Diversity of Trichoderma spp. in Marine Environments and Their Biological Potential for Sustainable Industrial Applications

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Trichoderma Cultures
2.2. Phylogenetic Analysis
2.3. Preparation of Fungal Extracts
2.4. Measurement of Antioxidant Activity by ABTS Scavenging Ability
2.5. Measurement of Antioxidant Activity by DPPH Radical Scavenging Ability
2.6. Measurement of Antifungal Activities
2.7. Tyrosinase Inhibition Activity
3. Results and Discussion
3.1. Identification and Phylogeny
3.2. Antioxidant Activity
3.3. Antifungal Activity
3.4. Tyrosinase Inhibition Activity
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Freguia, S.; Logrieco, M.E.; Monetti, J.; Ledezma, P.; Virdis, B.; Tsujimura, S. Self-powered bioelectrochemical nutrient recovery for fertilizer generation from human urine. Sustainability 2019, 11, 5490. [Google Scholar] [CrossRef] [Green Version]
- Chou, J.-S.; Yu, C.-P.; Truong, D.-N.; Susilo, B.; Hu, A.; Sun, Q. Predicting microbial species in a river based on physicochemical properties by bio-inspired metaheuristic optimized machine learning. Sustainability 2019, 11, 6889. [Google Scholar] [CrossRef] [Green Version]
- Vieira, T.A.; Panagopoulos, T. Urban forestry in brazilian amazonia. Sustainability 2020, 12, 3235. [Google Scholar] [CrossRef] [Green Version]
- Heo, Y.M.; Lee, H.; Kim, K.; Kwon, S.L.; Park, M.Y.; Kang, J.E.; Kim, G.-H.; Kim, B.S.; Kim, J.-J. Fungal diversity in intertidal mudflats and abandoned solar salterns as a source for biological resources. Mar. Drugs 2019, 17, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Lee, D.W.; Kwon, S.L.; Heo, Y.M.; Jang, S.; Kwon, B.-O.; Khim, J.S.; Kim, G.-H.; Kim, J.-J. Importance of functional diversity in assessing the recovery of the microbial community after the hebei spirit oil spill in korea. Environ. Int. 2019, 128, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Heo, Y.M.; Kwon, S.L.; Yoo, Y.; Lee, A.H.; Kwon, B.-O.; Kim, G.-H.; Khim, J.S.; Kim, J.-J. Recovery of the benthic bacterial community in coastal abandoned saltern requires over 35 years: A comparative case study in the yellow sea. Environ. Int. 2020, 135, 105412. [Google Scholar] [CrossRef]
- Romano, G.; Costantini, M.; Sansone, C.; Lauritano, C.; Ruocco, N.; Ianora, A. Marine microorganisms as a promising and sustainable source of bioactive molecules. Mar. Environ. Res. 2017, 128, 58–69. [Google Scholar] [CrossRef]
- Tourova, T.; Sokolova, D.; Nazina, T.; Grouzdev, D.; Kurshev, E.; Laptev, A. Biodiversity of microorganisms colonizing the surface of polystyrene samples exposed to different aqueous environments. Sustainability 2020, 12, 3624. [Google Scholar] [CrossRef]
- Ge, S.; Ma, J.; Liu, L.; Yuan, Z. The impact of exogenous aerobic bacteria on sustainable methane production associated with municipal solid waste biodegradation: Revealed by high-throughput sequencing. Sustainability 2020, 12, 1815. [Google Scholar] [CrossRef] [Green Version]
- Han, S.-I.; Jeon, M.S.; Heo, Y.M.; Kim, S.; Choi, Y.-E. Effect of pseudoalteromonas sp. Mebic 03485 on biomass production and sulfated polysaccharide biosynthesis in porphyridium cruentum utex 161. Bioresour. Technol. 2020, 302, 122791. [Google Scholar] [CrossRef]
- Ham, J.; Lim, W.; Kim, K.; Heo, Y.M.; Ryu, S.M.; Lee, D.; Kim, J.-J.; Song, G. Gentisyl alcohol inhibits proliferation and induces apoptosis via mitochondrial dysfunction and regulation of mapk and pi3k/akt pathways in epithelial ovarian cancer cells. Mar. Drugs 2019, 17, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, Y.M.; Lee, H.; Lee, C.; Kang, J.; Ahn, J.-W.; Lee, Y.M.; Kang, K.-Y.; Choi, Y.-E.; Kim, J.-J. An integrative process for obtaining lipids and glucose from chlorella vulgaris biomass with a single treatment of cell disruption. Algal Res.-Biomass Biofuels Bioprod. 2017, 27, 286–294. [Google Scholar] [CrossRef]
- Heo, Y.M.; Kim, K.; Kwon, S.L.; Na, J.; Lee, H.; Jang, S.; Kim, C.H.; Jung, J.; Kim, J.-J. Investigation of filamentous fungi producing safe, functional water-soluble pigments. Mycobiology 2018, 46, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Lee, Y.M.; Heo, Y.M.; Lee, J.; Kim, J.S.; Kang, K.Y.; Kim, J.-J. Utilization of agricultural residues for enhancement of cellulolytic enzyme production and enzymatic saccharification by trichoderma harzianum kuc1716. Ind. Crops Prod. 2017, 109, 185–191. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod. 2004, 67, 1216–1238. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.; Adholeya, A.; Barrow, C.J.; Deshmukh, S.K. Marine fungi: An untapped bioresource for future cosmeceuticals. Phytochem. Lett. 2018, 23, 15–20. [Google Scholar] [CrossRef]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69. [Google Scholar] [CrossRef]
- Troussellier, M.; Escalas, A.; Bouvier, T.; Mouillot, D. Sustaining rare marine microorganisms: Macroorganisms as repositories and dispersal agents of microbial diversity. Front. Microbiol. 2017, 8, 947. [Google Scholar] [CrossRef]
- Kong, Q. Marine microorganisms as biocontrol agents against fungal phytopathogens and mycotoxins. Biocontrol Sci. Technol. 2018, 28, 77–93. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749. [Google Scholar] [CrossRef] [PubMed]
- Sivasithamparam, K.; Ghisalberti, E. Secondary metabolism in trichoderma. Trichoderma Gliocladium. Vol. 1 Basic Biol. Taxon. Genet. 2014, 1, 139. [Google Scholar]
- Lu, Z.; Tombolini, R.; Woo, S.; Zeilinger, S.; Lorito, M.; Jansson, J.K. In vivo study of trichoderma-pathogen-plant interactions, using constitutive and inducible green fluorescent protein reporter systems. Appl. Environ. Microbiol. 2004, 70, 3073–3081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reino, J.L.; Guerrero, R.F.; Hernández-Galán, R.; Collado, I.G. Secondary metabolites from species of the biocontrol agent trichoderma. Phytochem. Rev. 2008, 7, 89–123. [Google Scholar] [CrossRef]
- Hong, J.-H.; Lee, J.; Min, M.; Ryu, S.-M.; Lee, D.; Kim, G.-H.; Kim, J.-J. 6-pentyl-α-pyrone as an anti-sapstain compound produced by trichoderma gamsii kuc1747 inhibits the germination of ophiostomatoid fungi. Holzforschung 2014, 68, 769–774. [Google Scholar] [CrossRef]
- Song, F.; Dai, H.; Tong, Y.; Ren, B.; Chen, C.; Sun, N.; Liu, X.; Bian, J.; Liu, M.; Gao, H.; et al. Trichodermaketones a−d and 7-o-methylkoninginin d from the marine fungus trichoderma koningii. J. Nat. Prod. 2010, 73, 806–810. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43. [Google Scholar] [CrossRef]
- Bissett, J.; Gams, W.; Jaklitsch, W.; Samuels, G.J. Accepted trichoderma names in the year 2015. IMA Fungus 2015, 6, 263–295. [Google Scholar] [CrossRef] [Green Version]
- Su, D.; Ding, L.; He, S. Marine-derived trichoderma species as a promising source of bioactive secondary metabolites. Mini-Rev. Med. Chem. 2018, 18, 1702–1713. [Google Scholar] [CrossRef]
- Zhang, J.-C.; Chen, G.-Y.; Li, X.-Z.; Hu, M.; Wang, B.-Y.; Ruan, B.-H.; Zhou, H.; Zhao, L.-X.; Zhou, J.; Ding, Z.-T. Phytotoxic, antibacterial, and antioxidant activities of mycotoxins and other metabolites from trichoderma sp. Nat. Prod. Res. 2017, 31, 2745–2752. [Google Scholar] [CrossRef]
- Samson, R.A.; Hoekstra, E.S.; Frisvad, J.C. Introduction to Food-and Airborne Fungi; Centraalbureau Voor Schimmelcultures (CBS): New York, NY, USA, 2004. [Google Scholar]
- Chaverri, P.; Branco-Rocha, F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T.; Samuels, G.J. Systematics of the trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia 2015, 107, 558–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lind, A.L.; Wisecaver, J.H.; Lameiras, C.; Wiemann, P.; Palmer, J.M.; Keller, N.P.; Rodrigues, F.; Goldman, G.H.; Rokas, A. Drivers of genetic diversity in secondary metabolic gene clusters within a fungal species. PLoS Biol. 2017, 15, e2003583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuels, G.J.; Dodd, S.L.; Gams, W.; Castlebury, L.A.; Petrini, O. Trichoderma species associated with the green mold epidemic of commercially grown agaricus bisporus. Mycologia 2002, 94, 146–170. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal rna genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear ribosomal internal transcribed spacer (its) region as a universal DNA barcode marker for fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. Mafft multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Maddison, D.R.; Maddison, W.P. Macclade 4; Sinauer: Sunderland, MA, USA, 2000. [Google Scholar]
- Swofford, D.L. Phylogenetic Analysis Using Parsimony; Illinois Natural History Survey: Champaign, IL, USA, 1993. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide-sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Hafidh, R.R.; Abdulamir, A.S.; Vern, L.S.; Bakar, F.A.; Abas, F.; Jahanshiri, F.; Sekawi, Z. Inhibition of growth of highly resistant bacterial and fungal pathogens by a natural product. Open Microbiol. J. 2011, 5, 96. [Google Scholar] [CrossRef]
- Lai, H.Y.; Lim, Y.Y.; Tan, S.P. Antioxidative, tyrosinase inhibiting and antibacterial activities of leaf extracts from medicinal ferns. Biosci. Biotechnol. Biochem. 2009, 73, 1362–1366. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhuang, W. Trichoderma (hypocrea) species with green ascospores from china. Pers. Mol. Phylogeny Evol. Fungi 2015, 34, 113. [Google Scholar] [CrossRef] [Green Version]
- Samuels, G.J.; Ismaiel, A.; Mulaw, T.B.; Szakacs, G.; Druzhinina, I.S.; Kubicek, C.P.; Jaklitsch, W.M. The longibrachiatum clade of trichoderma: A revision with new species. Fungal Divers. 2012, 55, 77–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval-Denis, M.; Sutton, D.A.; Cano-Lira, J.F.; Gené, J.; Fothergill, A.W.; Wiederhold, N.P.; Guarro, J. Phylogeny of the clinically relevant species of the emerging fungus trichoderma and their antifungal susceptibilities. J. Clin. Microbiol. 2014, 52, 2112–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaklitsch, W.; Samuels, G.; Ismaiel, A.; Voglmayr, H. Disentangling the trichoderma viridescens complex. Pers. Mol. Phylogeny Evol. Fungi 2013, 31, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaklitsch, W.M. European species of hypocrea part ii: Species with hyaline ascospores. Fungal Divers. 2011, 48, 1–250. [Google Scholar] [CrossRef] [Green Version]
- Park, M.S.; Oh, S.-Y.; Cho, H.J.; Fong, J.J.; Cheon, W.-J.; Lim, Y.W. Trichoderma songyi sp. Nov., a new species associated with the pine mushroom (tricholoma matsutake). Antonie Van Leeuwenhoek 2014, 106, 593–603. [Google Scholar] [CrossRef]
- Ze-Hong, W.; Dong, L.; Ying, X.; Jian-Liang, C.; Wen-Han, L. Antioxidant xanthones and anthraquinones isolated from a marine-derived fungus aspergillus versicolor. Chin. J. Nat. Med. 2018, 16, 219–224. [Google Scholar]
- Gogineni, V.; Hamann, M.T. Marine natural product peptides with therapeutic potential: Chemistry, biosynthesis, and pharmacology. Biochim. Biophys. Acta 2018, 1862, 81–196. [Google Scholar] [CrossRef]
- Douglas, T.E.; Dokupil, A.; Reczyńska, K.; Brackman, G.; Krok-Borkowicz, M.; Keppler, J.K.; Božič, M.; Van Der Voort, P.; Pietryga, K.; Samal, S.K. Enrichment of enzymatically mineralized gellan gum hydrogels with phlorotannin-rich ecklonia cava extract seanol® to endow antibacterial properties and promote mineralization. Biomed. Mater. 2016, 11, 045015. [Google Scholar] [CrossRef]
- Chang, T.-S. An updated review of tyrosinase inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [Green Version]
- Mas-Chamberlin, C.; Mondon, P.; Lamy, F.; Peschard, O.; Lintner, K. Strategies of antiaging actives in sunscreen products. Cosmet. Toilet. 2006, 121, 45. [Google Scholar]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, Y.M.; Kim, K.; Ryu, S.M.; Kwon, S.L.; Park, M.Y.; Kang, J.E.; Hong, J.-H.; Lim, Y.W.; Kim, C.; Kim, B.S.; et al. Diversity and ecology of marine algicolous arthrinium species as a source of bioactive natural products. Mar. Drugs 2018, 16, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, D. 13 mycoses of marine organisms: An overview of pathogenic fungi. In The Biology of Marine Fungi; Cambridge University Press: Cambridge, UK, 1986; ISBN 13: 978-0521308991. [Google Scholar]
- Skaptsov, M.; Smirnov, S.; Kutsev, M.; Uvarova, O.; Sinitsyna, T.; Shmakov, A.; Matsura, A. Antifungal activity of several isolates of trichoderma against cladosporium and botrytis. Ukr. J. Ecol. 2018, 8, 88–91. [Google Scholar] [CrossRef] [Green Version]
- Vinodkumar, S.; Indumathi, T.; Nakkeeran, S. Trichoderma asperellum (nvta2) as a potential antagonist for the management of stem rot in carnation under protected cultivation. Biol. Control 2017, 113, 58–64. [Google Scholar] [CrossRef]
- Wu, Q.; Sun, R.; Ni, M.; Yu, J.; Li, Y.; Yu, C.; Dou, K.; Ren, J.; Chen, J. Identification of a novel fungus, trichoderma asperellum gdfs1009, and comprehensive evaluation of its biocontrol efficacy. PLoS ONE 2017, 12, e0179957. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.S.; Wei, C.I.; Marshall, M.R. Inhibition mechanism of kojic acid on polyphenol oxidase. J. Agric. Food Chem. 1991, 39, 1897–1901. [Google Scholar] [CrossRef]
- May, O.; Moyer, A.; Wells, P.; Herrick, H. The production of kojic acid by aspergillus flavus. J. Am. Chem. Soc. 1931, 53, 774–782. [Google Scholar] [CrossRef]
- Parrish, F.; Wiley, B.; Simmons, E.; Long Jr, L. Production of aflatoxins and kojic acid by species of aspergillus and penicillium. Appl. Microbiol. 1966, 14, 139. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Wu, X.; Sun, M.; Li, M. Two novel tyrosinase inhibitory sesquiterpenes induced by cucl2 from a marine-derived fungus pestalotiopsis sp. Z233. Mar. Drugs 2013, 11, 2713–2721. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, T.; Yamada, K.; Minoura, K.; Miyamoto, K.; Usami, Y.; Kobayashi, T.; Hamada-Sato, N.; Imada, C.; Tsujibo, H. Purification and determination of the chemical structure of the tyrosinase inhibitor produced by trichoderma viride strain h1-7 from a marine environment. Biol. Pharm. Bull. 2008, 31, 1618–1620. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
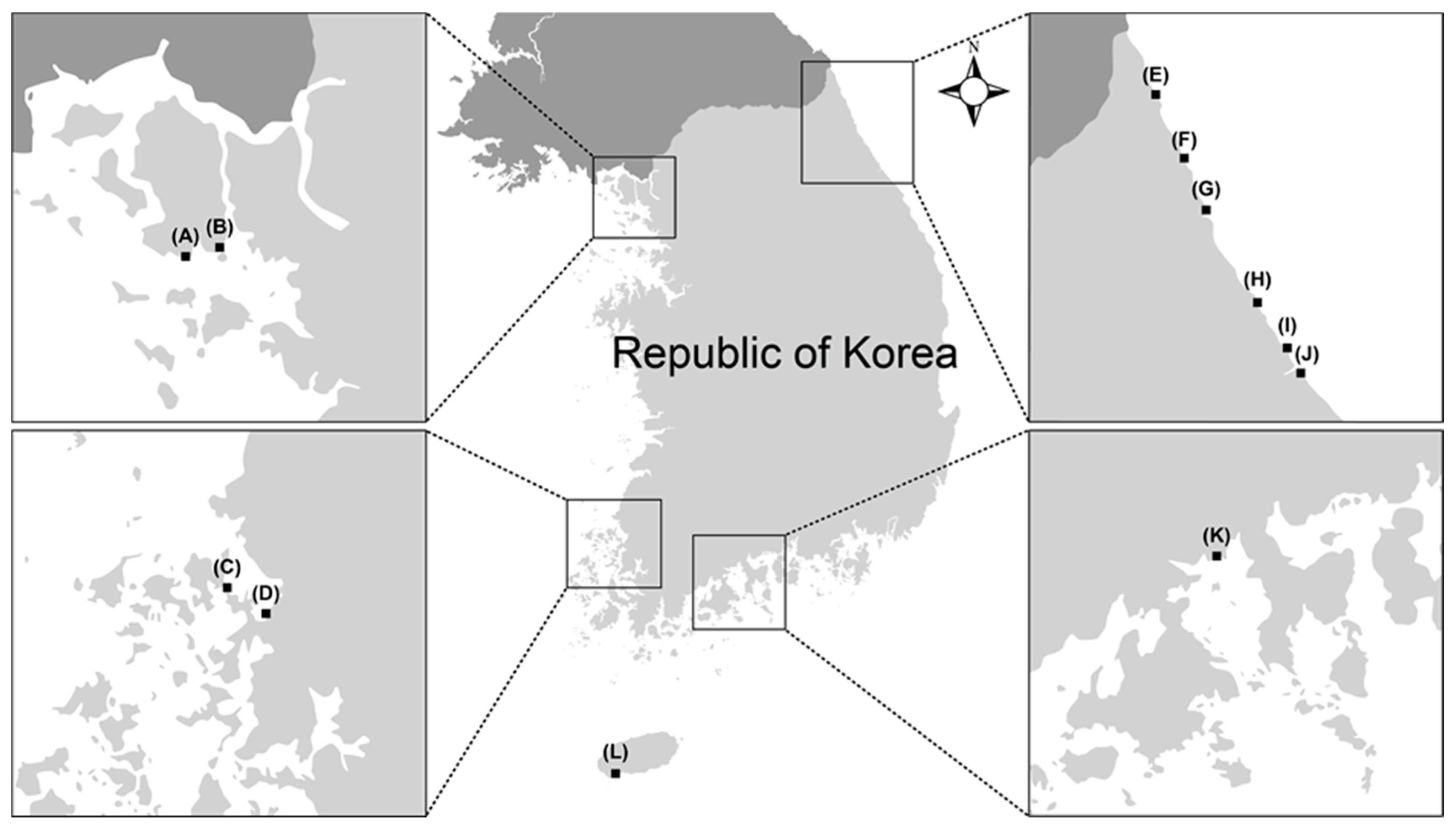
| Identity | ID | Sampling Date | Sampling SITE * | Isolation Source |
|---|---|---|---|---|
| Trichoderma afroharzianum | SFC20160907-M20 | January 2015 | Hyeonnae-myeon, Goseong-gun, Gangwon-do E | Eggs of Arctoscopus japonicus |
| SFC20180619-M25 | January 2017 | Gilsang-myeon, Ganghwa-gun, Incheon B | Mudflat | |
| SFC20190312-M15 | July 2016 | Gilsang-myeon, Ganghwa-gun, Incheon B | Mudflat | |
| T. asperelloides | SFC20180619-M20 | January 2015 | Hyeonnae-myeon, Goseong-gun, Gangwon-do E | Eggs of Arctoscopus japonicus |
| T. asperellum | SFC20160907-M21 | January 2015 | Yeongok-myeon, Gangneung-si, Gangwon-do J | Eggs of Arctoscopus japonicus |
| SFC20180619-M22 | October 2016 | Hyeongyeong-myeon, Muan-gun, Jeollanam-do D | Mudflat | |
| SFC20180619-M24 | October 2016 | Byeollyang-myeon, Suncheon-si, Jeollanam-do K | Sea sand | |
| T. atroviride | SFC20190312-M11 | January 2015 | Hyeonnae-myeon, Goseong-gun, Gangwon-do E | Eggs of Arctoscopus japonicus |
| T. atroviride | SFC20161110-M05 | December 2015 | Jugwang-myeon, Goseong-gun, Gangwon-do F | Agarum clathratum |
| T. bissettii | SFC20170821-M05 | October 2016 | Byeollyang-myeon, Suncheon-si, Jeollanam-do K | Mudflat |
| T. capillare | SFC20180619-M19 | January 2015 | Sokcho-si, Gangwon-do G | Eggs of Arctoscopus japonicus |
| T. citrinoviride | SFC20180510-M16 | July 2016 | Hwado-myeon, Ganghwa-gun, Incheon A | Sea sand |
| T. gamsii | SFC20160907-M22 | January 2015 | Yeongok-myeon, Gangneung-si, Gangwon-do J | Eggs of Arctoscopus japonicus |
| T. guizhouense | SFC20160907-M23 | January 2015 | Yeongok-myeon, Gangneung-si, Gangwon-do J | Eggs of Arctoscopus japonicus |
| SFC20180619-M21 | August 2015 | Jumunjin-eup, Gangneung-si, Gangwon-do I | Agarum clathratum | |
| SFC20180619-M23 | October 2016 | Hyeongyeong-myeon, Muan-gun, Jeollanam-do D | Mudflat | |
| SFC20190312-M12 | January 2017 | Andeok-myeon, Seogwipo-si, Jeju-do L | Sea sand | |
| T. hamatum | SFC20180510-M09 | July 2016 | Gilsang-myeon, Ganghwa-gun, Incheon B | Mudflat |
| T. longibrachiatum | SFC20171019-M03 | July 2016 | Gilsang-myeon, Ganghwa-gun, Incheon B | Mudflat |
| T. orientalis | SFC20170718-M02 | January 2017 | Byeollyang-myeon, Suncheon-si, Jeollanam-do K | Mudflat |
| T. paraviridescens | SFC20160907-M24 | January 2015 | Yeongok-myeon, Gangneung-si, Gangwon-do J | Eggs of Arctoscopus japonicus |
| T. pyramidale | SFC20160907-M25 | January 2015 | Yeongok-myeon, Gangneung-si, Gangwon-do J | Eggs of Arctoscopus japonicus |
| T. songyi | SFC20171120-M04 | October 2016 | Hyeongyeong-myeon, Muan-gun, Jeollanam-do C | Sea sand |
| T. subviride | SFC20170919-M07 | July 2016 | Andeok-myeon, Seogwipo-si, Jeju-do L | Sea sand |
| T. virens | SFC20180817-M24 | October 2016 | Hyeongyeong-myeon, Muan-gun, Jeollanam-do D | Mudflat |
| Trichoderma sp. 1 | SFC20190312-M13 | January 2015 | Yeongok-myeon, Gangneung-si, Gangwon-do J | Eggs of Arctoscopus japonicus |
| SFC20190312-M14 | July 2016 | Gilsang-myeon, Ganghwa-gun, Incheon B | Mudflat | |
| SFC20190312-M17 | November 2015 | Hyeonnam-myeon, Yangyang-gun, Gangwon-do H | Agarum clathratum | |
| Trichoderma sp. 2 | SFC20190312-M16 | October 2015 | Hyeonnam-myeon, Yangyang-gun, Gangwon-do H | Agarum clathratum |
| Trichoderma sp. 3 | SFC20161110-M06 | September 2015 | Jumunjin-eup, Gangneung-si, Gangwon-do I | Agarum clathratum |
| Fungal Name | ID | Radical-Scavenging Activity (%) | Antifungal Activity (MIC 3, μg/mL) | Tyrosinase Inhibition (IC50 4, μg/mL) | ||
|---|---|---|---|---|---|---|
| ABTS 1 | DPPH 2 | L. thalassiae | A. cruciatus | |||
| T. afroharzianum | SFC20160907-M20 | 27.16 | 24.47 | N.D. | 100 | N.D. 5 |
| SFC20190312-M15 | 69.92 | 25.29 | N.D. | >100 | N.D. | |
| SFC20180619-M25 | 66.70 | 33.77 | >100 | >100 | N.D. | |
| T. asperelloides | SFC20180619-M20 | 47.70 | 41.06 | 50 | >100 | N.D. |
| T. asperellum | SFC20160907-M21 | 73.38 | 30.62 | 6.25 | 50 | >417 |
| SFC20180619-M22 | 74.70 | 37.83 | >100 | 25 | >417 | |
| SFC20180619-M24 | 72.63 | 32.34 | 6.25 | 25 | 104.23 | |
| T. atroviride | SFC20161110-M05 | 32.18 | 38.03 | N.D. | N.D. | 15.30 |
| SFC20190312-M11 | 48.48 | 34.80 | N.D. | N.D. | >417 | |
| T. bissettii | SFC20170821-M05 | 57.17 | 79.60 | N.D. | >100 | >417 |
| T. capillare | SFC20180619-M19 | 29.37 | 49.62 | >100 | >100 | N.D. |
| T. citrinoviride | SFC20180510-M16 | 25.52 | 48.27 | >100 | >100 | N.D. |
| T. gamsii | SFC20160907-M22 | 15.82 | 14.69 | N.D. | N.D. | 52.08 |
| T. guizhouense | SFC20160907-M23 | 51.18 | 31.27 | N.D. | N.D. | N.D. |
| SFC20180619-M21 | 14.65 | 18.62 | >100 | N.D. | N.D. | |
| SFC20180619-M23 | 18.17 | 23.41 | N.D. | N.D. | 7.28 | |
| SFC20190312-M12 | 32.73 | 36.76 | N.D. | N.D. | 152.07 | |
| T. hamatum | SFC20180510-M09 | 23.29 | 35.33 | N.D. | N.D. | N.D. |
| T. longibrachiatum | SFC20171019-M03 | 66.36 | 72.72 | N.D. | >100 | 252.26 |
| T. orientalis | SFC20170718-M02 | 28.62 | 32.22 | N.D. | N.D. | N.D. |
| T. paraviridescens | SFC20160907-M24 | 49.59 | 38.32 | N.D. | >100 | 118.61 |
| T. pyramidale | SFC20160907-M25 | 25.02 | 2.36 | N.D. | >100 | N.D. |
| T. songyi | SFC20171120-M04 | 27.14 | 14.60 | N.D. | N.D. | 23.77 |
| T. subviride | SFC20170919-M07 | 61.71 | 53.43 | N.D. | N.D. | N.D. |
| T. virens | SFC20180817-M24 | 42.20 | 45.53 | >100 | 100 | N.D. |
| Trichoderma sp. 1 | SFC20190312-M14 | 42.09 | 44.05 | N.D. | N.D. | N.D. |
| SFC20190312-M13 | 61.57 | 55.03 | >100 | N.D. | 119.74 | |
| SFC20190312-M17 | 40.11 | 55.81 | >100 | N.D. | 148.1 | |
| Trichoderma sp. 2 | SFC20190312-M16 | 48.48 | 34.80 | N.D. | N.D. | N.D. |
| Trichoderma sp. 3 | SFC20161110-M06 | 43.96 | 33.57 | N.D. | N.D. | N.D. |
| Ascorbic acid * | 13.70 | 6.80 | ||||
| Kojic acid * | 49.32 | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.; Heo, Y.M.; Jang, S.; Lee, H.; Kwon, S.-L.; Park, M.S.; Lim, Y.W.; Kim, J.-J. Diversity of Trichoderma spp. in Marine Environments and Their Biological Potential for Sustainable Industrial Applications. Sustainability 2020, 12, 4327. https://0-doi-org.brum.beds.ac.uk/10.3390/su12104327
Kim K, Heo YM, Jang S, Lee H, Kwon S-L, Park MS, Lim YW, Kim J-J. Diversity of Trichoderma spp. in Marine Environments and Their Biological Potential for Sustainable Industrial Applications. Sustainability. 2020; 12(10):4327. https://0-doi-org.brum.beds.ac.uk/10.3390/su12104327
Chicago/Turabian StyleKim, Kyeongwon, Young Mok Heo, Seokyoon Jang, Hanbyul Lee, Sun-Lul Kwon, Myung Soo Park, Young Woon Lim, and Jae-Jin Kim. 2020. "Diversity of Trichoderma spp. in Marine Environments and Their Biological Potential for Sustainable Industrial Applications" Sustainability 12, no. 10: 4327. https://0-doi-org.brum.beds.ac.uk/10.3390/su12104327