Photofermentative Poly-3-Hydroxybutyrate Production by Rhodopseudomonas sp. S16-VOGS3 in a Novel Outdoor 70-L Photobioreactor

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
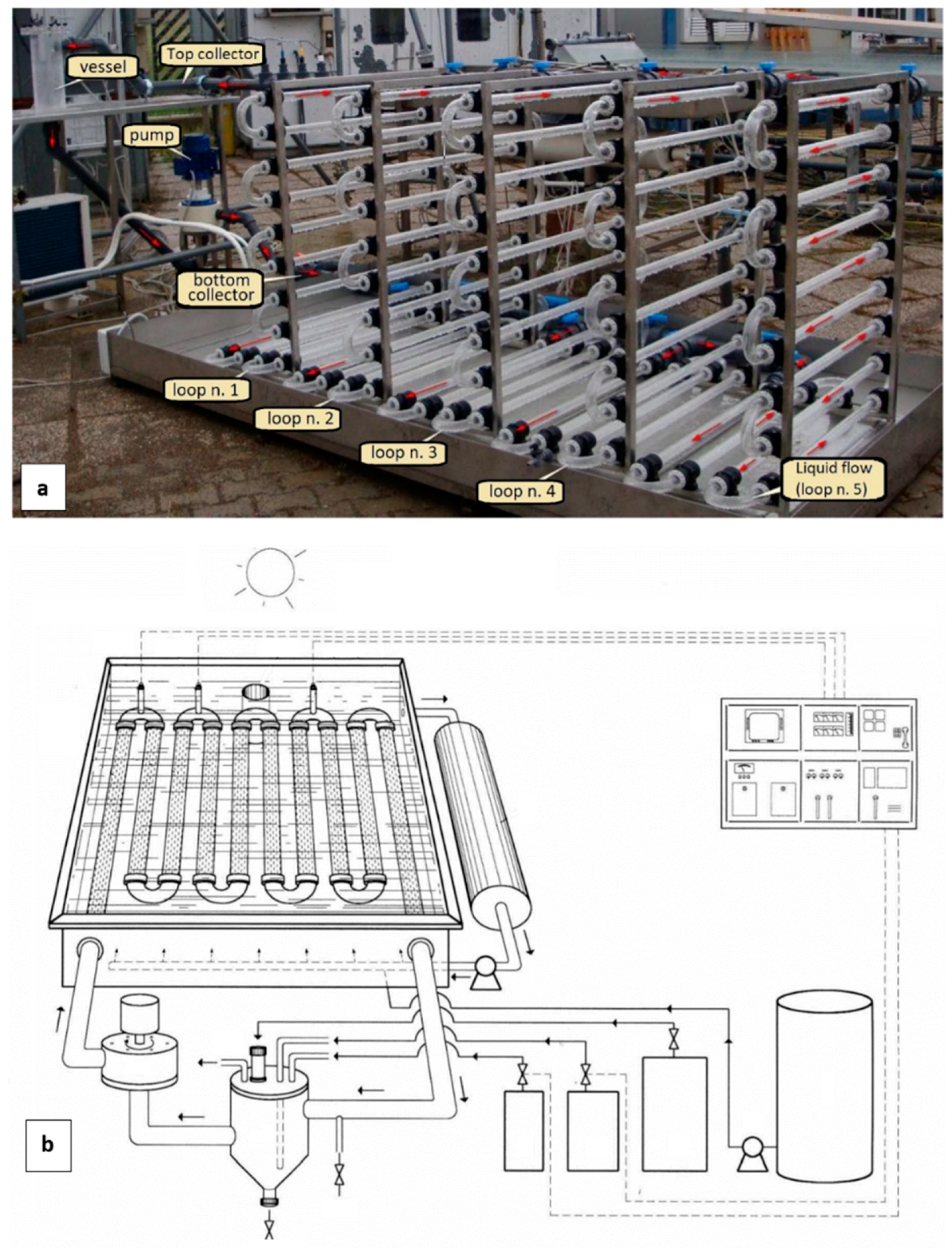
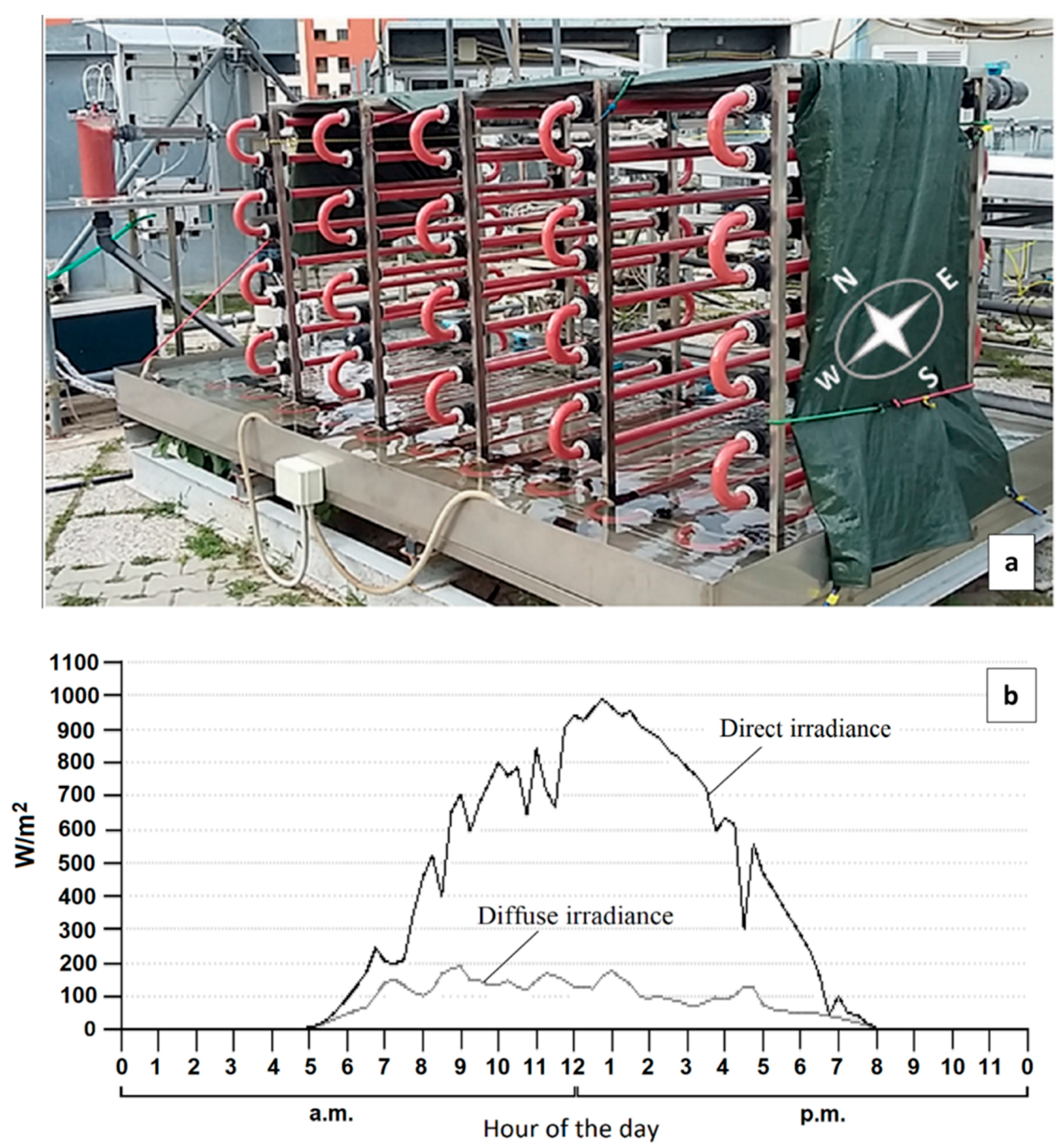
2.1. Photobioreactor
2.2. Microorganism Strain and Culture Conditions
2.3. Outdoor Bacterial Growth
2.4. PHB Recovery
2.5. Analytical Methods
2.6. Thermal Analysis
3. Results and Discussion
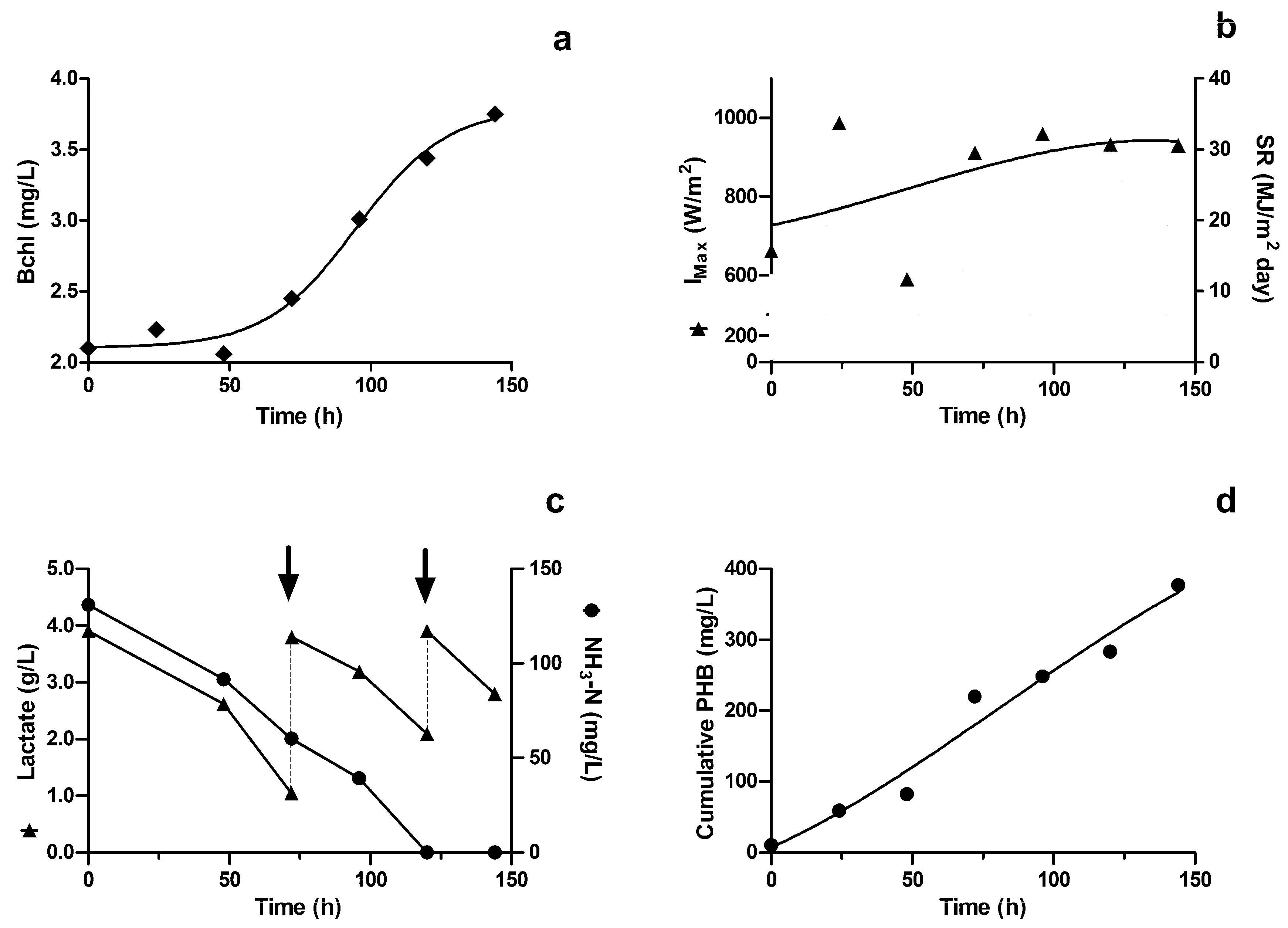
3.1. Photobioreactor Performance
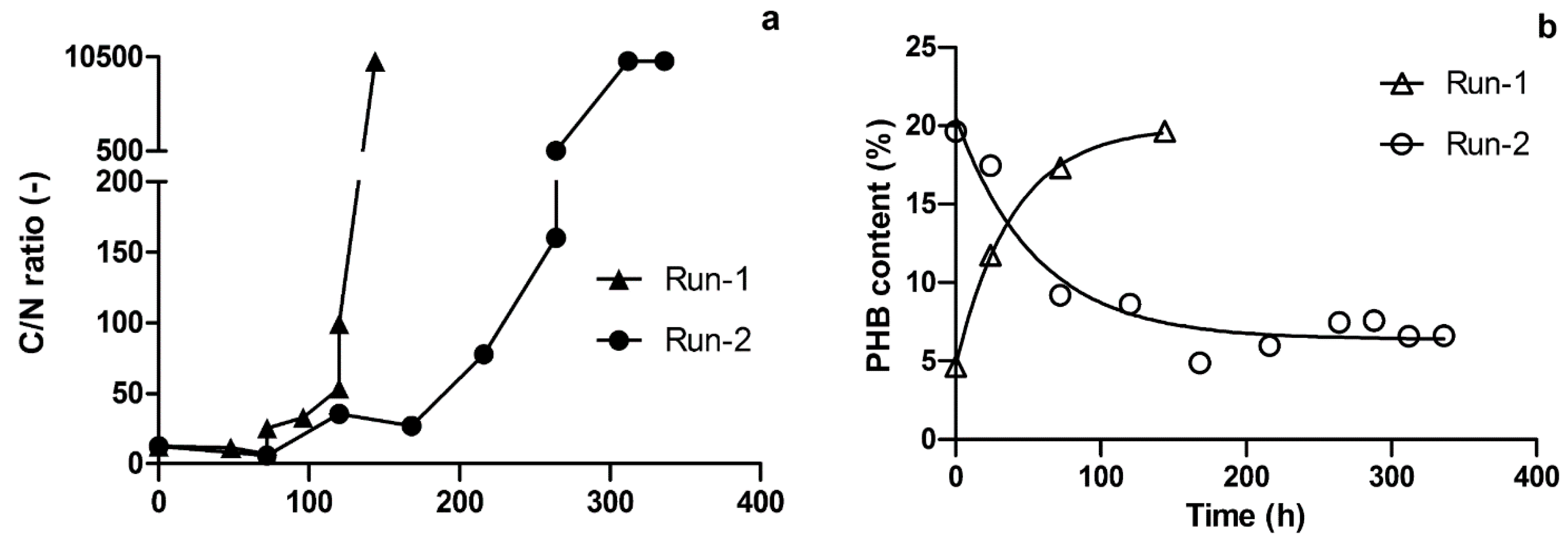
3.2. PHB Production
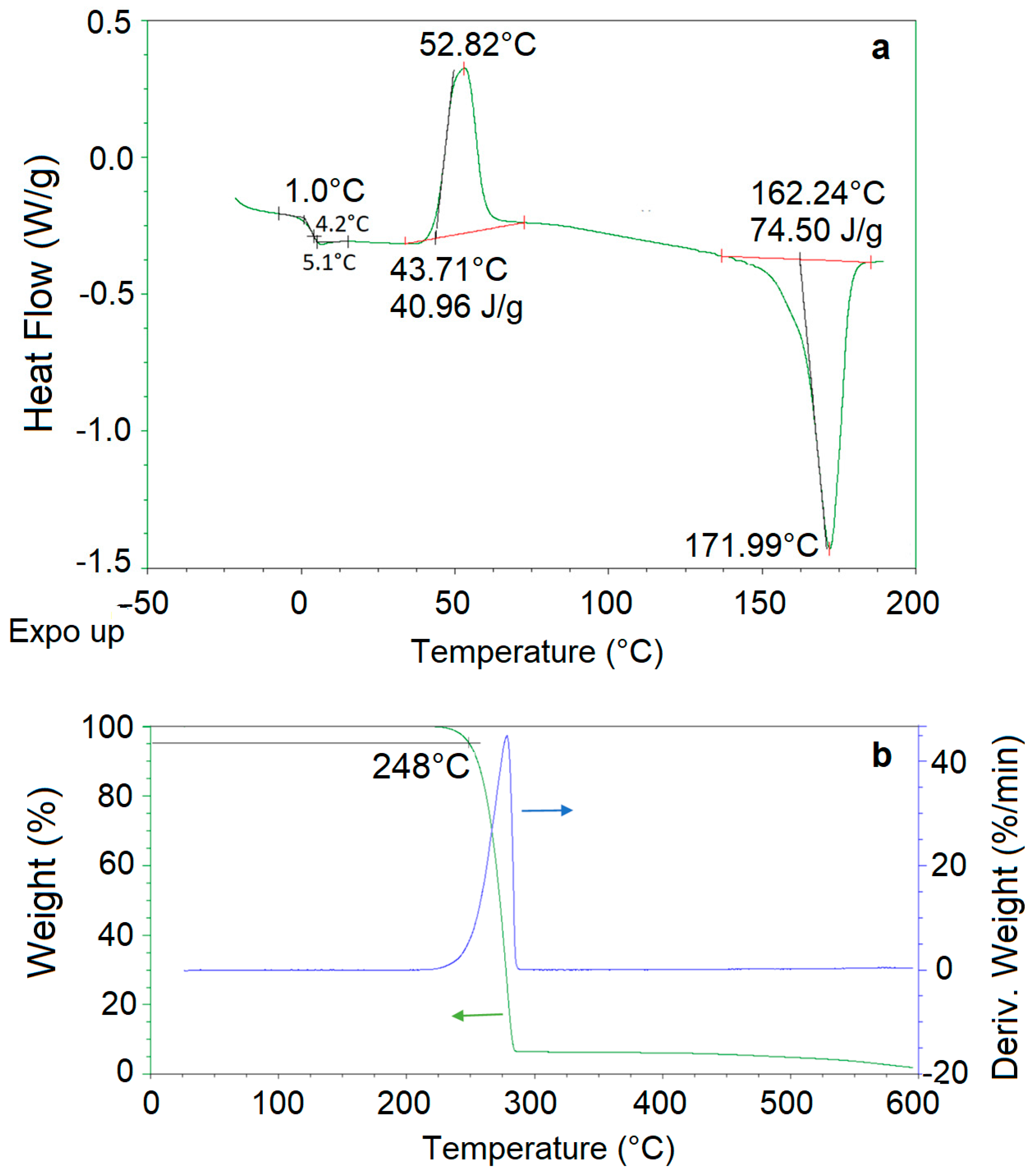
3.3. PHB Thermal Properties and Stability
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- COM (2018) 28. A European Strategy for Plastics in a Circular Economy. Available online: http://ec.europa.eu/environment/circular-economy/pdf/plastics-strategy.pdf (accessed on 2 September 2018).
- Valavanidis, A. Global Plastic Waste and Oceans Pollution Million Tons of Plastic Waste have Gone Missing in the World Oceans? Athens, Greece. May 2016. Available online: https://www.researchgate.net/publication/303549578 (accessed on 26 July 2018).
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic waste inputs from land into the ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Bioplastics Market Data 2017. Global Production Capacities of Bioplastics 2017–2022. REPORT European Bioplastics. Available online: https://www.european-bioplastics.org/market/ (accessed on 26 July 2018).
- Balaji, S.; Gopi, K.; Muthuvelan, B. A review on production of poly β-hydroxybutyrates from cyanobacteria for the production of bio plastics. Algal Res. 2013, 2, 278–285. [Google Scholar] [CrossRef]
- Carpine, R.; Olivieri, G.; Hellingwerf, K.; Pollio, A.; Marzocchella, A. The cyanobacterial route to produce poly-β-hydroxybutyrate. Chem. Eng. Trans. 2015, 43, 289–294. [Google Scholar] [CrossRef]
- Padovani, G.; Carlozzi, P.; Seggiani, M.; Cinelli, P.; Vitolo, S.; Lazzeri, A. PHB-rich biomass and bioH2 production by means of photosynthetic microorganisms. Chem. Eng. Trans. 2016, 49, 55–60. [Google Scholar] [CrossRef]
- Rydz, J.; Sikorska, W.; Kyulavska, M.; Christova, D. Polyester-based (bio) degradable polymers as environmentally friendly materials for sustainable development. Int. J. Mol. Sci. 2015, 16, 564–596. [Google Scholar] [CrossRef] [PubMed]
- Bugnicourt, E.; Cinelli, P.; Lazzeri, A.; Alvarez, V. Polyhydroxyalkanoate (PHA): Review of synthesis, characteristics, processing and potential applications in packaging. Express Polym. Lett. 2014, 8, 791–808. [Google Scholar] [CrossRef] [Green Version]
- Mozumder, M.D.S.I.; Garcia-Gonzalez, L.; De Wever, H.; Volcke, E.I.P. Poly(3-hydroxybutyrate) (PHB) production from CO2: Model development and process optimization. Biochem. Eng. J. 2015, 98, 107–116. [Google Scholar] [CrossRef]
- Jiang, G.; Hill, D.J.; Kowalczuk, M.; Johnston, B.; Adamus, G.; Irorere, V.; Iza Radecka, I. Carbon sources for polyhydroxyalkanoates and an integrated biorefinery. Int. J. Mol. Sci. 2016, 17, 1157. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Tsygankov, A.; Miyake, J.; Tokiwa, Y.; Asa-da, Y. Accumulation of poly-(hydroxybutyrate) by a non-sulfur photosynthetic bacterium, Rhodobacter sphaeroides RV at different pH. Biotechnol. Lett. 1995, 17, 395–400. [Google Scholar] [CrossRef]
- Khatipov, E.; Miyake, M.; Miyake, J.; Asada, Y. Accumulation of poly-β-hydroxybutyrate by Rhodobacter sphaeroides on various carbon and nitrogen substrates. FEMS Microbiol. Lett. 1998, 162, 39–45. [Google Scholar] [CrossRef]
- Kim, B.W.; Chang, K.P.; Chang, H.N. Effect of light source on the microbiological desulfurization in a photobioreactor. Bioprocess. Eng. 1997, 17, 343–348. [Google Scholar] [CrossRef]
- Padovani, G.; Emiliani, G.; Giovanelli, A.; Traversi, M.L.; Carlozzi, P. Assessment of glycerol usage by five different purple non-sulfur bacterial strains for bioplastic production. J. Environ. Chem. Eng. 2018, 6, 616–622. [Google Scholar] [CrossRef]
- Carlozzi, P.; Sacchi, A. Biomass production and studies on Rhodopseudomonas palustris grown in an outdoor, temperature controlled, underwater tubular photobioreactor. J. Biotechnol. 2001, 88, 239–249. [Google Scholar] [CrossRef]
- Carlozzi, P.; Pushparaj, B.; Degl’Innocenti, A.; Capperucci, A. Growth characteristics of Rhodopseudomonas palustris cultured outdoors, in an underwater tubular photobioreactor and investigation on photosynthetic efficiency. Appl. Microbiol. Biot. 2006, 73, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Al-Kaddo, K.B.; Sudesh, K.; Samian, M.R. Screening of bacteria for PHA production using waste glycerol as carbon source and the ability of new strain to produce P(3HB-co-3HV) copolymer. Malaysian J. Microbiol 2016, 12, 245-–253. [Google Scholar] [CrossRef]
- Padovani, G.; Vaičiulytė, S.; Carlozzi, P. BioH2 photoproduction by means of Rhodopseudomonas palustris sp. cultured in a lab-scale photobioreactor operated in batch, fed-batch and semi-continuous modes. Fuel 2016, 166, 203–210. [Google Scholar] [CrossRef]
- Arrieta, M.P.; Samper, M.D.; Lopez, J.; Jiménez, A. Combined Effect of Poly(hydroxybutyrate) and Plasticizers on Polylactic acid Properties for Film Intended for Food Packaging. J. Polym. Environ. 2014, 22, 460–470. [Google Scholar] [CrossRef]
- Carlozzi, P. Dilution of solar radiation through “culture” lamination in photobioreactor rows facing south north: A way to improve the efficiency of light utilization by cyanobacteria (Arthrospira platensis). Biotechnol. Bioeng. 2003, 81, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Hai, T.; Ahlers, H.; Gorenflo, V.; Steinbüchel, A. Axenic cultivation of anoxygenic phototrophic bacteria, cyanobacteria, and microalgae in a new closed tubular glass photobioreactor. Appl. Microbiol. Biot. 2000, 53, 383–389. [Google Scholar] [CrossRef]
- Palamae, S.; Choorit, W.; Dechatiwongse, P.; Zhang, D.; del Rio-Chanona, E.A.; Chisti, Y. Production of renewable biohydrogen by Rhodobacter sphaeroides S10: A comparison of photobioreactors. J. Clean Prod. 2018, 181, 318–328. [Google Scholar] [CrossRef]
- Watanabe, Y.; Saiki, H. Development of a photobioreactor incorporating Chlorella sp. for removal of CO2 in Stack gas. Energy Convers. Manag. 1997, 38, 499–503. [Google Scholar] [CrossRef]
- Khanna, S.; Srivastava, A.K. Optimization of nutrient feed concentration and addition time for production of poly(β-hydroxybutyrate). Enzyme Microb. Tech. 2006, 39, 1145–1151. [Google Scholar] [CrossRef]
- Wang, Y.J.; Hua, F.L.; Tsang, Y.F.; Chan, S.Y.; Sin, S.N.; Chua, H.; Yu, P.H.F.; Ren, N.Q. Synthesis of PHAs from wastes under various C:N ratios. Bioresour. Technol. 2007, 98, 1690–1693. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Kleerebezem, R.; van Loosdrecht, M.C.M. Influence of the C/N ratio on the performance of polyhydroxybutyrate (PHB) producing sequencing batch reactors at short SRTs. Water Res. 2010, 44, 2141–2152. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Jho, E.H.; Nam, K. Effect of C/N ratio on polyhydroxyalkanoates (PHA) accumulation by Cupriavidus necator and its implication on the use of rice straw hydrolysates. Environ. Eng. Res. 2015, 20, 246–253. [Google Scholar] [CrossRef]
- Carpine, R.; Raganati, F.; Olivieri, G.; Hellingwerf, K.J.; Pollio, A.; Salatino, P.; Marzocchella, A. Poly-β-hydroxybutyrate (PHB) production by Synechocystis PCC6803 from CO2: Model development. Algal Res. 2018, 29, 49–60. [Google Scholar] [CrossRef]
- Martino, L.; Cruz, M.V.; Scoma, A.; Freitas, F.; Bertin, L.; Scandola, M.; Reis, M.A.M. Recovery of amorphous polyhydroxybutyrate granules from Cupriavidus necator cells grown on used cooking oil. Int. J. Biol. Macromol. 2014, 71, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Altaee, N.; Fahdila, A.; Yousif, E.; Sudesh, K. Recovery and subsequent characterization of polyhydroxybutyrate from Rhodococcus equi cells grown on crude palm kernel oil. J. Taibah Univ. Sci. 2016, 10, 543–550. [Google Scholar] [CrossRef]
- Sindhu, R.; Ammu, B.; Binod, P.; Deepthi, S.K.; Ramachandran, K.B.; Soccol, C.R.; Pandey, A. Production and characterization of poly-3-hydroxybutyrate from crude glycerol by Bacillus sphaericus NII 0838 and improving its thermal properties by blending with other polymers. Braz. Arch. Biol. Technol. 2011, 54, 783–794. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| di (cm) | Working Volume (L) | Si (m2) | Si/V Ratio (m−1) | Ref. |
|---|---|---|---|---|
| 8.0 | 80.0 | 3.200 | 40.0 | [22] |
| 4.8 | 53.0 | 3.014 | 56.9 | [17] |
| 2.6 | 70.0 | 6.300 | 90.0 | This study |
| 2.2 | 1.0 | 0.182 | 181.7 | [23] |
| 1.6 | 22.1 | 2.200 | 99.5 | [24] |
| 1.0 | 11.0 | 4.400 | 400.0 | [21] |
| Microorganism | Biopolymer | Tc (°C) | Tg (°C) | Tm (°C) | ΔHm (J/g) | xc (%) | T (°C) | Ref. |
|---|---|---|---|---|---|---|---|---|
| Rhodopseudomonas sp. S16-VOGS3 | PHB | 52.8 | 4.2 | 172 | 90.7 | 51 | 248 | This study |
| Rhodococcus equi | PHB | nd* | 2.8 | 173 | 74.5 | 62 | 276 | [30] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carlozzi, P.; Seggiani, M.; Cinelli, P.; Mallegni, N.; Lazzeri, A. Photofermentative Poly-3-Hydroxybutyrate Production by Rhodopseudomonas sp. S16-VOGS3 in a Novel Outdoor 70-L Photobioreactor. Sustainability 2018, 10, 3133. https://0-doi-org.brum.beds.ac.uk/10.3390/su10093133
Carlozzi P, Seggiani M, Cinelli P, Mallegni N, Lazzeri A. Photofermentative Poly-3-Hydroxybutyrate Production by Rhodopseudomonas sp. S16-VOGS3 in a Novel Outdoor 70-L Photobioreactor. Sustainability. 2018; 10(9):3133. https://0-doi-org.brum.beds.ac.uk/10.3390/su10093133
Chicago/Turabian StyleCarlozzi, Pietro, Maurizia Seggiani, Patrizia Cinelli, Norma Mallegni, and Andrea Lazzeri. 2018. "Photofermentative Poly-3-Hydroxybutyrate Production by Rhodopseudomonas sp. S16-VOGS3 in a Novel Outdoor 70-L Photobioreactor" Sustainability 10, no. 9: 3133. https://0-doi-org.brum.beds.ac.uk/10.3390/su10093133