Maltose and Totally Impermeable Film Enhanced Suppression of Anaerobic Soil Disinfestation on Soilborne Pathogens and Increased Strawberry Yield

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design in Laboratory
2.1.1. Soil Sample Collection
2.1.2. Orthogonal Experimental Design
2.1.3. Experimental for Cover Tarps
2.2. Field Trials
2.2.1. Field Trials Design
2.2.2. Soil Sampling
2.2.3. Soil Physical and Chemical Properties
2.2.4. Strawberry Plant Growth, Mortality and Crop Yield
2.2.5. DNA Extraction
2.3. Statistical Analyses
2.3.1. Laboratory Studies
2.3.2. Field Trials
2.3.3. High Throughput Sequencing Data Analysis
3. Results
3.1. Laboratory Study
Orthogonal Experiments
3.2. Field Trials
3.2.1. Soil Temperature and Moisture
3.2.2. Soilborne Fungal Pathogens
3.2.3. Soil Physicochemical Parameters
3.2.4. Strawberry Growth, Mortality and Yield
3.2.5. Microbiota Analysis by High Throughput Sequencing (HTS)
Alpha Diversity Analysis
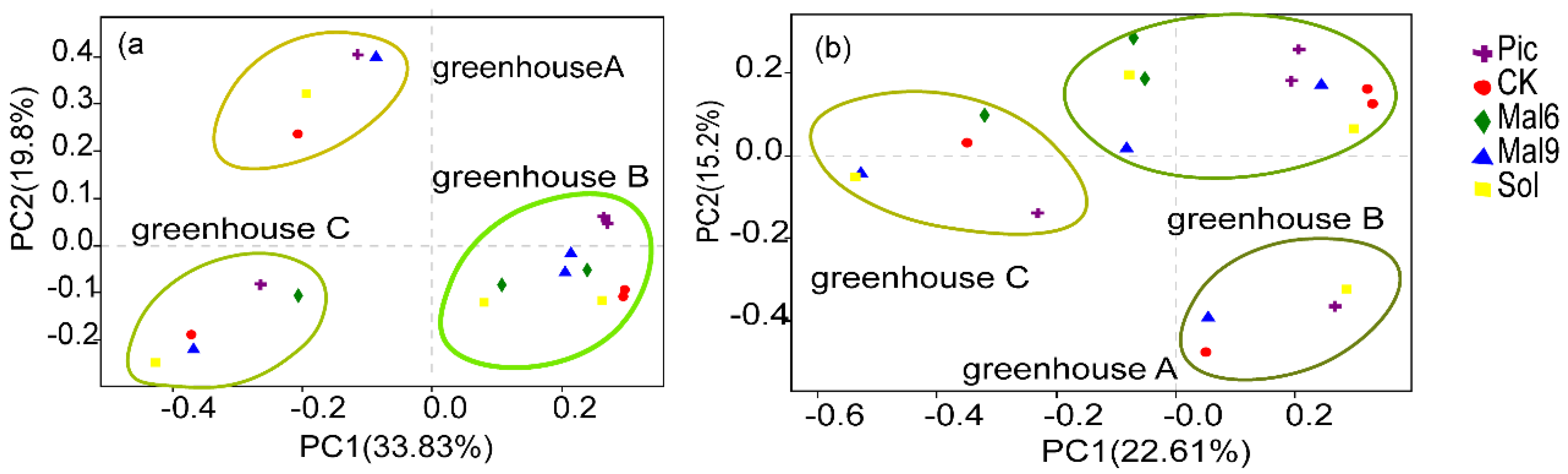
Principal Coordinate Analysis
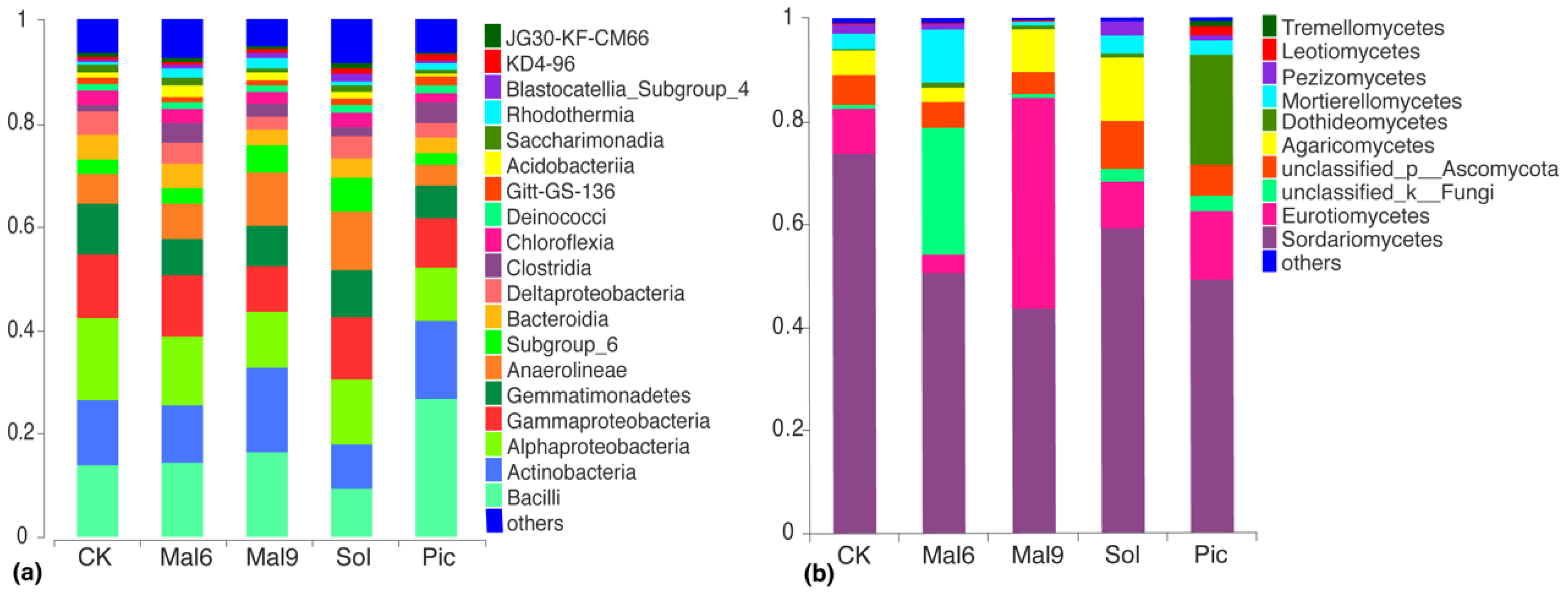
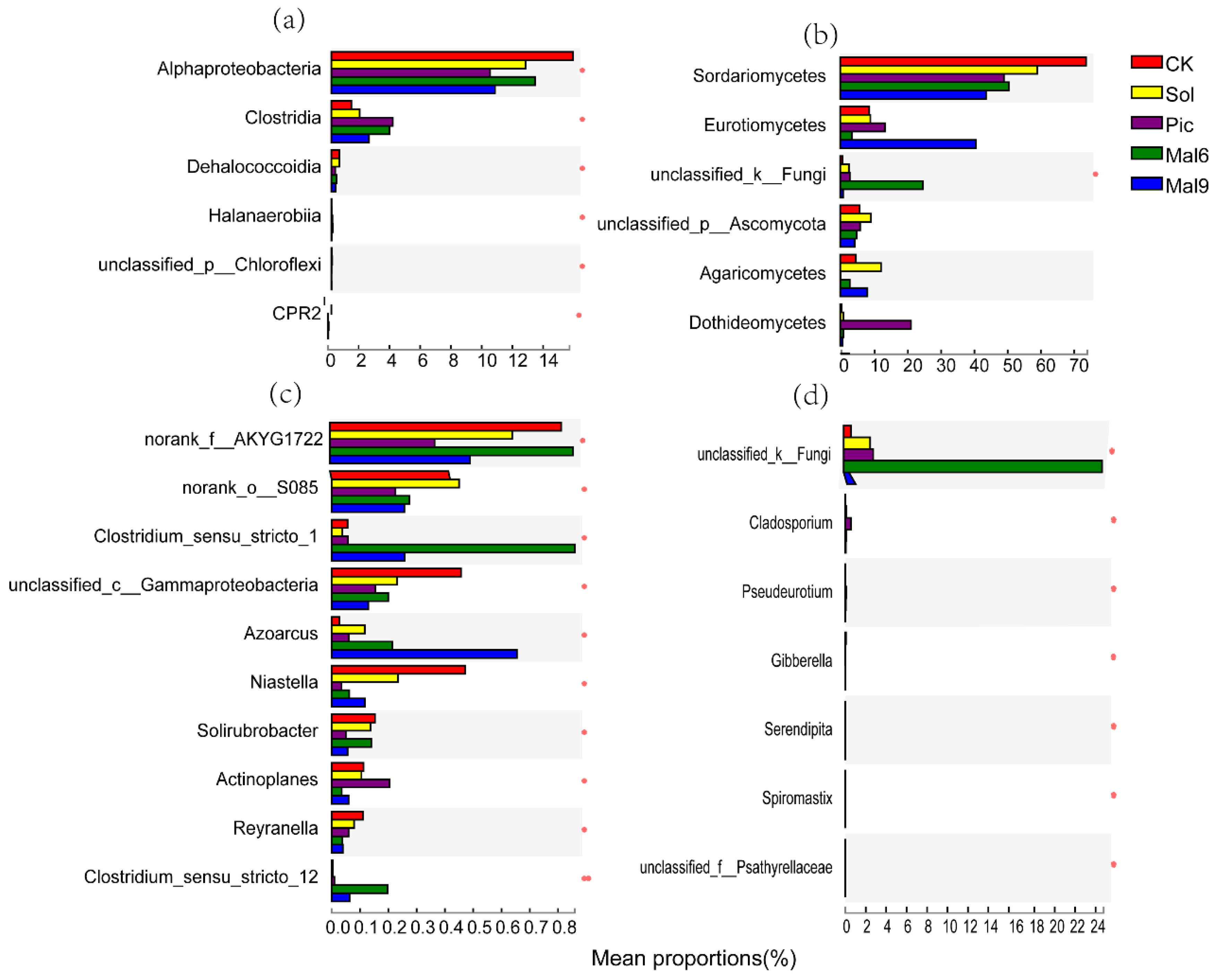
Microbiota Composition
4. Discussion
4.1. Effect of ASD on Soil Fungal Pathogenic Microorganisms
4.2. Effect of ASD on the Soil’s Physicochemical Properties
4.3. Effect of ASD on Strawberry Fruit Yield
4.4. Effect of ASD on Soil Bacterial and Fungal Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. FAOSTAT; FAO: Rome, Italy, 2017. [Google Scholar]
- Shennan, C.; Muramoto, J.; Koike, S.; Baird, G.; Fennimore, S.; Samtani, J.; Bolda, M.; Dara, S.; Daugovish, O.; Lazarovits, G.; et al. Anaerobic soil disinfestation is an alternative to soil fumigation for control of some soilborne pathogens in strawberry production. Plant Pathol. 2017, 67, 51–66. [Google Scholar] [CrossRef]
- Huang, X.; Wen, T.; Zhang, J.B.; Meng, L.; Bin Zhu, T.; Liu, L.L.; Cai, Z. Control of soil-borne pathogen Fusarium oxysporum by biological soil disinfestation with incorporation of various organic matters. Eur. J. Plant Pathol. 2015, 143, 223–235. [Google Scholar] [CrossRef]
- Koike, S.T.; Gordon, T.R. Management of Fusarium wilt of strawberry. Crop. Prot. 2015, 73, 67–72. [Google Scholar] [CrossRef]
- Zhang, Y. Identification and characterization of a Bacillus subtilis strain TS06 as bio-control agent of strawberry replant disease (Fusarium and Verticilium wilts). Afr. J. Biotechnol. 2012, 11, 570–580. [Google Scholar] [CrossRef]
- Han, D.; Yan, D.; Wang, Q.; Fang, W.; Wang, X.; Li, J.; Wang, D.; Li, Y.; Ouyang, C.; Cao, A. Effects of soil type, temperature, moisture, application dose, fertilizer, and organic amendments on chemical properties and biodegradation of dimethyl disulfide in soil. Land Degrad. Dev. 2018, 29, 4282–4290. [Google Scholar] [CrossRef]
- Yan, D.; Wang, Q.; Mao, L.; Xie, H.; Guo, M.; Cao, A. Evaluation of chloropicrin gelatin capsule formulation as a soil fumigant for greenhouse strawberry in China. J. Agric. Food Chem. 2012, 60, 5023–5027. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yan, D.; Fang, W.; Huang, B.; Wang, X.; Wang, X.; Zhu, J.; Liu, J.; Ouyang, C.; Li, Y.; et al. Chloropicrin alternated with biofumigation increases crop yield and modifies soil bacterial and fungal communities in strawberry production. Sci. Total Environ. 2019, 675, 615–622. [Google Scholar] [CrossRef]
- Donley, N. The USA lags behind other agricultural nations in banning harmful pesticides. Environ. Health 2019, 18, 44. [Google Scholar] [CrossRef] [Green Version]
- Fennimore, S.A.; Serohijos, R.; Samtani, J.B.; Ajwa, H.A.; Subbarao, K.V.; Martin, F.N.; Daugovish, O.; Legard, D.; Browne, G.T.; Muramoto, J.; et al. TIF film, substrates and nonfumigant soil disinfestation maintain fruit yields. Calif. Agric. 2013, 67, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.M.; Pruden, G.; Jenkinson, D.S. Mineralization and immobilization of nitrogen in fumigated soil and the measurement of microbial biomass nitrogen. Soil Biol. Biochem. 1984, 16, 437–444. [Google Scholar] [CrossRef]
- Yan, D.; Wang, Q.; Li, Y.; Ouyang, C.; Guo, M.; Cao, A. Analysis of the inhibitory effects of chloropicrin fumigation on nitrification in various soil types. Chemosphere 2017, 175, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Wang, Q.; Mao, L.; Li, W.; Xie, H.; Guo, M.; Cao, A. Quantification of the effects of various soil fumigation treatments on nitrogen mineralization and nitrification in laboratory incubation and field studies. Chemosphere 2013, 90, 1210–1215. [Google Scholar] [CrossRef]
- Shrestha, U.; Augé, R.M.; Butler, D.M. A meta-analysis of the impact of anaerobic soil disinfestation on pest suppression and yield of horticultural crops. Front. Plant Sci. 2016, 7, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Molendijk, L.P.G.; Bleeker, P.O.; Lamers, J.G.; Runia, W.T. Perspectives of anaerobic soil disinfestation. In Proceedings of the VII International Symposium on Chemical and Non-Chemical Soil and Substrate Disinfestation 883, Leuven, Belgium, 13–18 September 2009; pp. 277–283. [Google Scholar] [CrossRef] [Green Version]
- Momma, N.; Kobara, Y.; Uematsu, S.; Kita, N.; Shinmura, A. Development of biological soil disinfestations in Japan. Appl. Microbiol. Biotechnol. 2013, 97, 3801–3809. [Google Scholar] [CrossRef] [PubMed]
- Shennan, C.; Muramoto, J.; Lamers, J.; Mazzola, M.; Rosskopf, E.; Kokalis-Burelle, N.; Momma, N.; Butler, D.; Kobara, Y. Anaerobic soil disinfestation for soil borne disease control in strawberry and vegetable systems: Current knowledge and future directions. Acta Hortic. 2014, 1044, 165–175. [Google Scholar] [CrossRef]
- Strauss, S.L.; Kluepfel, D.A. Anaerobic soil disinfestation: A chemical-independent approach to pre-plant control of plant pathogens. J. Integr. Agric. 2015, 14, 2309–2318. [Google Scholar] [CrossRef]
- Strauss, S.L.; Greenhut, R.F.; McClean, A.E.; Kluepfel, D.A. Effect of anaerobic soil disinfestation on the bacterial community and key soilborne phytopathogenic agents under walnut tree-crop nursery conditions. Plant Soil 2017, 415, 493–506. [Google Scholar] [CrossRef]
- Butler, D.; Kokalis-Burelle, N.; Albano, J.P.; Mccollum, T.G.; Muramoto, J.; Shennan, C.; Rosskopf, E.N. Anaerobic Soil Disinfestation (ASD) Combined with Soil Solarization as a Methyl Bromide Alternative: Vegetable Crop Performance and Soil Nutrient Dynamics. Plant Soil 2014, 378, 365–381. [Google Scholar] [CrossRef]
- Wang, X.; Fang, W.; Yan, D.; Han, D.; Liu, J.; Ren, Z.; Ouyang, C.; Li, Y.; Wang, Q.; Cao, A. Evaluation of the influence of temperature and relative humidity on the permeability of four films to the fumigant dimethyl disulfide. J. Environ. Manag. 2019, 236, 687–694. [Google Scholar] [CrossRef]
- Browne, G.T.; Ott, N.; Poret-Peterson, A.; Gouran, H.; Lampinen, B. Efficacy of anaerobic soil disinfestation for control of prunus replant disease. Plant Dis. 2018, 102, 209–219. [Google Scholar] [CrossRef]
- Gao, S.; Sosnoskie, L.M.; Cabrera, J.A.; Qin, R.; Hanson, B.D.; Gerik, J.S.; Wang, N.; Browne, G.T.; Thomas, J.E. Fumigation efficacy and emission reduction using low-permeability film in orchard soil fumigation. Pest Manag. Sci. 2016, 72, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.C.; Freeman, J.H.; Dittmar, P.J. Impact of Totally Impermeable Film on the Efficacy of 1,3-Dichloropropene and Chloropicrin Mixtures for the Control of Nutsedge. Weed Technol. 2016, 30, 910–918. [Google Scholar] [CrossRef]
- Browning, M.; Dawson, C.; Alm, S.R.; Gorrës, J.H.; Amador, J.A. Differential effects of butyric acid on nematodes from four trophic groups. Appl. Soil Ecol. 2004, 27, 47–54. [Google Scholar] [CrossRef]
- Qin, R.; Gao, S.; Ajwa, H.; Sullivan, D.; Wang, D.; Hanson, B.D. Field Evaluation of a New Plastic Film (Vapor Safe) to Reduce Fumigant Emissions and Improve Distribution in Soil. J. Environ. Qual. 2011, 40, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Poret-Peterson, A.T.; Albu, S.; McClean, A.E.; Kluepfel, D.A. Shifts in soil bacterial communities as a function of carbon source used during anaerobic soil disinfestation. Front. Environ. Sci. 2019, 6, 1–15. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Doornbos, R.F.; Van Loon, L.C.; Bakker, P.A.H.M. Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere. A review. Agron. Sustain. Dev. 2012, 32, 227–243. [Google Scholar] [CrossRef]
- Li, X.; Ding, C.; Zhang, T.; Wang, X. Fungal pathogen accumulation at the expense of plant-beneficial fungi as a consequence of consecutive peanut monoculturing. Soil Biol. Biochem. 2014, 72, 11–18. [Google Scholar] [CrossRef]
- Komada, H. Development of a selective medium for quantitative isolation of Fusarium oxysporum from natural soil. Rev. Plant Prot. Res. 1975, 8, 114–124. [Google Scholar]
- Yoshikawa, M.; Masago, H.; Keen, N.T. Activated synthesis of poly (A)-containing messenger RNA in soybean hypocotyls inoculated with Phytophthora megasperma var. sojae. Physiol. Plant Pathol. 1977, 10, 125–138. [Google Scholar] [CrossRef]
- Schumacher, B.A. Methods for the Determination of Total Organic Carbon (TOC) in Soils and Sediments; United States Environmental, Protection Agency Environmental Sciences Division Nationalxposure Research Laboratory: Las Vefas, NV, USA, 2002; pp. 103–119. [Google Scholar]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; United States Department of Agriculture: Washington, DC, USA, 1954.
- Li, J.; Huang, B.; Wang, Q.; Li, Y.; Fang, W.; Yan, D.; Guo, M.; Cao, A. Effect of fumigation with chloropicrin on soil bacterial communities and genes encoding key enzymes involved in nitrogen cycling. Environ. Pollut. 2017, 227, 534–542. [Google Scholar] [CrossRef]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Massart, S.; Olmos, A.; Jijakli, H.; Candresse, T. Current impact and future directions of high throughput sequencing in plant virus diagnostics. Virus Res. 2014, 188, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Blok, W.J.; Lamers, J.G.; Termorshuizen, A.J.; Bollen, G.J. Control of soilborne plant pathogens by incorporating fresh organic amendments followed by tarping. Phytopathology 2000, 90, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, D.M.; Kokalis-Burelle, N.; Muramoto, J.; Shennan, C.; McCollum, T.G.; Rosskopf, E.N. Impact of anaerobic soil disinfestation combined with soil solarization on plant-parasitic nematodes and introduced inoculum of soilborne plant pathogens in raised-bed vegetable production. Crop. Prot. 2012, 39, 33–40. [Google Scholar] [CrossRef]
- Khadka, R.B.; Marasini, M.; Rawal, R.; Testen, A.L.; Miller, S.A. Effects of anaerobic soil disinfestation carbon sources on soilborne diseases and weeds of okra and eggplant in Nepal. Crop. Prot. 2020, 104846. [Google Scholar] [CrossRef]
- Liu, L.; Kong, J.; Cui, H.; Zhang, J.; Wang, F.; Cai, Z.; Huang, X. Relationships of decomposability and C/N ratio in different types of organic matter with suppression of Fusarium oxysporum and microbial communities during reductive soil disinfestation. Biol. Control. 2016, 101, 103–113. [Google Scholar] [CrossRef]
- Paudel, B.R.; Di Gioia, F.; Zhao, X.; Ozores-Hampton, M.; Hong, J.; Kokalis-Burelle, N.; Pisani, C.; Rosskopf, E.N. Evaluating anaerobic soil disinfestation and other biological soil management strategies for open-field tomato production in Florida. Renew. Agric. Food Syst. 2018. [Google Scholar] [CrossRef]
- Liu, L.; Chen, S.; Zhao, J.; Zhou, X.; Wang, B.; Li, Y.; Zheng, G.; Zhang, J.; Cai, Z.; Huang, X. Watermelon planting is capable to restructure the soil microbiome that regulated by reductive soil disinfestation. Appl. Soil Ecol. 2018, 129, 52–60. [Google Scholar] [CrossRef]
- Momma, N. Biological soil disinfestation (BSD) of soilborne pathogens and its possible mechanisms. Jpn. Agric. Res. Q. 2008, 42, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Ueki, A.; Takehara, T.; Ishioka, G.; Kaku, N.; Ueki, K. Degradation of the fungal cell wall by clostridial strains isolated from soil subjected to biological soil disinfestation and biocontrol of Fusarium wilt disease of spinach. Appl. Microbiol. Biotechnol. 2017, 101, 8267–8277. [Google Scholar] [CrossRef] [PubMed]
- Muck, R.E. Silage microbiology and its control through additives. Rev. Bras. Zootec. 2010, 39, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Bartkiene, E.; Lele, V.; Ruzauskas, M.; Domig, K.; Starkute, V.; Zavistanaviciute, P.; Bartkevics, V.; Pugajeva, I.; Klupsaite, D.; Juodeikiene, G.; et al. Lactic acid bacteria isolation from spontaneous sourdough and their characterization including antimicrobial and antifungal properties evaluation. Microorganisms 2020, 8. [Google Scholar] [CrossRef] [Green Version]
- Ebihara, Y.; Uematsu, S. Survival of strawberry-pathogenic fungi Fusarium oxysporum f. sp. fragariae, Phytophthora cactorum and Verticillium dahliae under anaerobic conditions. J. Gen. Plant Pathol. 2014, 80, 50–58. [Google Scholar] [CrossRef]
- Shrestha, U.; Dee, M.E.; Ownley, B.H.; Butler, D.M. Anaerobic soil disinfestation reduces germination and affects colonization of sclerotium rolfsii sclerotia. Phytopathology 2018, 108, 342–351. [Google Scholar] [CrossRef] [Green Version]
- Hewavitharana, S.S.; Mazzola, M. Carbon source-dependent effects of anaerobic soil disinfestation on soil microbiome and suppression of Rhizoctonia solani AG-5 and Pratylenchus penetrans. Phytopathology 2016, 106, 1015–1028. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.; Yan, D.; Huang, B.; Ren, Z.; Wang, X.; Liu, X.; Li, Y.; Ouyang, C.; Migheli, Q.; Cao, A.; et al. Biochemical pathways used by microorganisms to produce nitrous oxide emissions from soils fumigated with dimethyl disulfide or allyl isothiocyanate. Soil Biol. Biochem. 2019, 132, 1–13. [Google Scholar] [CrossRef]
- Muramoto, J.; Shennan, C.; Zavatta, M.; Baird, G.; Toyama, L.; Mazzola, M. Effect of Anaerobic Soil Disinfestation and Mustard Seed Meal for Control of Charcoal Rot in California Strawberries. Int. J. Fruit Sci. 2016, 16, 59–70. [Google Scholar] [CrossRef]
- Testen, A.L.; Miller, S.A. Carbon source and soil origin shape soil microbiomes and tomato soilborne pathogen populations during anaerobic soil disinfestation. Phytobiomes J. 2018, 2, 138–150. [Google Scholar] [CrossRef]
- Penton, C.R.; Gupta, V.V.S.R.; Yu, J.; Tiedje, J.M. Size matters: Assessing optimum soil sample size for fungal and bacterial community structure analyses using high throughput sequencing of rRNA gene amplicons. Front. Microbiol. 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Goldfarb, K.C.; Karaoz, U.; Hanson, C.A.; Santee, C.A.; Bradford, M.A.; Treseder, K.K.; Wallenstein, M.D.; Brodie, E.L. Differential growth responses of soil bacterial taxa to carbon substrates of varying chemical recalcitrance. Front. Microbiol. 2011, 2, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueki, A.; Takehara, T.; Ishioka, G.; Kaku, N.; Ueki, K. Production of β-1,3-glucanase and chitosanase from clostridial strains isolated from the soil subjected to biological disinfestation. AMB Express 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; He, Y.L.; Zhang, X.F. Nitrogen removal capability through simultaneous heterotrophic nitrification and aerobic denitrification by Bacillus sp. LY. Environ. Technol. 2010, 31, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Beust, A.; Sappa, P.K.; Völker, U.; Dinse, T.; Herglotz, J.; Reinhold-Hurek, B. Two functionally deviating type 6 secretion systems occur in the nitrogen-fixing endophyte azoarcus olearius BH72. Front. Microbiol. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Klaubauf, S.; Inselsbacher, E.; Zechmeister-Boltenstern, S.; Wanek, W.; Gottsberger, R.; Strauss, J.; Gorfer, M. Molecular diversity of fungal communities in agricultural soils from Lower Austria. Fungal Divers. 2010, 44, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Jiang, X.; Zhou, B.; Zhao, B.; Ma, M.; Guan, D.; Li, J.; Chen, S.; Cao, F.; Shen, D.; et al. Thirty four years of nitrogen fertilization decreases fungal diversity and alters fungal community composition in black soil in northeast China. Soil Biol. Biochem. 2016, 95, 135–143. [Google Scholar] [CrossRef]
- Liu, L.; Huang, X.; Zhao, J.; Zhang, J.; Caia, Z. Characterizing the key agents in a disease-suppressed soil managed by reductive soil disinfestation. Appl. Environ. Microbiol. 2019, 85, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yun, S.H.; Arie, T.; Kaneko, I.; Yoder, O.C.; Turgeon, B.G. Molecular organization of mating type loci in heterothallic, homothallic, and asexual gibberella/fusarium species. Fungal Genet. Biol. 2000, 31, 7–20. [Google Scholar] [CrossRef]
- Abdel-Baky, N.F.; Abdel-Salam, A.H. Natural incidence of Cladosporium spp. as a bio-control agent against whiteflies and aphids in Egypt. J. Appl. Entomol. 2003, 127, 228–235. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatments * | Organic Amendment | Rate (%, w/w) | Film Type | Formulated Product Manufacturer |
|---|---|---|---|---|
| CK | – | – | – | |
| VIFCK | – | – | 0.03 mm VIF | VIF: Guangdong Jinming Machinery Co., Ltd. China |
| VIFMal | maltose | 5 | 0.03 mm VIF | |
| PECK | – | – | 0.05 mm PE | PE: Shandong Longxin Plastic Industry Co., Ltd. China |
| PEMal | maltose | 5 | 0.05 mm PE | |
| TIFCK | – | – | 0.05 mm TIF | TIF: Shandong Longxin Plastic Industry Co., Ltd. China |
| TIFMal | maltose | 5 | 0.05 mm TIF | |
| WaterCK | – | – | 150 mL deionized water | |
| WaterMal | maltose | 5 | 150 mL deionized water |
| Site | pH (1:2.5) | Organic Matter Content (g kg−1) | NH4+-N (mg kg−1) | NO3−-N (mg kg−1) | Available Phosphorus (mg kg−1) | Available Potassium (mg kg−1) | Oxidation-Reduction (mv) | Electrical Conductivity (μs cm−1) |
|---|---|---|---|---|---|---|---|---|
| Trial I | 7.83 | 13.00 | 6.61 | 144.69 | 701.67 | 452.20 | 209.50 | 1222.33 |
| Trial II | 7.98 | 23.63 | 8.09 | 152.40 | 1259.00 | 912.90 | 208.50 | 949.67 |
| Site | Strawberry Variety | Plot Area (m2) | Disinfestation Date | Film Removal | Strawberry Transplant | End of the Trial |
|---|---|---|---|---|---|---|
| Trial I | Red face | 2.8 × 9.5 = 26 | 2017/7/20 | 2017/8/12 | 2017/9/30 | 2018/6/4 |
| Trial II | Red face | 3.2 × 7.5 = 22 | 2017/7/27 | 2017/8/23 | 2017/8/28 | 2018/5/15 |
| Site | Treatments | Dose (t/ha) | Application Method | Water Consumption (t/ha) | Film Type (0.05 mm) | Cover Time (weeks) | Replicates |
|---|---|---|---|---|---|---|---|
| Trial I | Mal 6 | 6 | Drip irrigation | 50 | TIF | 3 | 4 |
| Mal 9 | 9 | Drip irrigation | 50 | TIF | 3 | 4 | |
| Pic | 0.375 | Machine Injection | – | PE | 2 | 4 | |
| Sol | – | Drip irrigation | 50 | TIF | 3 | 4 | |
| CK | – | – | – | – | – | 4 | |
| Trial II | Mal 6 | 6 | Drip irrigation | 50 | TIF | 3 | 4 |
| Mal 9 | 9 | Drip irrigation | 50 | TIF | 3 | 4 | |
| Pic | 0.375 | Machine Injection | – | PE | 2 | 4 | |
| Sol c | – | Drip irrigation | 50 | TIF | 3 | 4 | |
| CK | – | – | – | – | – | 4 |
| Fungi | Source a | Sum of Squares | Degree of Freedom | Mean Square | F | Significant |
|---|---|---|---|---|---|---|
| FUS | A | 621 | 2 | 310 | 2.43 | |
| B | 1994 | 2 | 997 | 7.81 | * | |
| C | 3063 | 2 | 1531 | 12.00 | * | |
| A × B | 4176 | 4 | 1044 | 8.18 | * | |
| A × C | 864 | 4 | 216 | 1.69 | ||
| B × C | 677 | 4 | 169 | 1.33 | ||
| Error | 1021 | 8 | 128 | |||
| PFY | A | 156 | 2 | 78 | 0.27 | |
| B | 7339 | 2 | 3670 | 12.74 | * | |
| C | 3135 | 2 | 1568 | 5.44 | * | |
| A × B | 310 | 4 | 77 | 0.27 | ||
| A × C | 3910 | 4 | 978 | 3.39 | ||
| B × C | 2853 | 4 | 713 | 2.48 | ||
| Error | 2305 | 8 | 288 |
| Treatments | Rate (%, w/w) | Tarp Kind | FUS | PHY | ||
|---|---|---|---|---|---|---|
| CFUg−1 * | % Reduction | CFU g−1 | % Reduction | |||
| CK | – | – | 11,960 ± 339 b | – | 14,234 ± 320 a | – |
| VIFCK | – | 0.05 mm VIF | 14,400 ± 200 ab | – | 16,767 ± 306 a | – |
| VIFMal | 5 | 0.05 mm VIF | 442 ± 31 d | 96.3 | 675 ± 146 d | 91.0 |
| PECK | – | 0.05 mm PE | 18,700 ± 700 ab | – | 21,600 ± 541 a | – |
| PEMal | 5 | 0.05 mm PE | 1912 ± 20 c | 84.0 | 1925 ± 357 c | 86.5 |
| TIFCK | – | 0.03 mm TIF | 17,967 ± 404 ab | – | 17,867 ± 729 a | – |
| TIFMal | 5 | 0.03 mm TIF | 58 ± 8 d | 99.5 | 100 ± 26 d | 99.3 |
| WaterCK | – | 150 mL deionized water | 15,155 ± 751 ab | – | 14,810 ± 834 a | – |
| WaterMal | 5 | 150 mL deionized water | 34,696 ± 1175 a | – | 6970 ± 485 b | 51.0 |
| Item | Soil Temperature at Different Depths (°C) | Soil Moisture (%) | ||||
|---|---|---|---|---|---|---|
| −20 cm | −15 cm | −10 cm | −5 cm | 0 cm | ||
| Trial I | ||||||
| Accumulated temperature * | 1016 | 1022 | 1031 | 1048 | 987 | – |
| Average value | 36.3 | 36.5 | 36.8 | 37.4 | 35.2 | 36.2 |
| Highest value | 42.9 | 45.0 | 46.6 | 52.0 | 78.0 | 41.9 |
| Lowest value | 27.9 | 25.8 | 24.3 | 24.1 | 23.4 | 28.6 |
| Trial II | ||||||
| Accumulated temperature * | 1007 | 1019 | 1039 | 1060 | 1011 | – |
| Average value | 37.1 | 37.6 | 38.3 | 39.1 | 37.3 | 36.8 |
| Highest value | 44.5 | 48.8 | 49.5 | 54.3 | 61.2 | 43.4 |
| Lowest value | 32.4 | 31.5 | 30.0 | 29.4 | 24.2 | 30.5 |
| Site | Treatments | FUS (30 d) | PHY (30 d) | FUS (120 d) | PHY (120 d) | ||||
|---|---|---|---|---|---|---|---|---|---|
| CFU·g−1 | % Reduction | CFU·g−1 | % Reduction | CFU·g−1 | % Reduction | CFU·g−1 | % Reduction | ||
| Trial I 2017–2018 | Mal 6 | 830 ± 154 c | 88.9 | 1040 ± 98 c | 84.6 | 2095 ± 128 a | 3288 ± 240 a | ||
| Mal 9 | 681± 95 c | 89.3 | 532 ± 102 d | 92.3 | 2254 ± 340 a | 3173 ± 386 a | |||
| Pic | 33 ± 8 d | 99.4 | 126 ± 30 e | 98.2 | 1630 ± 230 a | 3007 ± 392 a | |||
| Sol | 226 ± 477 b | 77.7 | 3045 ± 128 b | 66.6 | 1541 ± 285 a | 2319 ± 284 a | |||
| CK | 10,255 ± 743 a | 8803 ± 468 a | 899 ± 64 a | 2029 ± 312 a | |||||
| Trial II 2017–2018 | Mal 6 | 422 ± 84 bc | 75.2 | 458 ± 87 bc | 69.7 | 1486 ± 108 a | 2196 ± 114 a | ||
| Mal 9 | 275 ± 46 c | 81.7 | 255 ± 42 c | 75.4 | 1448 ± 56 a | 1668 ± 86 a | 11.3 | ||
| Pic | 14 ± 4 d | 99.1 | 8 ± 3 d | 99.1 | 641 ± 74 a | 24.9 | 1353 ± 164 a | 28.0 | |
| Sol | 629 ± 112 b | 69.7 | 677 ± 88 b | 57.8 | 942 ± 48 a | 1549 ± 35 a | 17.6 | ||
| CK | 2375 ± 340 a | 2743 ± 324 a | 853 ±102 a | 1880 ± 58 a | |||||
| Treatments | NH4+-N (mg·kg−1) | NO3−-N (mg·kg−1) | Available P (mg·kg−-1) | Available K (mg·kg−1) | Organic Matter (mg·kg−1) | Electrical Conductivity (μs·cm−1) | pH (1:2.5) |
|---|---|---|---|---|---|---|---|
| Mal 6 | 28.56 ± 1.5 a | 64.85 ± 1.2 e | 889 ± 36.4 a | 813 ± 70 b | 19.03 ± 1.6 a | 948 ± 49 b | 7.58 ± 0.2 ab |
| Mal 9 | 15.12 ± 1.2 cd | 84.05 ± 8.8 d | 693 ± 7.7 b | 1150 ± 41 a | 19.67 ± 1.0 a | 1171 ± 47 a | 7.54 ± 0.2 ab |
| Pic | 18.76 ± 2.3 bc | 123.95 ± 4.1 b | 800 ± 78.2 ab | 409 ± 37 c | 13.42 ± 1.4 b | 1115 ± 103 ab | 7.41 ± 0.2 b |
| Sol | 23.64 ± 3.1 ab | 103.81 ± 4.0 c | 906 ± 57.2 a | 725 ± 90 b | 16.18 ± 3.0 ab | 1144 ± 47 ab | 7.58 ± 0.1 ab |
| CK | 12.30 ± 1.0 d | 182.91 ± 4.1 a | 931 ± 29.0 a | 800 ± 71 b | 16.43 ± 0.2 ab | 561 ± 52 c | 8.06 ± 0.2 a |
| Treatments | Plant Height (cm) | Stem Diameter (mm) | Plant Height (cm) | Stem Diameter (mm) | Mortality (%) | Yield (t/ha) |
|---|---|---|---|---|---|---|
| Date | 28 December 17 | 28 December 17 | 4 May 18 | 4 May 18 | 5 June 18 | February 2018 |
| Mal 6 | 18.91 ± 0.4 bc | 21.37 ± 1.0 b | 37.20 ± 0.9 a | 36.48 ± 0.6 a | 6.00 ± 0.4 ab | 5.10 ± 0.3 a |
| Mal 9 | 21.99 ± 0.6 a | 27.61 ± 1.8 a | 35.78 ± 0.5 a | 35.76 ± 1.9 a | 4.50 ± 0.5 b | 5.66 ± 0.2 a |
| Pic | 21.36 ± 1.3 ab | 24.11 ± 2.1 ab | 36.15 ± 0.8 a | 34.87 ± 0.9 a | 5.00 ± 0.7 b | 5.49 ± 0.1 a |
| Sol | 19.59 ± 0.9 b | 21.92 ± 0.9 b | 29.35 ± 1.7 b | 30.86 ± 1.7 b | 6.75 ± 0.5 ab | 3.48 ± 0.1 b |
| CK | 18.16 ± 0.7 c | 21.26 ± 0.5 b | 28.53 ± 1.9 b | 28.72 ± 0.9 b | 11.25 ± 0.8 a | 2.35 ± 0.1 c |
| Treatments | Chao | Shannon | Coverage | Shannon Evenness |
|---|---|---|---|---|
| Bacteria | ||||
| Mal 6 | 4419 ± 148 a | 6.42 ± 0.1 a | 0.980 ± 0.001 a | 0.79 ± 0.01 a |
| Mal 9 | 3860 ± 273 a | 5.99 ± 0.2 a | 0.981 ± 0.001 a | 0.76 ± 0.01 a |
| Pic | 4304 ± 471 a | 6.04 ± 0.4 a | 0.980 ± 0.002 a | 0.75 ± 0.03 a |
| Sol | 4603 ± 446 a | 6.40 ± 0.2 a | 0.980 ± 0.002 a | 0.79 ± 0.01 a |
| CK | 4149 ± 171 a | 6.31 ± 0.1 a | 0.980 ± 0.002 a | 0.79 ± 0.01 a |
| Fungi | ||||
| Mal6 | 433 ± 32.8 a | 2.78 ± 0.39 a | 0.998 ± 0.0001 a | 0.47 ± 0.06 a |
| Mal9 | 298 ± 53.5 a | 1.94 ± 0.36 a | 0.999 ± 0.0003 a | 0.36 ± 0.06 a |
| Pic | 342 ± 31.9 a | 2.55 ± 0.35 a | 0.999 ± 0.0002 a | 0.45 ± 0.06 a |
| Sol | 363 ± 56.6 a | 2.82 ± 0.37 a | 0.999 ± 0.0004 a | 0.49 ± 0.06 a |
| CK | 378 ± 63.9 a | 2.46 ± 0.29 a | 0.998 ± 0.0003 a | 0.44 ± 0.04 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Z.; Yan, D.; Fang, W.; Huang, B.; Wang, X.; Zhang, D.; Zhu, J.; Liu, J.; Ouyang, C.; Li, Y.; et al. Maltose and Totally Impermeable Film Enhanced Suppression of Anaerobic Soil Disinfestation on Soilborne Pathogens and Increased Strawberry Yield. Sustainability 2020, 12, 5456. https://0-doi-org.brum.beds.ac.uk/10.3390/su12135456
Song Z, Yan D, Fang W, Huang B, Wang X, Zhang D, Zhu J, Liu J, Ouyang C, Li Y, et al. Maltose and Totally Impermeable Film Enhanced Suppression of Anaerobic Soil Disinfestation on Soilborne Pathogens and Increased Strawberry Yield. Sustainability. 2020; 12(13):5456. https://0-doi-org.brum.beds.ac.uk/10.3390/su12135456
Chicago/Turabian StyleSong, Zhaoxin, Dongdong Yan, Wensheng Fang, Bin Huang, Xianli Wang, Daqi Zhang, Jiahong Zhu, Jie Liu, Canbin Ouyang, Yuan Li, and et al. 2020. "Maltose and Totally Impermeable Film Enhanced Suppression of Anaerobic Soil Disinfestation on Soilborne Pathogens and Increased Strawberry Yield" Sustainability 12, no. 13: 5456. https://0-doi-org.brum.beds.ac.uk/10.3390/su12135456