ground beetle habitat suitability [47] and bury weed seeds, thereby excluding seed-preying beetles [54], and increasing the beetle larvae and adult mortality [55]. Assessing the mechanisms driving farming systems impacts on ground beetles was beyond the scope of our study, but it could be a valuable topic for further investigation to better understand their population dynamics and functional role in the dryland agroecosystems.
Ground beetle community structure differed between the two studied farming systems, suggesting that in the NGP, no-till conventional and tilled organic small grain systems act as distinct ecological filters of beetle biodiversity. Specifically, agricultural management practices such as tillage, cover cropping, crop rotation, and herbicide use that could have altered the habitat suitability or imposed mortality for ground beetles asymmetrically between conventional systems and organic systems [56,57]. Additionally, as in weed communities, we found greater temporal variability in ground beetle community composition in conventional fields than in organic fields. Previous studies have suggested that the weed community composition could be an effective predictor of the beetle community composition [58], but we need further studies to test if the temporal variability in weed communities is directly associated with that of the beetle communities.
Our results showed that H. fraternus preferentially occurred in tilled organic fields but was very rare in no-till conventional fields. A detailed description of the ecology of H. fraternus is lacking, so it is not clear why this species was more abundant and strongly associated with organic fields. Native to North America, H. fraternus is widely distributed across the northwest United States [59] and has been reported to prefer dry, open, and sandy soils [21]; the type of soil observed in both conventional and organic fields [59]. Several other Harpalus species, including H. pensylvanicus, H. herbivagus, and H. affinis are known as seed predators, at least in their adult stages [60,61]. Hence, it is possible that, due to its seed consuming habit, H. fraternus was strongly associated with organic fields that have a more abundant seedbank than conventional fields [62]. In addition, some studies have reported H. fraternus to consume darkling beetles (Eleodes extricata (Say) and E. hispilabris (Say); Coleoptera: Tenebrionidae) and grasshoppers [10,63], but the currently known traits of H. fraternus are not sufficient to explain why it was strongly associated with organic farming, and future studies are required to explore the habitat requirements and food preferences of this species.
5. Conclusions
Our study demonstrated that farming systems can similarly impact two contrasting components of the associated biodiversity: weeds and carabid beetles. Changes in weed community due to the ecological filters of farm management systems may impose bottom-up impacts that could alter insect communities [64,65]. Alternatively, generalist seed predators like ground beetles are impacted by management systems and, in turn, could impose top down impacts on weed communities [10]. Overall, our study showed an increased abundance of weeds and ground beetles in organically managed wheat fields and posed an interesting question for future studies that these two contrasting components of associated biodiversity could be directly (positively or negatively) associated with each other [66]. Additionally, a greater activity density and diversity of carabid beetles in organic fields than in conventional fields may provide increased biological control of weed seeds in these systems [47,67].
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2071-1050/10/7/2146/s1. Table S1: Precipitation and temperature data for Big Sandy, MT, USA; Table S2: Crop rotation in conventional (C) and organic (O) spring wheat fields between 2013 and 2015. All fields were sampled in ‘Year 5’ of the crop phases; Table S3: Agronomic management details for conventional and organic spring wheat fields between 2013 and 2015; Table S4: Complete list of weeds (grasses* and forbs) with their relative proportions observed in organic and conventional wheat fields between 2013 and 2015; Table S5: Complete list of ground beetle species and their activity densities at organic and conventional fields, Big Sandy, MT between 2013 and 2015; Figure S1: Study area map showing nine conventional and nine organic fields across three years in Big Sandy, MT, United States; Figure S2: Schematic diagram of one of the fields in Big Sandy, MT, United States used to assess weed and ground beetle communities in conventional and organic cropping systems. The solid black line at the center of field represents the main transect (55m), three orthogonal dotted lines represent sub-transects (25m) used to assed weed communities, and black circles represent pitfall traps used for beetle sampling (A). An example of pitfall traps (B); Figure S3: Principal component analysis plot showing the associations of different weed species with conventional or organic systems.
Author Contributions
Conceptualization, S.A. and F.D.M.; Formal analysis, S.A.; Funding acquisition, F.D.M.; Methodology, S.A.; Project administration, S.A. and F.D.M.; Supervision, F.D.M.; Writing—original draft, S.A.; Writing—review & editing, F.D.M.
Funding
This research was supported by the United States Department of Agriculture National Institute of Food and Agriculture [grant numbers MONB00314 and MONB00128].
Acknowledgments
We thank Sean McKenzie, Stephen Johnson, Nar Ranabhat, Madison Nixon, Ali Thornton, Sam Leuthold, Andrew Thorson and Wyatt. Holmes for their assistance in the field and laboratory.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Quijas, S.; Schmid, B.; Balvanera, P. Plant diversity enhances provision of ecosystem services: A new synthesis. Basic Appl. Ecol. 2010, 11, 582–593. [Google Scholar] [CrossRef] [Green Version]
- Vandermeer, J.; Lawrence, D.; Symstad, A. Effect of biodiversity on ecosystem functioning in managed ecosystems. In Biodiversity and Ecosystem Functioning; Loreau, M., Naeem, S., Inchausti, P., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 157–168. [Google Scholar]
- Duru, M.; Therond, O.; Martin, G.; Martin-Clouaire, R.; Magne, M.A.; Justes, E.; Journet, E.P.; Aubertot, J.N.; Savary, S.; Bergez, J.E.; et al. How to implement biodiversity-based agriculture to enhance ecosystem services: A review. Agron. Sustain. Dev. 2015, 35, 1259–1281. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260. [Google Scholar] [CrossRef] [PubMed]
- DeDecker, J.J.; Masiunas, J.B.; Davis, A.S.; Flint, C.G. Weed management practice selection among midwest U.S. organic growers. Weed Sci. 2014, 62, 520–531. [Google Scholar] [CrossRef]
- Jordan, N.; Vatovec, C. Agroecological benefits from weeds. In Weed Biology and Management; Springer: Dordrecht, The Netherlands, 2004; pp. 137–158. [Google Scholar]
- Carvalheiro, L.G.; Veldtman, R.; Shenkute, A.G.; Tesfay, G.B.; Pirk, C.W.W.; Donaldson, J.S.; Nicolson, S.W. Natural and within-farmland biodiversity enhances crop productivity. Ecol. Lett. 2011, 14, 251–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siemann, E. Experimental tests of effects of plant productivity and diversity on grassland arthropod diversity. Ecology 1998, 79, 2057–2070. [Google Scholar] [CrossRef]
- Bengtsson, J.; Ahnstrom, J.; Weibull, A.C. The effects of organic agriculture on biodiversity and abundance: A meta-analysis. J. Appl. Ecol. 2005, 42, 261–269. [Google Scholar] [CrossRef]
- Larochelle, A. The food of carabid beetles (Coleoptera: Carabidae, including Cicindelidae). In Fabreries Supplement; Association des entomologistes amateurs du Québec: Varennes, QC, France, 1990; Volume 5, pp. 1–132. [Google Scholar]
- Sunderland, K.D. Invertebrate Pest Control by Carabids; Holland, J.M., Ed.; Intercept Limited: Anover, UK, 2002. [Google Scholar]
- Gaines, H.R.; Gratton, C. Seed predation increases with ground beetle diversity in a Wisconsin (USA) potato agroecosystem. Agric. Ecosyst. Environ. 2010, 137, 329–336. [Google Scholar] [CrossRef]
- Honek, A.; Martinkova, Z.; Jarosik, V. Ground beetles (Carabidae) as seed predators. Eur. J. Entomol. 2003, 100, 531–544. [Google Scholar] [CrossRef] [Green Version]
- Johnston, A.M.; Tanaka, D.L.; Miller, P.R.; Brandt, S.A.; Nielsen, D.C.; Lafond, G.P.; Riveland, N.R. Oilseed crops for semiarid cropping systems in the Northern Great Plains. Agron. J. 2002, 94, 231–240. [Google Scholar] [CrossRef]
- Miller, P.R.; Lighthiser, E.J.; Jones, C.A.; Holmes, J.A.; Rick, T.L.; Wraith, J.M. Pea green manure management affects organic winter wheat yield and quality in semiarid Montana. Can. J. Plant Sci. 2011, 91, 497–508. [Google Scholar] [CrossRef]
- Hoekman, D.; LeVan, K.E.; Ball, G.E.; Browne, R.A.; Davidson, R.L.; Erwin, T.L.; Knisley, C.B.; LaBonte, J.R.; Lundgren, J.; Maddison, D.R.; et al. Design for ground beetle abundance and diversity sampling within the National Ecological Observatory Network. Ecosphere 2017, 8, e01744. [Google Scholar] [CrossRef] [Green Version]
- Funk, J.L.; Cleland, E.E.; Suding, K.N.; Zavaleta, E.S. Restoration through reassembly: Plant traits and invasion resistance. Trends Ecol. Evol. 2008, 23, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Keddy, P.A. Assembly and response rules: Two goals for predictive community ecology. J. Veg. Sci. 1992, 3, 157–164. [Google Scholar] [CrossRef]
- Adhikari, S.; Seipel, T.; Menalled, F.D.; Weaver, D.K. Farming system and wheat cultivar affect infestation of and parasitism on Cephus cinctus in the Northern Great Plains. Pest Manag. Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kotze, D.J.; Brandmayr, P.; Casale, A.; Dauffy-Richard, E.; Dekoninck, W.; Koivula, M.J.; Lövei, G.L.; Mossakowski, D.; Noordijk, J.; Paarmann, W.; et al. Forty years of carabid beetle research in Europe—From taxonomy, biology, ecology and population studies to bioindication, habitat assessment and conservation. Zookeys 2011, 100, 55–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindroth, C.H. The ground beetles (Carabidae, excl. Cicindelinae) of Canada and Alaska, Part 1–6. Opusc. Entomol. Entomologiska Siillskapet, Lund, Sweden. 1969, 1–1192. Available online: http://www.librarything.com/work/10804214 (accessed on 16 June 2018).
- Bousquet, Y. Catalogue of Geadephaga (Coleoptera, Adephaga) of America, north of Mexico. Zookeys 2012, 245. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Pielou, E.C. Species-diversity and pattern-diversity in the study of ecological succession. J. Theor. Biol. 1966, 10, 370–383. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution and measurement of species diversity. Taxon 1972, 21, 213. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B. Overview of community matrices. Anal. Ecol. Communities 2002, 28, 3–12. [Google Scholar]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; ISBN 3-900051-07-0. Available online: https://stat.ethz.ch/pipermail/r-help/2008-May/161481.html (accessed on 29 October 2017).
- Roberts, D.W. Labdsv: Ordination and Multivariate Analysis for Ecology. R Package Version 1.8-0. Available online: https://CRAN.R-project.org/package=labdsv (accessed on 20 December 2016).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.4-4. Available online: https://CRAN.R-project.org/package=vegan (accessed on 5 January 2017).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Morales, M. Package ‘sciplot’: Scientific graphing functions for factorial designs. 2017. Available online: https://cran.r-project.org/web/packages/sciplot/sciplot.pdf (accessed on 16 June 2018).
- Pollnac, F.W.; Rew, L.J.; Maxwell, B.D.; Menalled, F.D. Spatial patterns, species richness and cover in weed communities of organic and conventional no-tillage spring wheat systems. Weed Res. 2008, 48, 398–407. [Google Scholar] [CrossRef]
- Adhikari, S. Impacts of Dryland Farming Systems on Biodiversity, Plant-Insect Interactions, and Ecosystem Services. Ph.D. Thesis, Montana State University, Bozeman, MT, USA, 2018. [Google Scholar]
- DiTommaso, A.; Averill, K.M.; Hoffmann, M.P.; Fuchsberg, J.R.; Losey, J.E. Integrating insect, resistance, and floral resource management in weed control decision-making. Weed Sci. 2016, 64, 743–756. [Google Scholar] [CrossRef]
- Orloff, N.; Mangold, J.; Miller, Z.; Menalled, F. A meta-analysis of field bindweed (Convolvulus arvensis L.) and Canada thistle (Cirsium arvense L.) management in organic agricultural systems. Agric. Ecosyst. Environ. 2018, 254, 264–272. [Google Scholar] [CrossRef]
- Tautges, N.E.; Goldberger, J.R.; Burke, I.C. A survey of weed management in organic small grains and forage systems in the Northwest United States. Weed Sci. 2016, 64, 513–522. [Google Scholar] [CrossRef]
- Smith, R.G.; Atwood, L.W.; Pollnac, F.W.; Warren, N.D. Cover-crop species as distinct biotic filters in weed community assembly. Weed Sci. 2015, 63, 282–295. [Google Scholar] [CrossRef]
- Grossmann, K. Auxin herbicides: Current status of mechanism and mode of action. Pest Manag. Sci. 2009, 66. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.G.; Derksen, D.A.; Blackshaw, R.E.; Van Acker, R.C.; Légère, A.; Watson, P.R.; Turnbull, G.C. Symposium: A multistudy approach to understanding weed population shifts in medium- to long-term tillage systems. Weed Sci. 2004, 52, 874–880. [Google Scholar] [CrossRef]
- Barroso, J.; Miller, Z.J.; Lehnhoff, E.A.; Hatfield, P.G.; Menalled, F.D. Impacts of cropping system and management practices on the assembly of weed communities. Weed Res. 2015, 55, 426–435. [Google Scholar] [CrossRef]
- Menalled, F.D.; Gross, K.L.; Hammond, M. Weed aboveground and seedbank community responses to agricultural management systems. Ecol. Appl. 2001, 11, 1586–1601. [Google Scholar] [CrossRef]
- McKenzie, S.C.; Goosey, H.B.; O’Neill, K.M.; Menalled, F.D. Impact of integrated sheep grazing for cover crop termination on weed and ground beetle (Coleoptera:Carabidae) communities. Agric. Ecosyst. Environ. 2016, 218, 141–149. [Google Scholar] [CrossRef]
- Goosey, H.B.; Mckenzie, S.C.; Rolston, M.G.; O’Neill, K.M.; Menalled, F.D. Impacts of contrasting alfalfa production systems on the drivers of carabid beetle (Coleoptera: Carabidae) community dynamics. Environ. Entomol. 2015, 44, 1052–1064. [Google Scholar] [CrossRef] [PubMed]
- Jonason, D.; Smith, H.G.; Bengtsson, J.; Birkhofer, K. Landscape simplification promotes weed seed predation by carabid beetles (Coleoptera: Carabidae). Landsc. Ecol. 2013, 28, 487–494. [Google Scholar] [CrossRef]
- Melnychuk, N.; Olfert, O.; Youngs, B.; Gillott, C. Abundance and diversity of Carabidae (Coleoptera) in different farming systems. Agric. Ecosyst. Environ. 2003, 95, 69–72. [Google Scholar] [CrossRef]
- Menalled, F.D.; Smith, R.G.; Dauer, J.T.; Fox, T.B. Impact of agricultural management on carabid communities and weed seed predation. Agric. Ecosyst. Environ. 2007, 118, 49–54. [Google Scholar] [CrossRef]
- Diekötter, T.; Wamser, S.; Dörner, T.; Wolters, V.; Birkhofer, K. Organic farming affects the potential of a granivorous carabid beetle to control arable weeds at local and landscape scales. Agric. For. Entomol. 2016, 18, 167–173. [Google Scholar] [CrossRef]
- Kulkarni, S.S.; Dosdall, L.M.; Willenborg, C.J. The role of ground beetles (Coleoptera: Carabidae) in weed seed consumption: A Review. Weed Sci. 2015, 63, 355–376. [Google Scholar] [CrossRef]
- Menalled, F.D.; Lee, J.C.; Landis, D.A. Manipulating carabid beetle abundance alters prey removal rates in corn fields. BioControl 1999, 43, 441–456. [Google Scholar] [CrossRef]
- Williams, I.H.; Ferguson, A.W.; Kruus, M.; Veromann, E.; Warner, D.J. Ground beetles as predators of oilseed rape pests: Incidence, spatio-temporal distributions and feeding. In Biocontrol-Based Integrated Management of Oilseed Rape Pests; Springer: Dordrecht, The Netherlands, 2010; pp. 115–149. [Google Scholar]
- Hatten, T.D.; Bosque-pé Rez, N.A.; Labonte, J.R.; Guy, S.O.; Eigenbrode, S.D. Effects of tillage on the activity density and biological diversity of carabid beetles in spring and winter crops. Environ. Entomol 2007, 36, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.R.; Smith, R.G.; Mirsky, S.B.; Mortensen, D.A.; Seidel, R. Management filters and species traits: Weed community assembly in long-term organic and conventional systems. Weed Sci. 2010, 58, 265–277. [Google Scholar] [CrossRef]
- Kulkarni, S.S.; Dosdall, L.M.; Spence, J.R.; Willenborg, C.J. Depth of seed burial and gender influence weed seed predation by three species of ground beetle (Coleoptera: Carabidae). Weed Sci. 2015, 63, 910–915. [Google Scholar] [CrossRef]
- Blubaugh, C.K.; Kaplan, I. Tillage compromises weed seed predator activity across developmental stages. Biol. Control 2015, 81, 76–82. [Google Scholar] [CrossRef]
- Dritschilo, W.; Wanner, D. Ground beetle abundance in organic and conventional corn fields. Environ. Entomol. 1980, 9, 629–631. [Google Scholar] [CrossRef]
- Ward, M.J.; Ryan, M.R.; Curran, W.S.; Barbercheck, M.E.; Mortensen, D.A. Cover crops and disturbance influence activity-density of weed seed predators Amara aenea and Harpalus pensylvanicus (Coleoptera: Carabidae). Weed Sci. 2011, 59, 76–81. [Google Scholar] [CrossRef]
- Schaffers, A.P.; Raemakers, I.P.; Sýkora, K.V.; ter Braak, C.J.F. Arthropod assemblages are best predicted by plant species composition. Ecology 2008, 89, 782–794. [Google Scholar] [CrossRef] [PubMed]
- Noonan, G.R. Biogeographical Patterns of North American Harpalus Latreille (Insecta: Coleoptera: Carabidae). J. Biogeogr. 1990, 17, 583. [Google Scholar] [CrossRef]
- Barney, R.J.; Pass, B.C. Foraging behavior and feeding preference of ground beetles (Coleoptera: Carabidae) in Kentucky alfalfa. J. Econ. Entomol. 1986, 79, 1334–1337. [Google Scholar] [CrossRef]
- O’Rourke, M.E.; Heggenstaller, A.H.; Liebman, M.; Rice, M.E. Post-dispersal weed seed predation by invertebrates in conventional and low-external-input crop rotation systems. Agric. Ecosyst. Environ. 2006, 116, 280–288. [Google Scholar] [CrossRef]
- Harbuck, K.S.B.; Menalled, F.D.; Pollnac, F.W. Impact of cropping systems on the weed seed banks in the northern Great Plains, USA. Weed Biol. Manag. 2009, 9, 160–168. [Google Scholar] [CrossRef]
- Lavigne, R.J.; Pfadt, R.E. Parasites and predators of Wyoming rangeland: Grasshoppers. Wyo. Agric. Exp. Sta. Sci. Monogr. 1966, 3, 31. [Google Scholar]
- Haddad, N.M.; Crutsinger, G.M.; Gross, K.; Haarstad, J.; Tilman, D. Plant diversity and the stability of foodwebs. Ecol. Lett. 2011, 14, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef]
- Diehl, E.; Wolters, V.; Birkhofer, K. Arable weeds in organically managed wheat fields foster carabid beetles by resource- and structure-mediated effects. Arthropod Plant Interact. 2012, 6, 75–82. [Google Scholar] [CrossRef]
- Bohan, D.A.; Boursault, A.; Brooks, D.R.; Petit, S. National-scale regulation of the weed seedbank by carabid predators. J. Appl. Ecol. 2011, 48, 888–898. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Weed species richness (A) and weed species evenness (B) on conventional and organic farms. Error bars represent the standard errors of the mean, while letters above bars indicate significant differences between systems at p < 0.05.
Figure 1.
Weed species richness (A) and weed species evenness (B) on conventional and organic farms. Error bars represent the standard errors of the mean, while letters above bars indicate significant differences between systems at p < 0.05.

Figure 2.
Non-metric multidimensional scaling (NMDS) ordination of weed communities observed in conventional and organic wheat fields. Dotted ellipses inscribe communities in 2013, dashed ellipses inscribe communities in 2014, and solid ellipses inscribe communities in 2015.
Figure 2.
Non-metric multidimensional scaling (NMDS) ordination of weed communities observed in conventional and organic wheat fields. Dotted ellipses inscribe communities in 2013, dashed ellipses inscribe communities in 2014, and solid ellipses inscribe communities in 2015.

Figure 3.
Ground beetle activity density (A), species richness (B) and species evenness (C) on conventional and organic farms. Error bars represent the standard errors of the mean, while letters above bars indicate significant differences between systems at p < 0.05.
Figure 3.
Ground beetle activity density (A), species richness (B) and species evenness (C) on conventional and organic farms. Error bars represent the standard errors of the mean, while letters above bars indicate significant differences between systems at p < 0.05.

Figure 4.
Non-metric multidimensional scaling (NMDS) ordination of ground beetle communities observed in conventional and organic wheat fields. Solid ellipses inscribe communities in 2013, dashed ellipses inscribe communities in 2014, and dotted ellipses inscribe communities in 2015.
Figure 4.
Non-metric multidimensional scaling (NMDS) ordination of ground beetle communities observed in conventional and organic wheat fields. Solid ellipses inscribe communities in 2013, dashed ellipses inscribe communities in 2014, and dotted ellipses inscribe communities in 2015.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Adhikari, S.; Menalled, F.D. Impacts of Dryland Farm Management Systems on Weeds and Ground Beetles (Carabidae) in the Northern Great Plains. Sustainability 2018, 10, 2146. https://0-doi-org.brum.beds.ac.uk/10.3390/su10072146
AMA Style
Adhikari S, Menalled FD. Impacts of Dryland Farm Management Systems on Weeds and Ground Beetles (Carabidae) in the Northern Great Plains. Sustainability. 2018; 10(7):2146. https://0-doi-org.brum.beds.ac.uk/10.3390/su10072146
Chicago/Turabian StyleAdhikari, Subodh, and Fabian D. Menalled. 2018. "Impacts of Dryland Farm Management Systems on Weeds and Ground Beetles (Carabidae) in the Northern Great Plains" Sustainability 10, no. 7: 2146. https://0-doi-org.brum.beds.ac.uk/10.3390/su10072146
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.